
Abstract
Diatoms often outnumber other eukaryotic algae in the oceans, especially in coastal environments characterized by frequent fluctuations in light intensity. The identities and operational mechanisms of regulatory factors governing diatom acclimation to high light stress remain largely elusive. Here, we identified the AUREO1c protein from the coastal diatom Phaeodactylum tricornutum as a crucial regulator of non-photochemical quenching (NPQ), a photoprotective mechanism that dissipates excess energy as heat. AUREO1c detects light stress using a light-oxygen-voltage (LOV) ___domain and directly activates the expression of target genes, including LI818 genes that encode NPQ effector proteins, via its bZIP DNA-binding ___domain. In comparison to a kinase-mediated pathway reported in the freshwater green alga Chlamydomonas reinhardtii, the AUREO1c pathway exhibits a faster response and enables accumulation of LI818 transcript and protein levels to comparable degrees between continuous high-light and fluctuating-light treatments. We propose that the AUREO1c-LI818 pathway contributes to the resilience of diatoms under dynamic light conditions.
Similar content being viewed by others

Introduction
Photosynthetic organisms provide oxygen, food, and other resources that sustain the human society and the entire biosphere. A striking pattern exists in the distribution of eukaryotic photosynthetic groups. While terrestrial areas are predominantly covered by green plants (Viridiplantae) that harbor a plastid that likely originated from an engulfed cyanobacterium, marine environments are dominated by algae with complex plastids that may have evolved from eukaryotic endosymbionts1. Despite the presence of many aquatic species within Viridiplantae (the green algae), photosynthetic eukaryotes in oceans are mainly represented by diatoms, accounting for 20% of global primary productivity2,3. In addition to serving as a major carbon sink, diatoms represent a sustainable source of many nutraceuticals, such as the highly valued carotenoid fucoxanthin4. Therefore, comprehending the molecular mechanisms underlying the ecological success of diatoms bears significant biogeochemical and economic implications. Moreover, diatoms have unique photosynthetic properties, like their superior ability to harness blue-green light common in underwater settings5,6,7,8 (Supplementary Fig. 1). If integrated into food crops, this could help fill the green gap in their light utilization spectrum.
In their natural environment, diatoms are exposed not only to a different light spectrum compared to green plants, but also to different dynamics of light input. They therefore require distinct strategies for light acclimation. Specifically, although light provides energy for photosynthetic organisms, high levels of light can cause damage9. Acclimation to high light thus entails the activation of various mechanisms to prevent such damage.
A broadly-conserved strategy for preventing light damage involves dissipation of excess light energy as heat, a process known as the non-photochemical quenching (NPQ) of chlorophyll fluorescence9,10,11. The short-term component of NPQ in photosynthetic eukaryotes, energy-dependent quenching (qE)12, involves the concerted actions of de-epoxidized xanthophyll carotenoids and effector proteins11,13. The effector proteins of qE are proteins in the PSBS or LI818 families14,15. Notably, LI818 proteins, initially discovered in green algae, are not found in the closely-related vascular plants, but are present in the more distantly-related diatoms14,16,17,18,19,20,21,22,23. They are generally referred to as LHCSR proteins in green algae and LHCX proteins in diatoms.
Estuarine and coastal diatoms continually experience minute-scale light fluctuations due to water motion, with light intensities reaching 2000 µmol photons m−2 s−1 at mid-day24,25. They show stronger induction of NPQ relative to algae living in other habitats; furthermore, in laboratory settings mimicking fluctuating light, estuarine and coastal diatoms show greater resistance to high light than the other algae26,27,28,29,30. Previous work identified the possession of dual pathways of xanthophyll de-epoxidation in diatoms (in contrast to the single pathway in green plants) to be a cause of the strong induction of NPQ in diatoms. It was also shown that the additional pathway in diatoms acts with fast kinetics31,32,33,34. Additionally, several studies have demonstrated that certain members in the LHCX family are rapidly transcriptionally activated by the onset of high light, in some cases within 5 min22,35,36,37,38; and the importance of LHCX members in diatom NPQ has been demonstrated17,20,21,23. However, the mechanisms by which high light regulates the expression of specific LHCX isoforms remain unknown.
Recent studies in land plants, green algae, and diatoms all suggest that both chloroplast signals and photoreceptor-mediated light signaling contribute to high light acclimation11,39,40,41,42,43,44,45,46,47. For example, in the land plant Arabidopsis thaliana, the two phototropins, PHOT1 (aka. NPH1) and PHOT2 (aka. NPL1), each comprising two blue light-sensing LOV domains and a serine/threonine kinase ___domain, have overlapping functions in modulating blue light-mediated phototropism and chloroplast accumulation under low light. However, PHOT2 exhibits lower light sensitivity, i.e., it requires a higher light intensity to reach its maximum function44. Pertinent to this, PHOT2 possesses a specialized role in mediating the photoprotective chloroplast avoidance response under light stress, which is not shared by PHOT143,44. In the green alga Chlamydomonas reinhardtii, the singular phototropin (PHOT1) has been recently identified as a key player in activating LHCSR3 expression under high light conditions45. For diatoms, while tremendous progress has been made in the understanding of the chloroplast signals48,49,50, a photoreceptor that is essential for acclimation and upregulation of LHCX genes under high light has not been reported. Previous photoreceptor studies in diatoms mostly addressed gene expression or other responses to blue or red light with an intensity below 150 µmol photons m−2 s−1 51,52,53,54,55,56,57. However, exposure to higher light intensities could potentially manifest a pronounced phenotype, such as a visible bleaching phenotype45, in mutants of photoreceptors that play a key role in the regulation of photoprotection.
In this study, we took advantage of the recently developed CRISPR/Cas9 technologies in the model coastal diatom Phaeodactylum tricornutum58 to disrupt its candidate regulatory genes and investigate their roles in LHCX gene expression and acclimation to light stress. Our efforts uncovered the blue light receptor PtAUREO1c to be an important and specific regulator of genes involved in photoprotection and photosynthesis under high light. By combining biochemical, cell biological, and multi-omic approaches, we found that AUREO1c perceives blue light and instantaneously activates the transcription of two LHCX genes, in a manner quantitatively dependent on the light intensity. Finally, we demonstrated the importance of this pathway in acclimation to prolonged high light and rapidly fluctuating light.
Results
The photoreceptor AUREO1c is required for LHCX2/3 gene expression, NPQ development, and maintenance of photosynthetic performance under high light
To understand how diatoms respond to high-light stress, we initially aimed to identify the relevant LHCX family members involved in acclimation to high light in P. tricornutum. Among its four LHCX isoforms, although LHCX2 and LHCX3 are upregulated under high light39,59, P. tricornutum cells harboring a mutation in LHCX2 or silenced LHCX3 expression exhibit only modestly lower NPQ than wild-type cells20,60. To investigate whether LHCX2 and LHCX3 can functionally compensate each other, we generated and phenotyped their single and double mutants (see Supplementary Data 1 for all primers) under light stress. We found that double, but not single, mutants deficient in LHCX2 and LHCX3, were bleached under very high light (1200 µmol photons m−2 s−1 of white light; referred to as “VHL”; see Supplementary Fig. 2 and Supplementary Data 2 for culture conditions throughout the paper), suggesting that LHCX2 and LHCX3 act redundantly in maintaining cell fitness under light stress (Supplementary Figs. 3a and 4a–c). Using lower light intensities that did not cause bleaching, we observed that the double mutants show decreased NPQ and a lower fraction of variable chlorophyll fluorescence (Fv/Fm) ratio compared to wild-type cells (Supplementary Fig. 3b, c); lower Fv/Fm could arise from oxidative damage of photosystem II (PSII)61,62. These findings indicate that LHCX2 and LHCX3 play a critical function in photoprotection.
We then sought for P. tricornutum photoreceptors important for its high light acclimation. Among the multiple photoreceptors encoded by the P. tricornutum genome, three proteins (AUREO1a, AUREO1b, and AUREO1c) belong to the stramenopile-specific, blue light-sensing aureochrome family63,64. Aureochromes contain a single C-terminal LOV ___domain and an N-terminal bZIP transcriptional factor ___domain, and can mediate light-induced transcriptional regulation65,66,67,68,69. We prioritized this family because of three reasons. First, in a large-scale transcriptomics study, AUREO1b and AUREO1c appeared in the same co-expression modules with genes implicated in photosynthesis (chlorophyll biosynthesis genes, “lightsteelblue1” module) and photoprotection (LHCX3, “blue” module) respectively70. Secondly, previous studies demonstrated the importance of blue, but not red, light signals in the induction of LHCX2/3 expression39,59,71. Thirdly, photoreceptors exhibit conformational changes upon exposure to light and revert to their original conformation upon exposure to darkness and the reversion kinetics are negatively correlated with their light sensitivity72. In an in vitro study, recombinant AUREO1c proteins showed faster recovery than AUREO1a proteins after blue light excitation, suggesting that AUREO1c has lower light sensitivity and may play a physiological role in high light sensing73.
We therefore mutagenized AUREO1a, AUREO1b, and AUREO1c and selected three mutants for each gene displaying unambiguous sequencing results near the guide region for further characterizations (Fig. 1a and Supplementary Figs. 4d–f and 5; “Methods”). In these selected mutants, we observed a lack of mosaicism (evident homozygosity) near the guide region. This could be the result of identical repair outcomes following biallelic cutting or gene conversion, a process regularly observed in P. tricornutum58,74. Unique insertions or deletions (InDels) were found in each of the three mutant lines, according to our sequencing results. These InDels are anticipated to induce a frameshift during translation, leading to functional inactivation of the protein (Supplementary Fig. 4). The single-nucleotide polymorphisms (SNPs) observed in wild type could be found in some of the mutants, and their disappearance in other mutants may have resulted from mismatch repair associated with gene conversion75 (Supplementary Fig. 5). All the mutants resembled wild type in appearance and physiological parameters when grown under our standard light conditions (referred to as “growth light” or “GL”; Fig. 1b, c).

a Diagrams of the three aureochromes in P. tricornutum. b Photographs of cultures of wild-type (WT) and aureochrome mutants grown under our standard light conditions (GL, 40 µmol photons m−2 s−1 of white light) and treated for 2 days under very high light (VHL; 1200 µmol photons m−2 s−1 of white light). c Maximal quantum yields of photosystem II (PSII) for WT cells and aureochrome mutants, after growth under GL and 3 days of treatment under high light (HL; 550 µmol photons m−2 s−1 of white light), measured as the ratio of variable chlorophyll fluorescence (Fv) to the maximum fluorescence (Fm) after dark acclimation (see “Methods”). The Fv/Fm ratios are shown as false-color images, with values indicated below each image. d NPQ phenotypes of WT, the aureo1c-1 mutant, and the COMP strain treated for 6 h under 900 µmol photons m−2 s−1 of white light. Data are presented as mean values ± standard deviations (SD). e Relative transcript abundance of LHCX2 and LHCX3 in WT, aureochrome mutants, and the aureo1c-1 complemented line (COMP; expressing the wild-type AUREO1c gene in aureo1c-1 mutant cells) normalized to the average of the WT-GL samples. The measurements were performed by RT-qPCR. Data are presented as mean values ± SD. The GL and HL growth conditions were the same as those in (c). HL treatment continued for 1 h before cells were collected for analysis. The expression level of LHCX2/3 in aureochrome mutants and COMP line under high light conditions was compared with that of WT cells using a two-sided t-test. *p < 0.05; **p < 0.01; ns, not significant. For LHCX2, the p values for the comparisons were 0.0061 for aureo1a-1 vs. WT, 0.0002 for aureo1b-1 vs. WT, 0.002 for aureo1c-1 vs. WT, and 0.002 for COMP vs. WT, respectively; for LHCX3, the p values were 0.493 for aureo1a-1 vs. WT, 0.040 for aureo1b-1 vs. WT, 0.016 for aureo1c-1 vs. WT, and 0.291 for COMP vs. WT, respectively. f Phenotypic rescue of aureo1c-1 by complementation with the wild-type AUREO1c gene (the COMP strain), observed as appearance under VHL (same condition with VHL in (b)). In addition, photographs of the cultures of the two other mutant lines of AUREO1c are shown. g Phenotypic rescue of aureo1c-1 by genetic complementation based on the Fv/Fm ratio. The GL and HL conditions were the same as those in (c). Uncropped images with additional replicates (independent cultures or independent mutant lines) for (c, g) are provided in Supplementary Fig. 6a, b. For (b–g), three independent cultures were used for the measurements. The experiment was repeated three times independently with consistent results and a representative result is shown. Source data are provided as a Source Data file.
Under VHL, aureo1c-1 cells were bleached, while aureo1a-1 and aureo1b-1 mutants were not (Fig. 1b). Under lower light intensities, aureo1c mutants exhibited a lower Fv/Fm ratio and NPQ (Fig. 1c, d). We conclude that aureo1c-1, but not aureo1a-1 or aureo1b-1, resembled lhcx2 lhcx3 double mutants (Supplementary Fig. 3) in high light sensitivity and AUREO1c is important for high light acclimation.
To delve deeper into the function of AUREO1c, we examined the performance of the aureo1c-1 mutant under VHL using quantitative methods and over a time scale of hours to better simulate natural habitats. Wild type and the aureo1c-1 mutant did not bear a major difference in pigment composition in GL (Supplementary Table 1). During the first day of VHL exposure, the cell concentration of the wild type doubled, whereas the aureo1c-1 mutant exhibited no growth (Supplementary Fig. 7a). After a full day of exposure to VHL, a substantial loss in chlorophyll fluorescence was observed in most aureo1c-1 cells, as evidenced by flow cytometry (Supplementary Fig. 7b–h). Concurrent HPLC measurements indicated an 8-fold decrease in chlorophyll a content in wild type but a 60-fold decrease in the aureo1c-1 mutant, compared to the GL control cells (Supplementary Table 1). Following 2 days of VHL treatment, the pigment levels in the aureo1c-1 cells approached our detection limit, rendering accurate quantification unfeasible. We decided to assess photosynthetic parameters within the initial 6 h of VHL treatment, during which the aureo1c-1 mutant still retained chlorophyll levels comparable to the wild type (Supplementary Table 1). At this stage or earlier timepoints, we observed the mutant exhibiting a lower Fv/Fm (noticeable as early as 2 h into the treatment), and NPQ (measured at 4 h) already (Supplementary Figs. 8 and 9), demonstrating that AUREO1c’s role is likely important in the natural habitats, where VHL would not last longer than several hours.
We then investigated the mechanism how AUREO1c might impact NPQ. Our analysis of the expression of the four LHCX genes revealed that aureo1c-1 deviated markedly from the wild type, showing a lower accumulation of LHCX2 and LHCX3 transcripts under high light conditions (Fig. 1e and Supplementary Fig. 10). Because NPQ also depends on the activity of de-epoxidized xanthophyll carotenoids, we measured the xanthophyll de-epoxidation state (DES). Intriguingly, the aureo1c-1 mutant exhibited a DES higher than wild type under two different light intensities tested (6 h in 1200 µmol photons m−2 s−1, Supplementary Table 1; 2 days in 900 µmol photons m−2 s−1, Supplementary Table 2). These results suggest that AUREO1c is specifically required for induction of LHCX2 and LHCX3 under high light. Intriguingly, the DES in the aureo1c-1 mutant is elevated compared to that in wild type. A higher DES was also found in the C. reinhardtii npq4 mutant16 and this may be a common compensatory response to impaired NPQ.
To rule out potential off-target effects associated with the single guide RNA chosen for mutagenesis, we performed genetic complementation on one of the mutants disrupted in AUREO1c (“Methods”). The complemented line (COMP), expressing an AUREO1c-GFP fusion protein in the background of the aureo1c-1 mutant, exhibited rescued LHCX2/3 expression, NPQ, and Fv/Fm, and showed tolerance to very high light (Fig. 1d–g and Supplementary Figs. 6b, 7–9 and 11), confirming that the phenotypes of the aureo1c-1 mutant were caused by the disruption of AUREO1c. We observe that the COMP line was even higher than wild type in NPQ (Fig. 1d and Supplementary Fig. 9), higher in LHCX2/3 expression in certain conditions (Fig. 1e; “Dark to GL 10 min” samples in Supplementary Fig. 13), and lower in DES (Supplementary Tables 1 and 2). This apparent “over-complementation” could potentially be attributed to the use of the potent EF1α promoter, which drives the expression of AUREO1c-GFP in this line (“Methods”).
Based on the above results, we conclude that AUREO1c is the key regulator required for the NPQ response and for survival of P. tricornutum after exposure to high light. Furthermore, it exerts its function (partially) through regulating the expression of LHCX2/3 genes but not xanthophyll de-epoxidation.
The contribution of AUREO1c to the accumulation of LHCX2/3 transcripts increases with light intensity
In A. thaliana, the function of PHOT1 in modulating phototropism reaches saturation at light intensities above 10 µmol photons m−2 s−1. Conversely, the function of PHOT2 remains unsaturated at this intensity, aligning with its physiological role in regulating photoprotective responses under high light conditions42,44. To explore the effective light intensity range for AUREO1c to regulate LHCX2/3 expression, we measured the expression of these two genes in wild type and the aureo1c-1 mutant samples exposed to varying light intensities. In wild type, we observed a continuous increase in LHCX2/3 expression with escalating light intensity up to the highest intensity investigated (640 µmol photons m−2 s−1 of white light). Furthermore, the ratio of LHCX2/3 expression in wild type to that in the aureo1c-1 mutant also showed a continuous increase within this range (Supplementary Fig. 12), instead of saturating at a low light intensity. This indicates that AUREO1c function indeed has a potential to serve as a quantitative indicator for the onset of high light stress.
Light intensity fluctuates during the day, and also changes during day-night transitions. In the case of C. reinhardtii, it has been observed that during dawn, despite the low light intensity, the transition from darkness to light also activates CrLHCSR3 expression76; and this is partially dependent on CrPHOT1. Analogously, we found a similar response in PtLHCX2 and PtLHCX3, which were expressed at a higher level when dark-acclimated cells were shifted to growth light, akin to CrLHCSR3; and the aureo1c-1 mutant was partially blocked in this process (Supplementary Fig. 13). Moreover, the aureo1c-1 mutant exhibits a deficiency in the upregulation of PtLHCX2/3 during the transition of cells from darkness to high light, in addition to the previously observed deficiency during the shift from growth light to high light (Fig. 1e and Supplementary Fig. 12). These findings suggest that AUREO1c functions in modulating LHCX2/3 across various degrees of light intensity augmentation.
Subsequently, we evaluated LHCX2/3 expression under a spectrum of blue light intensities after a period of incubation in darkness (Supplementary Fig. 14). For a comprehensive understanding, we deliberately incorporated light intensities as low as 5 µmol photons m−2 s−1, the minimal threshold our laboratory apparatus can provide, to investigate the “minimum light intensity” necessary to activate AUREO1c’s role in modulating LHCX2/3 expression. Our observations were multifold. Initially, we noted that LHCX2/3 expression in the wild type augmented in parallel with blue light intensity. Transitioning from darkness to 5 µmol photons m−2 s−1 of blue light, LHCX2 and LHCX3 respectively exhibited approximately 7-fold and 4-fold increases in transcript abundance. This finding aligns with a prior study54, although their documented upregulation was more pronounced (25-fold at 3.3 µmol photons m−2 s−1 of blue light) and this may be related to the different lengths of acclimation in dark (60 h in their study versus 12 h in this study). Secondly, we observed a progressive increase in the ratio of LHCX2/3 expression in the wild type compared to the aureo1c-1 mutant. Thirdly, even under a light intensity of 5 µmol photons m−2 s−1, the wild-type expression surpassed that of the aureo1c-1 mutant by a factor of 2–3. However, the minimum intensity of blue light required to elicit an observable activation of AUREO1c function—specifically, as evidenced by a discernible difference in LHCX2/3 expression from the wild type—remains undetermined.
We observed a few complications in the above experiments. First, it’s important to note that regulators of LHCX2/3 expression extend beyond just AUREO1c. These additional factors become particularly noticeable during transitions from darkness to growth light or high light (“Dark to GL 10 min” and “Dark to HL 10 min” samples in Supplementary Fig. 13), or to blue light (Supplementary Fig. 14). During these transitions, the aureo1c-1 mutant also demonstrated a prominent induction of LHCX2/3 relative to its own control in darkness. The AUREO1c-independent factors, which could possibly encompass the electron transport chain or other photoreceptors39, remain to be conclusively identified and warrant further investigation. Secondly, for wild type, the aureo1c-1 mutant, and the COMP line, cells switched from darkness to growth light 10 min prior to the quantification (“Dark to GL 10 min” samples in Supplementary Fig. 13) had a higher level of LHCX2/3 transcript abundance than cells cultured in GL (“GL” samples). This outcome is not surprising because gene induction can be transient and relaxed after a period of time39,40.
The aforementioned results, coupled with the presence of the bZIP ___domain, support the notion of AUREO1c functioning as both a blue light sensor and a transcription factor. Nevertheless, certain caveats exist: (1) The observed upregulation of LHCX2/3 expression in conjunction with increasing light intensity via AUREO1c could potentially be attributed to the effect of amplified light energy, rather than light signaling; (2) The localization and DNA-binding capabilities of AUREO1c remain unexplored through experimental investigation. In the following sections, we address these considerations in greater detail.
AUREO1c is a nucleus-localized sensor for blue light
To gain insights into the mechanism by which AUREO1c may regulate NPQ and LHCX gene expression, we first examined its subcellular localization. To do so, we took advantage of the COMP line that expresses the AUREO1c-GFP fusion protein. We found that the GFP fluorescence signal co-localized with DNA staining by 4′,6-diamidino-2-henylindole (DAPI) in cells grown under both growth light and high light. Control wild-type cells did not exhibit a visible signal in the GFP channel (Fig. 2a). These results indicate a nuclear localization of AUREO1c, consistent with its proposed role as a transcription factor. Moreover, our results suggest that light level does not affect its localization. Instead, it may alter its conformation, as reported previously for other aureochromes77,78,79 and supported by the evidence presented below.

a Localization of AUREO1c-GFP protein under growth light (GL) and high light (HL). Wild-type (WT) cells lacking the transgene did not show detectable signal in the GFP channel, suggesting the signal for the COMP line is from the AUREO1c-GFP fusion protein. b Schematic showing conditions and treatments for transcriptome analysis. Three independent cultures were sampled each for WT and the aureo1c-1 mutant. HWL, HRL, HBL stand for high white light, high red light, and high blue light respectively. The photosynthetic active radiation (PAR), photosynthetic usable radiation (PUR), and the blue light fraction of the PAR [PAR (BL)] for each condition were calculated and shown in the table. See “Methods”, Supplementary Fig. 2 and Supplementary Data 2 for further details. c Principal component (PC) analysis of the transcriptomes of WT and the aureo1c-1 mutant under various light conditions. Note that PC1 explains the bulk (77%) of the variation. d Correlation between the effects of the aureo1c-1 mutation on transcriptomes under different conditions. The y-axes show the log2 values of the ratios between normalized expression levels in WT and mutant cells, under one of these four light regimes: GL, HRL, HBL1, and HBL2. The corresponding values under HWL condition are plotted along the x-axes. Each dot represents one gene. The squared Pearson’s correlation coefficients (R2) are indicated. All correlations had a two-sided p value < 0.0001. In each quadrant, the number of genes showing significant (FDR < 0.01; see “Methods”) differences between WT and the mutant under both conditions is shown in the corner. e Size-exclusion chromatography of the recombinant proteins of AUREO1c, after dark incubation or blue light irradiation, followed by crosslinking. The molecular weight (Mw) labels are based on data in Supplementary Fig. 15. f Absorption spectra of recombinant AUREO1c proteins after dark incubation or blue light irradiation. For (a, e, f), the experiment was repeated three times independently, yielding similar results and a representative result is shown.
To understand whether the function of AUREO1c is blue light signal-associated, we compared the transcriptomes of the aureo1c-1 mutant and wild-type cells under different light input conditions: growth light (GL), high white light (HWL), high red light (HRL), high blue light 1 (HBL1) and high blue light 2 (HBL2) (Fig. 2b, Supplementary Fig. 2, and Supplementary Data 2). HBL2 was similar to HRL and HWL in photosynthetic active radiation (PAR) but higher in photosynthetically usable radiation (PUR) by 70% and 36%, respectively. HBL1 was additionally included to control for the actual photosynthetic energy intake; it was 18% lower than HWL and similar to HRL in PUR (Fig. 2b). Each of these conditions yields a distinct transcriptomic signature (Supplementary Data 3 and 4). However, principal component analysis (PCA) revealed major deviations of aureo1c-1 mutant cells from wild-type cells under HWL, HBL1, and HBL2, but not under GL or HRL conditions (Fig. 2c). This analysis suggests that the deficiency in AUREO1c function mainly affects the cells’ transcriptomic response to an increase in blue light, rather than total input light energy.
We further confirmed the blue-light specificity of AUREO1c by comparing the effect of aureo1c-1 mutations on the transcriptome of cells grown under different light conditions (GL, HRL, HBL1, and HBL2) to the effect of aureo1c-1 mutations on cells grown under HWL. We observed a positive correlation between transcriptomic changes under the two HBL conditions and HWL. In contrast, correlations between the effects on cells grown under GL or HRL and those grown under HWL were rather weak (Fig. 2d), despite that HRL and HBL1 were similar in PUR. Specifically, the aureo1c-1 mutation caused consistent changes from wild type in the expression of more than 300 genes between HWL and HBL1/HBL2; “consistent” indicates a positive value in both the HWL and the HBL1/HBL2 axes or a negative value in both axes (Fig. 2d). By contrast, only around 100 or fewer genes showed a consistent change caused by the aureo1c-1 mutation when HWL was compared to GL or HRL conditions (Fig. 2d and Supplementary Data 5). These results further support that AUREO1c orchestrates P. tricornutum gene expression responses to high light by sensing the strong blue light component, and validate that our datasets can be used to characterize the AUREO1c-dependent response to blue light.
Since AUREO1c mediates a response to blue light, we asked how blue light might impact the protein to allow regulation of its target genes. Previous work suggested distinct, protein-specific models of aureochrome activation. For instance, it was discovered that VfAUREO1 from the xanthophyte Vaucheria frigida transitions from a monomer to a dimer after blue light exposure, while PtAUREO1a remains predominantly dimeric in both dark and illuminated conditions69,77,78,80,81. We investigated the elution profiles of recombinant AUREO1c protein in size-exclusion chromatography (SEC) after a period of incubation in the dark or under blue light. The blue light-exposed sample eluted earlier than the dark-incubated sample (Fig. 2e). The absorption spectra of dark- or blue light-incubated AUREO1c proteins proved that the chromophore flavin mononucleotide (FMN) was embedded in the recombinant proteins that we obtained. Specifically, we detected a peak at 450 nm in the dark-incubated sample, attributable to non-covalently bound FMN, and a peak at 390 nm in the blue light-exposed sample, corresponding to the FMN-cysteine adduct82 (Fig. 2f). Because earlier elution in SEC can be caused by either a larger size (associated with oligomerization) or a change in protein shape, we applied ultra high-performance liquid chromatography-native mass spectrometry (UHPLC–nMS) to measure the molecular weight of the protein samples. Both blue light-exposed and dark-incubated AUREO1c samples were determined to be predominantly dimerized (Supplementary Fig. 15). This indicates that AUREO1c can form a dimer under both conditions, a characteristic reminiscent of the behavior observed in AUREO1a78. The structural differences between dark-state and lit-state AUREO1c dimers await further investigations.
AUREO1c regulates multiple processes related to photosynthesis and photoprotection, and can bind the promoters of LHCX2 and LHCX3
Given its nuclear localization and the presence of a bZIP transcriptional regulatory ___domain, AUREO1c is likely to function as a transcriptional regulator. We therefore sought to identify its targets. The above transcriptomic analysis showed that HWL, HBL1 and HBL2 caused both AUREO1c-dependent and AUREO1c-independent changes from GL (Fig. 2c). To identify genes specifically regulated by AUREO1c, we selected genes that showed differential regulation under HWL relative to GL in wild-type cells and then asked whether those changes were disrupted in the aureo1c-1 mutant (“Methods”). Gene ontology (GO) analysis showed that the AUREO1c-dependent set is enriched for genes involved in photosynthesis and chloroplast activities, whereas the AUREO1c-independent set is enriched for genes involved in other processes, such as nicotinamide adenine dinucleotide (NAD) binding (Fig. 3a and Supplementary Data 6).

a Enriched gene ontology (GO) terms for genes regulated in HWL compared to GL in wild type (WT), separated into AUREO1c-dependent and AUREO1c-independent gene groups (see “Methods” for definitions). These GO terms had an adjusted p value (p. adjust) <0.05. The p values were obtained from the one-sided Fisher’s exact test used in the clusterProfiler package, and Benjamini–Hochberg method was used to adjust for multiple comparisons (see “Methods”). The length of each bar is calculated based on its adjusted p value. b A scatterplot of all the genes in P. tricornutum genome showing their responses to HWL in WT cells and the effect of the aureo1c-1 mutation on their expression. LHC family genes and nuclear genes encoding subunits of the photosynthetic electron transport chain (ETC) are highlighted. The data plotted were averages from three independent cultures. c Genes encoding ETC components color-coded to show relative expression levels. The ratio of the transcript abundance of ETC genes between aureo1c-1 and WT under HWL is shown as a heatmap; red and blue indicate higher and lower expression in aureo1c-1 compared to WT respectively. Many of the ETC subunits are encoded by the chloroplast genome and could not be detected in regular RNA-Seq experiments; these are shown as the gray background for each complex. Association between the light-harvesting complex (LHC) family members with photosystem II or photosystem I has not been entirely elucidated. They are shown as green rods here, with their transcript abundance details in (d). Fld flavodoxin, FNR ferredoxin-NADP+ oxidoreductase, PS photosystem, Cyt b6f cytochrome b6f complex. Two FLD and four FNR gene isoforms were detected and plotted. The data plotted were averages from three independent cultures. d A heatmap showing the expression levels of the chlorophyll-binding protein-encoding genes in the LHC family. The genes were hierarchically clustered into five major categories (I–V). The expression level for each of the three independent cultures under each condition was plotted separately as a column. e DNA affinity purification sequencing (DAP-Seq) results showing enrichment of promoter regions of LHCX2 and LHCX3 in sequences bound to the AUREO1c protein. Note that different gene model annotations exist for LHCX2, and the Phatr2_54065 model shown in this panel is supported by our RNA-Seq and PCR validation results presented in Supplementary Fig. 16. This experiment was repeated twice independently with consistent results and a representative result is shown. f Electrophoretic mobility shift assays (EMSAs) showing the binding of recombinant AUREO1c to DNA probe harboring sequences from the promoters of LHCX2 and LHCX3 respectively. The competing unlabeled probes were applied in excess, as indicated (50–200x). An upper shift of the labeled probes suggests their binding to the AUREO1c protein. The experiment was repeated three times independently, yielding similar results and a representative result is shown. Source data are provided as a Source Data file.
As an alternative approach to characterizing AUREO1c-dependent genes, we specifically examined the expression of candidate genes that may play a role in sunlight energy harvesting and conversion under growth light or in NPQ under high light6. These include genes encoding components of the photosynthetic electron transport chain (ETC) and genes in the light-harvesting complex (LHC; including LHCX) family (Fig. 3b). Most of these genes were downregulated in wild-type cells upon exposure to HWL, but retained a higher expression level in the aureo1c-1 mutant cells under the same conditions. A smaller number of genes were upregulated in wild-type cells in response to HWL compared to GL, but not in aureo1c-1 mutant cells (Fig. 3b). This indicates that AUREO1c is important for both down- and up-regulation of high light-responsive genes.
We next investigated ETC and LHC genes in detail. Among ETC genes, we observed a specific effect on PSII. Under HWL, aureo1c-1 mutant cells expressed higher levels of PSII reaction center subunits compared to wild-type cells (Fig. 3c), suggesting a role for AUREO1c in modulating the abundance of and potentially the ratios between different ETC complexes. Among LHC genes, we identified five different response groups, two of which depended on AUREO1c. Most of the LHCF genes (category I) were downregulated in wild-type cells under HWL, HBL1, and HBL2 compared to GL. LHCX2, LHCX3, LHCR6, LHCR8, and LHCR10 (category II) were up-regulated under HWL, HBL1, and HBL2 relative to GL, consistent with previous reports22,38,39,59 (Fig. 3d). Changes in the expression of genes in both of these categories were abolished or weakened in the aureo1c-1 mutant cells, suggesting that their regulation by blue light depended on AUREO1c. Category III genes were induced by HBL1/HBL2 in wild-type cells, but in an AUREO1c-independent manner. Finally, LHCF15 (category IV) and LHCZ1 (category V) did not fall into these three clusters, consistent with previous expression clustering studies carried out in wild type59,83.
The lower read counts for LHCX2 and LHCX3 in aureo1c-1 mutant compared to wild-type cells is consistent with our RT-qPCR results presented above (Fig. 1e), providing a validation of the transcriptome datasets and confirming that LHCX2 and LHCX3 are regulated by AUREO1c at the transcriptional level.
LHCX2 and LHCX3 are among the most prominent downstream effectors of AUREO1c, based on the significant change in gene expression levels between wild-type and aureo1c-1 mutant cells under HWL (Fig. 3b). They also show a significant difference in gene expression between HWL and GL in wild-type cells. These observations prompted us to ask whether LHCX2 and LHCX3 might be direct targets of AUREO1c. Because chromatin immunoprecipitation sequencing (ChIP-Seq) methods have not been established for transcription factors in P. tricornutum, we investigated whether AUREO1c could bind the promoter of LHCX2 and LHCX3 by performing DNA affinity purification sequencing (DAP-Seq)84. Enriched signals were observed at the promoters of these two genes, but not in the negative control sample performed using unfused GST proteins instead of GST-AUREO1c (Fig. 3e and Supplementary Fig. 16). We further confirmed the interactions between LHCX2/3 promoters and AUREO1c by performing an electrophoretic mobility shift assay (EMSA). The probe containing the LHCX2 or LHCX3 promoter sequence exhibited reduced mobility in the presence of AUREO1c (Fig. 3f). Adding a large excess of unlabeled probes reverted the effect on mobility of the labeled probes; in contrast, when the competition was performed using the promoter sequence of alpha tubulin, a gene not affected by the aureo1c-1 mutation (Supplementary Data 4), most of the labeled probes were still in the upper band. These results demonstrate that AUREO1c can bind to the promoter of LHCX2 and LHCX3 in a sequence-specific manner and may regulate their transcription directly.
Rapid induction of LHCX2/3 gene expression in P. tricornutum enables a strong response to fluctuating light
In the green alga Chlamydomonas reinhardtii, the high light-sensing phototropin protein (PHOT kinase) also contains a LOV ___domain and it transmits strong blue light signals to upregulation of LHCSR genes, by inhibiting a ubiquitin ligase complex that degrades the transcription factors required for LHCSR expression85,86,87. The direct regulation of LHCX expression by AUREO1c suggested that diatoms may be able to mount a faster response to high light. To test this idea, we compared the kinetics of LI818 induction in diatoms and green algae using RT-qPCR over a time course during exposure to HBL. Interestingly, PtLHCX2 and PtLHCX3 transcript abundance was already statistically significantly elevated, relative to baseline, within 2 min of HBL exposure (Fig. 4a). By contrast, CrLHCSR3.1 and CrLHCSR3.2 required 6 min to reach significant induction; in this experiment, C. reinhardtii cells were grown photoautotrophically in the high-salt (HS) medium as P. tricornutum cells were (Fig. 4a). Besides the HS medium, C. reinhardtii can grow mixotrophically in the tris-acetate-phosphate (TAP) medium and acetate was found to repress CrLHCSR3 expression40. However, the induction kinetics of CrLHCSR3 transcript accumulation were consistent in TAP medium (Supplementary Fig. 17). The similar fold changes noted after 10 min of high light exposure between HS- and TAP-grown cells could be attributed to an elevated baseline expression in GL conditions in the HS medium (Supplementary Fig. 18), which is consistent with previous studies40.

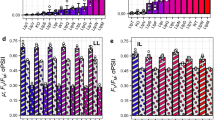
a Transcript abundance of LI818 genes in wild-type C. reinhardtii (shown in green) and P. tricornutum (shown in orange brown) based on RT-qPCR at different time points after the switch from growth light (GL; 40 µmol photons m−2 s−1 of white light) to high blue light (260 µmol photons m−2 s−1). Three independent cultures were used for the quantification. Data are presented as mean values ± SD. The data were analyzed by using a one-sided unpaired t-test (*p < 0.05; **p < 0.01; and the p value corresponding to ** or * for LHCSR3.1, LHCSR3.2, LHCX2 and LHCX3 are 0.0024, 0.0048, 0.0183 and 0.00009 respectively). Correction for multiple comparisons was not performed. b Diagram of light regimes for LI818 transcript abundance and LI818 protein abundance analysis. Cells were initially grown under GL and then switched to continuous high light (CL; 550 µmol photons m−2 s−1 of white light) or fluctuating light (FL; 550 µmol photons m−2 s−1 of white light alternating with 40 µmol photons m−2 s−1 of white light). c Transcript abundances of LI818 genes in wild-type C. reinhardtii and P. tricornutum, after CL or FL treatments, relative to those under GL, measured by RT-qPCR. Samples from each condition receiving the same total length of high light treatment were compared. Three independent cultures were used for the quantification. Data are presented as mean values ± SD. d LI818 protein abundance in C. reinhardtii wild type (WT), P. tricornutum WT and the aureo1c-1 mutant, measured by proteomics. CrLHCSR3.1 and CrLHCSR3.2 are identical in amino acid sequence and are shown collectively as “CrLHCSR3” for C. reinhardtii WT, the data were normalized to the protein abundance of LHCSR3 under GL (shown as a dashed line). For P. tricornutum WT and the aureo1c-1 mutant, the data were normalized to the protein abundance of LHCX2 or LHCX3 in WT under GL (shown as a dashed line). Two independent cultures were used for the quantification and the means are presented. This experiment was repeated twice independently with similar results and a representative result is shown. e The NPQ of C. reinhardtii and P. tricornutum after 2 h of CL and FL treatments. Three independent cultures were used for the quantification. Data are presented as mean values ± SD. f Maximal quantum yields of PSII (Fv/Fm) of wild type (WT), and two lhcx2 lhcx3 mutants grown under fluctuating light (as the FL scheme in b) for 5 days. The Fv/Fm ratios are shown as false-color images with values shown below. g Maximal quantum yields of PSII (Fv/Fm) of WT, the aureo1c-1 mutant, and the COMP line, grown under fluctuating light (as FL in b) for 5 days. Uncropped images with additional replicates (independent cultures) for (f, g) are provided in Supplementary Fig. 6c, d. For (a, c, e–g), the experiment was repeated three times independently with similar results and a representative result is shown. Source data are provided as a Source Data file.
We hypothesized that a rapid induction of LHCX2 and LHCX3 expression in P. tricornutum may be critical in the context of the rapid, minute-scale, light fluctuations in estuarine or coastal habitats24,25. By contrast, the C. reinhardtii pathway may not enter a rapid accumulation phase for LHCSR3 transcripts in each high light period. To test this hypothesis, we compared two light stress regimes: continuous high light treatment (CL) versus fluctuating light treatment (FL) with repeated cycles of 5-min high light followed by 5-min growth light. We asked whether the P. tricornutum kinetic pattern would allow significant accumulation of LHCX2/3 transcripts and proteins (and hence protection from light stress) under the FL regime. We performed RT-qPCR to compare LI818 expression between comparable CL and FL treatments in both C. reinhardtii and P. tricornutum. Transcript abundances of PtLHCX2 and PtLHCX3 were similar after either FL or CL treatment, bearing a 50–75x upregulation from 0 to 30 min of treatment (Fig. 4b, c). The drop in PtLHCX3 expression after 1 h of high light treatment is consistent with previous observations39. CrLHCSR3.1 or CrLHCSR3.2 was expressed to a much lower level in the FL regime compared to the CL regime in both the HS medium (Fig. 4c) and the TAP medium (Supplementary Fig. 19), at 30 and 60 min of treatment. It should be noted that in cells cultivated in the HS medium, the LHCSR3 gene expression ceased to escalate after 60 min of CL treatment (Fig. 4c and Supplementary Fig. 20) whereas this cessation has not occurred at 60 min in the TAP medium (Supplementary Fig. 19). We further caution that the difference between FL and CL in C. reinhardtii is in part due to the large fold increase in the CL samples.
To confirm the functional relevance of the above results, we also examined LI818 protein abundance. We performed quantitative proteomics to measure the abundance of LI818 proteins in C. reinhardtii and P. tricornutum (Supplementary Data 7–9). We sampled wild-type cells grown under normal light conditions right before the transition to CL or FL (0 min), wild-type cells after 120 min of CL treatment, and FL cells that had undergone 120 min of high-light periods in total. LHCSR3 protein accumulation in C. reinhardtii was lower under FL compared to CL in HS medium (Fig. 4d), and the difference is more pronounced in TAP medium (Supplementary Fig. 21). In contrast, LHCX2 protein abundance increased to a similar degree under either CL or FL in P. tricornutum (Fig. 4d). Consistent with the above transcript abundance data, LHCX3 induction at 120 min of either CL or FL was limited. These results support the robustness of the P. tricornutum kinetic pattern when environmental light intensity fluctuates in minute-scale cycles. Conversely, the kinetic pattern in C. reinhardtii shows a less efficient induction of NPQ effectors under FL compared to CL. Corroborating these findings, we observed a lower NPQ in C. reinhardtii following FL treatment compared to CL treatment (Fig. 4e). Intriguingly, despite similar levels of LHCX2 and LHCX3 proteins, P. tricornutum displayed a higher efficiency in inducing NPQ under FL conditions compared to CL. This finding does not contradict our model, but does underscore the potential influence of various confounding factors that may affect NPQ. Further investigations are warranted to elucidate these effects.
Finally, for P. tricornutum, we additionally asked whether the ability to accumulate LHCX proteins under CL or FL depended on AUREO1c. To address this question, we compared protein accumulation between wild-type and aureo1c-1 mutant cells at the same timepoint (120 min) under either CL or FL. The aureo1c-1 mutant was unable to induce LHCX2 protein accumulation by either CL or FL treatment (Fig. 4d). For LHCX3, aureo1c mutations did not significantly impact protein abundance. However, the limited effect may be due to the fact that LHCX3 protein accumulation was already limited at 120 min even in wild-type cells. These results are consistent with the idea that AUREO1c is required for an optimal response to fluctuating light in P. tricornutum. Further supporting this idea, we found that the aureo1c-1 mutant and lhcx2 lhcx3 double mutant in P. tricornutum showed sensitivity to prolonged growth in FL (Fig. 4f, g).
We conclude that the AUREO1c-dependent regulatory strategy used in P. tricornutum to respond to light stress increases the robustness of this organism under conditions of rapidly fluctuating light.
Discussion
In our research, we initially discovered the critical role of AUREO1c in acclimating P. tricornutum cells to high light conditions and in initiating the expression of the NPQ effector genes LHCX2 and LHCX3. We further analyzed the transcript abundance of LHCX2 and LHCX3 under various light treatments in both the wild type and an AUREO1c mutant (aureo1c-1), demonstrating that AUREO1c functions across a broad range of light intensities, notably exerting a stronger influence under more intense light conditions. We also conducted RNA-Seq under different colors and intensities of light and found that AUREO1c’s function is contingent on blue light. Collectively, these data support a model where AUREO1c detects blue light signals and triggers high-level NPQ effector gene expression under high light conditions. Indeed, previous work, in both laboratory settings and environmental surveys, has shown that diatom AUREO1c proteins are highly expressed during the day68,78, consistent with a role in photoprotection.
Our finding that AUREO1c acts by regulating the transcription of LI818 genes (LHCX2 and LHCX3) reveals unexpected convergence between diatoms and green algae, groups that are only distantly related to each other, having separated about two billion years ago88. Although both express blue-light sensors—PHOT kinases in green algae and aureochromes in diatoms—that contain LOV domains, the proteins differ in topology and in the way they activate downstream effects. PHOT proteins contain a kinase ___domain is C-terminal to the LOV ___domain, whereas aureochromes possess a bZIP ___domain is positioned N-terminal to the LOV ___domain. It is therefore remarkable that both species use of LOV family photoreceptors to control the expression of orthologous NPQ effector genes. The linking of the LOV ___domain with a bZIP ___domain in the diatom AUREO1c protein has streamlined the NPQ regulatory pathway, allowing AUREO1c to directly regulate LHCX2/3 (Fig. 3e). This rewiring obviates the need for an additional regulatory step, and likely enables the rapid induction of LHCX2 and LHCX3 proteins in response to high light. This observation is consistent with a recent modeling study predicting that for the same input and output, molecular switch architecture based on direct activation (rather than derepression) allows faster induction of the output89. This dynamic regulation feature of diatoms likely contributes to their dominant occupancy of coastal habitats (Fig. 5). On the other hand, the green algal PHOT-LHCSR3 pathway may allow transcription factors to increase in abundance over time, leading to a faster-than-linear accumulation of LHCSR3 transcripts; this rapid accumulation was observable within a span of 30 min for cells cultivated in the HS medium, and within 120 min for those grown in the TAP medium (Supplementary Figs. 19 and 20). This pattern is potentially advantageous when high light duration is longer than several minutes. Our study provides a model system for comparative characterizations of the different regulatory strategies.

Note that factors beyond the photoreceptor-mediated pathways may also contribute to the kinetic features of LI818 gene induction, as cautioned in the “Discussion”.
We would like to underscore several precautionary points that could guide future research directions. Firstly, the breadth of light conditions we could test was confined by the limitations of our laboratory equipment. For a more comprehensive validation of our model, future research should consider investigating a wider array of light conditions. In particular, utilizing conditions like sinusoidal fluctuations may more closely mimic natural light environments35. Second, as revealed in this study (Fig. 4c and Supplementary Fig. 20) and previous studies on CrLHCSR3.1, CrLHCSR3.2 (in the HS medium) and PtLHCX3, relaxation of gene induction occurs within 2 h39,40,85. To capture more detailed kinetics in future research, consideration could be given to incorporating more data collection points.
Another caveat in our study is that the AUREO1c-LHCX pathway operates in the context of pathways mediated by other photoreceptor proteins, the chloroplast electron transport chain (ETC), reactive oxygen species, and/or other regulatory proteins50,51,54,55, that together constitute the full light-stress response in P. tricornutum. For instance, the activation of LHCX2/3 expression during dark-to-light transitions also occurs in the aureo1c-1 mutant (Supplementary Figs. 13 and 14), indicating the presence of AUREO1c-independent factors to activate LHCX2/3 expression. In addition, a previous study showed the presence of negative regulators of LHCX2/3, namely CPF1, a cryptochrome-type photoreceptor54. Moreover, a recent investigation in C. reinhardtii suggested that a low intracellular CO2 level plays a crucial role in inducing CrLHCSR3 expression40; although the conservation of this pathway in diatoms remains to be investigated. We acknowledge that some of these additional factors could also contribute to the kinetic features of diatom acclimation to a dynamic light climate or even interact with the AUREO1c pathway. To disentangle the regulatory and kinetic roles of each factor, future kinetic studies could be conducted in the context of genetic or pharmacological perturbations, or under varying light conditions, including transitioning from a dark environment.
From a broader perspective, the AUREO1c light switch has significant potential in synthetic circuit design. For land plants, accelerated induction and relaxation of photoprotection can confer enhanced protection and minimize energy losses90,91. The demand for rapid high light-based switches may also increase in the context of a changing climate92. In addition, photoreceptors have been extensively employed in neuroscience and developmental biology studies, and in the design of controllable cells93. The commonly used algal channelrhodopsin ChR2 directly mediates ion transport, bypassing the involvement of G proteins, enzymes, and ion channels necessary for the action of typical animal rhodopsins, making the phototransduction instantaneous94. Analogously, the AUREO1c-LHCX promoter module circumvents a need for additional E3 ubiquitin ligase proteins and separate transcription factors. The rapid (2–3 min) response time of AUREO1c may therefore expand the light-dependent transcriptional regulatory toolbox and help overcome kinetic challenges in this field95,96. More broadly, our study demonstrates the promise of studying organisms occupying diverse niches; environmental fluctuations occur at a wide range of timescales in nature, providing a large, mostly unexplored gene reservoir for synthetic biology uses97,98,99.
Methods
Algal strains and growth conditions
The wild-type Phaeodactylum tricornutum (P. tricornutum) strain CCMP2561 was acquired from The National Center for Marine Algae and Microbiota (https://ncma.bigelow.org/). P. tricornutum cells were grown in f/2 medium at 21 °C. Solid f/2 medium was supplemented with 1% agar. Liquid cultures are shaken at a speed of 120 rpm and grown under white light (40 µmol photons m−2 s−1), which is defined as the growth light (GL) condition. Other light conditions used are described in the corresponding figure legends. For RT-qPCR analysis of LHCX gene expression on cells shifted from darkness to light, the cells were incubated in darkness for a period of 12 h before the transition. For white light, intensities in the range of 400–550 µmol photons m−2 s−1 were referred to as “high light (HL)” or “high white light (HWL)” in the “Results”; and intensities of 1200 µmol photons m−2 s−1 or above were referred to as “very high light (VHL).” The light conditions used for transcriptome analysis are additionally provided in the corresponding “Methods” section below. All the light sources were light-emitting diodes. The wavelengths for blue and red light were 450 ± 5 nm and 660 ± 5 nm respectively. The light spectrum of each culture chamber is shown in Supplementary Fig. 2. The conversion from radiometric units [W/m2/nm] to PAR (μmol photons m−2 s−1) follows the formula: PAR = I × λ × 0.836 × 10−2, where I is the light intensity in W/m2/nm and λ is the wavelength in nm. PUR is calculated from PAR using the formula: \({\int }_{400}^{700}{{{{{\rm{PAR}}}}}}\left({{\uplambda }}\right){{A}}({{\uplambda }}){{d}}\left({{\uplambda }}\right)\), where λ denotes the wavelength and A(λ) is the normalized absorption spectrum. Conversion of the photosynthetic active radiation (PAR) to the photosynthetic usable radiation (PUR) was based on the light absorption spectrum of P. tricornutum and C. reinhardtii cells and light spectrum of each culture chamber. The conversion was according to a formula in published work100,101. The light intensity and quality details for each experiment are shown in Supplementary Data 2.
For the green alga Chlamydomonas reinhardtii, we employed the widely used strain CC-5325 (aka. CC-4533)102. Cells were grown in the Tris-acetate-phosphate (TAP) medium103 under 22 °C. Both TAP and the acetate-free high-salt (HS) medium104 have been used for transcriptional studies of LHCSR genes40 and both were employed in our studies. When HS medium was used, cells grown in TAP were washed twice with HS medium, resuspended in HS medium and incubated overnight before our light treatments.
CRISPR/Cas9-mediated mutagenesis
CRISPR/Cas9 mutagenesis, including guide design105, construction of vectors expressing Cas9 and guide RNAs106, and conjugation-based transformation107, was carried out using the episome-based procedure described in our previous research, which generated the zep1-b mutant108. Primers that were annealed for generation of the guide RNA template are provided in Supplementary Data 1. The vectors contained a Blasticidin S Deaminase gene expression cassette that allowed for selection of successfully transformed clones on solid f/2 medium containing 5 μg/ml blasticidin S (60218ES50, Yeasen Biotechnology). The primary transformants underwent PCR amplification and sequencing of the guide region, yielding either completely wild-type or mixed sequencing results; note that we designed primers (Supplementary Data 1) from regions without polymorphisms between the two chromosomes to avoid missing the amplification of one allele. Colonies with mixed sequencing outcomes were then cultured in liquid medium and subsequently spread onto selective solid medium for another round of colony formation. The sub-clones derived from these colonies could present with wild-type sequence, mixed sequencing results, or unequivocal sequencing results. Those sub-clones still yielding mixed results could be either heterozygous or mosaic, containing a mix of mutant and wild-type cells; and the two possibilities cannot be easily distinguished in our procedure. Therefore, sub-clones with unequivocal sequencing results were selected for further physiological analyses. Note that although P. tricornutum is a diploid, it exhibits a high frequency of gene conversion58,74. This characteristic has made it easy to obtain biallelic (apparently homozygous) mutants for each targeted gene.
Complementation of the aureo1c-1 mutant (generation of the COMP line)
The coding sequence of AUREO1c was PCR amplified as two fragments from total cDNA using the Phanta Polymerase (Vazyme) and two pairs of primers (the HZ-797/HZ-798 pair and the HZ-799 and HZ-800 pair; Supplementary Data 1). The two fragments were integrated into the homemade pHZ-174 expression vector by homologous recombination (ClonExpress II One Step Cloning Kit, Vazyme). The primers HZ-798 and HZ-799 were designed to introduce synonymous mutations at the sgRNA site to prevent the transgene from being edited by Cas9 expressed from the vector used in initial mutagenesis of AUREO1c. The pHZ-174 destination vector contained the endogenous P. tricornutum EF1α promoter to drive the expression of the AUREO1c gene and a GFP open reading frame to be fused to the C-terminus of AUREO1c. In addition, it contained the Ble marker gene that confers resistance to zeocin109. The final construct was transformed into aureo1c-1 cells by electroporation110. After 30 days of selection on solid f/2 medium containing 75 μg/ml zeocin (R25001, Thermo), several clones were obtained, with one of them positive in the analysis of GFP fluorescence by flow cytometry (Supplementary Fig. 11). This line was designated the COMP line and stably maintained for further analyses.
Immunoblotting to verify AUREO1c-GFP expression in the COMP line
A total of 3 × 107 diatom cells were collected by centrifugation and subsequently disrupted by sonication using Bioruptor® Plus (Diagenode). Total proteins were extracted by RIPA buffer (P0013B, Beyotime) with concentrations determined using a bicinchoninic acid assay kit (C503021-0500, Sangon Biotech) according to the manufacturer’s instructions. After being denatured at 95 °C for 5 min and separated on a premade denaturing polyacrylamide gel, proteins were transferred onto a polyvinylidene difluoride membrane for blocking and incubations with antibodies. The primary and secondary antibodies were from SICGEN (AB2166; 1:2000 dilution) and Beyotime (A0181; donkey anti-goat, horseradish peroxidase-conjugated, 1:5000 dilution) respectively. Detection was performed using the Immobilon Western Chemiluminescent HRP Substrate kit (WBKLS0500, Millipore) according to the manual. Wild-type P. tricornutum cells were used as a negative control, and blotting using an antibody (AF5012, Beyotime, 1:2000 dilution) against α tubulin was used as the loading control.
Whole-cell proteomics
Proteomic analyses were performed at the Biomedical Research Core Facilities of Westlake University. P. tricornutum or C. reinhardtii cells were harvested by centrifugation and lysed by sonication in lysis buffer (RIPA P0013B, Beyotime) containing phosphatase and protease inhibitors (P1045 cocktail, Beyotime). For each strain and condition analyzed, two independent cultures were processed. The same amount of protein extract from each sample replicate was digested using trypsin, targeting cleavage sites at the lysine and arginine residues, and derivatized to harbor tandem mass tags111 (TMTs) using the TMT10plexTM (for P. tricornutum; 90110, Thermo) and TMT6plexTM kits (for C. reinhardtii; 90061, Thermo) respectively, following the manufacturer’s instructions. The TMT-labeled peptides were separated by a 135 min gradient elution at a flow rate of 0.300 µl/min using a Thermo UltiMate 3000-HPLC system directly interfaced with an Orbitrap Fusion Lumos Tribrid Mass Spectrometer (Thermo) equipped with FAIMS. The analytical column was a home-made fused silica capillary column (75 µm ID, 250 mm length; Upchurch, Oak Harbor, WA) packed with C-18 resin (300 Å, 2 µm, Varian, Lexington). Mobile phase A consisted of 0.1% formic acid in water, and mobile phase B consisted of 80% acetonitrile and 0.1% formic acid. The mass spectrometer was operated in data-dependent acquisition mode using Xcalibur 4.4 software, and the −40 V −55 V, and −70 V CV values of FAIMS were set as the three independent acquisition events. Under one event, a single full-scan mass spectrum in the Orbitrap (375–1800 m/z, 120,000 resolution) was followed by several data-dependent MS2 and MS3 scans. The ten most intense ions were first isolated for ion trap CID-MS2 at a precursor ion isolation width of 2 m/z, using a maximum ion accumulation time of 35 ms. Directly after each MS2 experiment, the most intense fragment ion in an m/z range between 375–1800 m/z was selected for HCD-MS3. The MS3 AGC was 1 × 105 and the maximum ion time was 200 ms. Normalized collision energy was set to 35% and 65% at an activation time of 10 ms and 35 ms for MS2 and MS3 scans, respectively. Each mass spectrum was analyzed using the Thermo Xcalibur Qual Browser (4.4) and Proteome Discoverer (2.5.0.400) for the database searching against ASM15095v2_bd (Annotation Source: Ensemble) for P. tricornutum and Chlamydomonas reinhardtii v5.6 (Annotation Source: JGI Genome Portal) for C. reinhardtii, and TMT quantification (Supplementary Data 7–9). The Sequest search parameters included a 10 ppm precursor mass tolerance, 0.02 Da fragment ion tolerance, and up to 2 internal cleavage sites. Fixed modifications included cysteine alkylation and TMT10 modification, and the methionine oxidation was a variable modification. The minimum length of peptides was six and the peptides were filtered at a 1% false discovery rate (FDR). An ANOVA test was employed for comparing individual proteins across different conditions. The protein inventory (Supplementary Data 7–9) encompasses a minimum of one unique peptide for each protein, with the precise tally of unique peptides detailed within the respective datasets. Specifically, proteins of interest such as PtLHCX2, PtLHCX3, and CrLHCSR3 were identified with a minimum of two unique peptides each.
Chlorophyll fluorescence techniques
For the analysis of the maximal quantum efficiency of photosystem II, approximately 5 × 106 cells were collected by centrifugation and resuspended in 7 μl of f/2 medium and subsequently spotted onto a 96-well plate with solid f/2 medium. For Supplementary Fig. 8, approximately 5 × 106 cells were collected and suspended in 200 μl f/2 medium. The cells were dark-adapted for 40 min prior to the measurements of minimal (Fo) and maximal chlorophyll fluorescence (Fm) by using the Imaging-PAM M-Series platform (Walz, blue light version). The maximal quantum efficiency of photosystem II was calculated as Fv/Fm = (Fm − Fo)/Fm62 and exported from the equipped software as a false-color image.
For measurements of NPQ in P. tricornutum, a total of 4.5 × 107 cells were harvested and resuspended in 2 ml f/2 medium, and dark-adapted for 40 min with shaking before NPQ detection. For C. reinhardtii, around 5 × 106 cells were used and dark-adapted for 15 min. NPQ was measured on the cell suspensions using Dual-PAM-100 (Walz, red light version) as previously described and calculated using the formula NPQ = (Fm − Fm’)/Fm'49,112. Fm: maximal fluorescence in the dark-adapted state; Fm’: maximal fluorescence at each illuminated timepoint.
For detection of cellular chlorophyll fluorescence after VHL treatment, cells were subjected to flow cytometry (CytoFLEX, Beckman Coulter). The excitation/emission wavelengths are 488 nm/690 nm. The FlowJo software (BD Biosciences) was used for data visualization.
Pigment analyses by high-performance liquid chromatography (HPLC)
For Supplementary Table 1, cells of wild type, the aureo1c-1 mutant, and the COMP line were initially grown under 40 µmol photons m−2 s−1 of white light and transferred to 1200 µmol photons m−2 s−1 of white light for 6 h and 1 day. For Supplementary Table 2, cells of wild type, the aureo1c-1 mutant, and the COMP line were initially grown under 40 µmol photons m−2 s−1 of white light and treated by switching to 900 µmol photons m−2 s−1 of white light for 10 min or 2 days. Cells were then collected by centrifugation, with pigments extracted in 90% aqueous acetone (v/v) containing 10 mM ammonium acetate, facilitated by sonication. The pigment extracts were analyzed using a UPLC-MS system (Waters/Acquity) equipped with photodiode array (PDA) & QDa detectors. The ACQUITY UPLC HSS T3 C18 column (3 μm, 100 Å, 2.1 × 150 mm, Waters) was used for resolving the pigments. Identities of pigments were assigned based on absorption properties measured by the PDA. Compositions of the mobile phase, flow rate, and the temperature of chromatography were as previously reported113. Xanthophyll de-epoxidation state (DES) was calculated as the ratio of diatoxanthin to the sum of diadinoxanthin and diatoxanthin.
Reverse transcription and quantitative real-time PCR (RT-qPCR)
Cells were collected by centrifugation and subjected to RNA extraction. For the time course experiment (Fig. 4a and Supplementary Fig. 17), at each timepoint, cells were quickly mixed with equal volumes of stop solution (60% ethanol, 2% phenol, 10 mM EDTA; pre-cooled at −20 °C) to quench their transcription114 before being centrifuged. For other experiments including light treatment shorter than 1 h (see details in Supplementary Data 2), the cells were also mixed with a stop solution in a 1:1 volume ratio. Total RNA was extracted using the RNA Easy Fast Plant Tissue Kit (DP452, Tiangen; for P. tricornutum) and the RNeasy Plus Mini Kit (74136, Qiagen; for C. reinhardtii) respectively, following the manufacturers’ instructions. Reverse transcription was performed using the HiScript III RT SuperMix for qPCR (+gDNA wiper) kit (R323, Vazyme). Each qPCR reaction consisted of 10 μl of ChamQ Universal SYBR qPCR Master Mix (Q711, Vazyme), 8.2 μl of water, 1 μl of cDNA template, and 0.4 μl each of forward and reverse primers (concentration before mixing: 10 μM). RT-qPCR was performed on the qTOWER3G system (Analytik Jena). The RPS and RACK1 genes were used as reference genes for P. tricornutum or C. reinhardtii respectively. The delta-delta Ct method115 was used for relative quantification of transcript abundance of target genes across samples. Primers employed for PtLHCX2, PtLHCX3, CrLHCSR3.1, CrLHCSR3.2, and the reference genes are provided in Supplementary Data 1.
Transcriptome sequencing
Cells of wild type and the aureo1c-1 mutant were grown in standard growth light (GL) conditions and then transferred to the following conditions: high white light (HWL; 430 µmol photons m−2 s−1), high red light (HRL; 440 µmol photons m−2 s−1), high blue light 1 (HBL1; 260 µmol photons m−2 s−1), and high blue light 2 (HBL2; 430 µmol photons m−2 s−1) and cultured for 1 h with shaking. The calculation of PUR was performed as described above in the “Algal strains and growth conditions” section. For calculation of the blue light fraction of the PAR, photons with a wavelength within 400–500 nm were included. The results are shown in Fig. 2b and Supplementary Data 2.
For transcriptome sequencing, a total of 1 × 108 cells were harvested by centrifugation and flash frozen in liquid nitrogen. RNA was extracted with the RNeasy Plus Mini Kit (74136, Qiagen). Library preparation and sequencing on the Nova-PE150 platform (Illumina) was performed by Novogene Co as a service.
Transcriptome analyses
The raw RNA sequencing (RNA-Seq) reads in the FASTQ format were filtered using fastp116 (parameters: q, 20; u, 30; n, 5; l, 130) and then mapped to genomic loci117 using HISAT2118. Sequence alignment files in the SAM format were sorted using the SAMtools119, with read counts exported120. For LHCX2, reads aligned to Chromosome 1: 2,471,163–2,472,285 (Phatr2_54065) instead of the entire Phatr3_EG02404 gene model (Chromosome 1: 2,469,549–2,472,285) were included; the Phatr2_54065 model is supported by our RNA-Seq reads and PCRs (Supplementary Fig. 16). Read count normalization (Supplementary Data 4) and principal component analysis (PCA) were performed using the DESeq2 environment121.
To compare the transcript abundances between wild type and the aureo1c-1 mutant for individual genes, the p value and the false discovery rates (FDRs) were calculated using the t-test and the Benjamini–Hochberg multiple testing procedure122 respectively (Supplementary Data 5). Prism 8.0 (Graphpad) was used to plot the correlations between the log2(WT/aureo1c-1) values under different light conditions and yield the Pearson’s correlation values (Fig. 2d).
To distinguish between AUREO1c-dependent and -independent genes among those regulated by high light in wild type (Fig. 3a), we defined a gene as AUREO1c-dependent if it satisfied one of the two criteria below regarding the ratio of the normalized read counts: (1) (WT-HWL)/(WT-GL) > 4 and (WT-HWL)/(aureo1c-1-HWL) > 2; (2) (WT-HWL)/(WT-GL) < 0.25 and (WT-HWL)/(aureo1c-1-HWL) < 0.5.
A gene was defined AUREO1c-independent if it satisfied one of these two criteria: (1) (WT-HWL)/(WT-GL) > 4 and (WT-HWL)/(aureo1c-1-HWL) < 1.6; (2) (WT-HWL)/(WT-GL) < 0.25 and (WT-HWL)/(aureo1c-1-HWL) > 0.625.
Enrichments of gene ontology (GO) terms among regulated genes were calculated using the software clusterProfiler123 (v4.4.4), which employs the Benjamini–Hochberg multiple testing procedure122 to derive adjusted p values (Supplementary Data 6).
Fluorescence microscopy to determine AUREO1c localization
Cells of the COMP lines or wild-type cells grown under normal light (40 µmol photons m−2 s−1 of white light) or high light (550 µmol photons m−2 s−1 of white light) were mounted with equal volumes of SlowFade Glass Soft-set Antifade Mountant (S36920, Thermo) containing 4′,6-diamidino-2-phenylindole (DAPI) and imaged on a live-cell imaging microscope (DeltaVision, GE) with an oil immersion objective (Olympus 100 × /1.40). Excitation/emission collection wavelengths used for chlorophyll autofluorescence, DAPI, and GFP were: 632 nm/679 nm, 390 nm/435 nm, and 475 nm/525 nm respectively. For each channel, the same exposure settings were applied onto cells of wild type and cells of the COMP line.
Production of recombinant His-tagged or glutathione S-transferase (GST)-tagged AUREO1c protein for in vitro experiments
To obtain His-tagged AUREO1c proteins, the cDNA sequence of AUREO1c was PCR amplified from total cDNA using primers HZ-1043 and HZ-1044 (Supplementary Data 1) and cloned into the pET28a vector (69864, Novagen) by homologous recombination using the ClonExpress II One Step Cloning Kit (C112, Vazyme); the vector contained a 6x-His tag at the N-terminus of the integrated gene. The completed construct was transformed into Transetta (DE3) E. coli strain (CD801, TransGen). Expression of AUREO1c was induced by supplementation with isopropyl ß-D-1-thiogalactopyranoside (IPTG) to a final concentration of 0.1 mM. Cells were broken by a high-pressure homogenizer (AH-Basic, ATS) and the recombinant AUREO1c proteins were purified by affinity chromatography on a gravity column using a Ni-NTA His binding resin (88222, Thermo). A further round of purification was achieved by fast protein liquid chromatography (ÄKTA pureTM 25, GE) using the HiTrap Q HP column (Cytiva). Finally, the proteins were concentrated by ultrafiltration using Amicon MWCO 30 kDa Ultra tubes (UFC903008, Millipore). Protein concentrations were determined using the Bradford method124.
To obtain GST-tagged AUREO1c proteins, the cDNA sequence of AUREO1c was cloned using primers HZ-1387 and HZ-1388 into pGEX4T-1, also by homologous recombination. Induction of protein expression, cell lysis, and protein concentration measurements were performed as described above for His-tagged AUREO1c proteins. Purification of GST-AUREO1c was conducted using the GSTSep Glutathione MagBeads (20562ES08, Yeasen). Unfused GST was purified to be used as a negative control in experiments afterwards.
In vitro analyses of AUREO1c responses to blue light
Recombinant His-tagged AUREO1c proteins were diluted by using 1 × phosphate-buffered saline (PBS) to a concentration of 5 µM and incubated in the dark or under blue light (200 µmol photons m−2 s−1) for 60 min. They were then crosslinked by the supplementation of bis(sulfosuccinimidyl)suberate (BS3) to a final concentration of 5 µM. After 60 min of incubation on ice, the crosslinking was quenched by the addition of Tris to 50 mM. Incubation continued for 15 min before further analyses.
One fraction of the samples was loaded onto the ÄKTA pureTM 25 platform for size-exclusion chromatography (SEC; Fig. 2e), using the Superdex 200 Increase 10/300 GL column. Absorbance at 280 nm was recorded.
Another fraction was subjected to ultra high-performance liquid chromatography-native mass spectrometry (UHPLC–nMS) analysis (Supplementary Fig. 15) on the Thermo Vanquish UHPLC system coupled to a Thermo Q Exactive UHMR mass spectrometer. The LC separation was carried out on a Waters BEH SEC column (2.1 × 150 mm, 1.7 μm) at room temperature, and isocratic separation was implemented using 100 mM ammonium acetate as the mobile phase with the flow rate at 0.2 ml/min. The MS parameters used were as follows: capillary voltage, 3500 V in positive mode; resolution, 12,500; SID voltage, 50 eV; capillary temperature, 275 °C; sheath gas, 20; aux gas, 5. Full scan spectra were acquired with a mass range from 1000 to 8000 m/z, and the molecular weight range of the protein was calculated manually.
For measurements of absorption properties, recombinant His-tagged AUREO1c proteins were first purified by Ni2+-column affinity purification and then further purified using the Superdex 200 column on the ÄKTA pureTM 25system. The fraction containing AUREO1c was diluted by using 1 × HEPES buffer (200 mM NaCl contained, pH: 7.9) to a concentration of 5 µM and incubated in darkness or under blue light (1000 µmol photons m−2 s−1) for 15 min. Then absorption spectra were measured on a microplate reader (Thermo Varioskan LUX).
DNA affinity purification sequencing (DAP-Seq)
DAP-seq and sequencing read alignment were performed as described in a published protocol125, with minor modifications. Genomic DNA from wild-type cells was fragmented using a sonifier (ME220, Covaris), with the following parameters: “duty factor”, 15%; “peak power”, 50; “cycles/burst”, 200; “duration”: 200 s. The end-repair reactions, A-tailing reactions, and adapter ligation reactions were carried out using the Fast DNA End Repair kit (K0771, Thermo), the DNA Polymerase I Klenow Fragment exo- kit (N105, Vazyme), and the T4 DNA Ligase kit (M1804, Promega), respectively, according to the manufacturer’s instructions. The adapter for ligation was generated by annealing of primers HZ-1795 and HZ-1796. After ligation, the mixture was purified using the GeneJET PCR Purification Kit (K0701, Thermo). For each DNA affinity purification reaction, 500 ng of ligation products were incubated with 5 μg of GST-AUREO1c proteins or GST proteins under room temperature. The incubation lasted for 1 h. The beads were washed 6 times with PBS supplemented with IGEPAL CA-630 (I8896, Sigma; final concentration: 0.005%) and additionally washed twice with PBS. The eluted DNA was used to construct the sequencing library by PCR using primers HZ-1797/HZ-1800 (for GST) or HZ-1799/HZ-1800 (for GST-AUREO1c), followed by gel purification using the E.Z.N.A. Gel Extraction Kit (D2500, Omega). Sequencing of the libraries were performed by Novogene Co on the Illumina Nova-PE150 platform. The reads are illustrated using the Integrative Genomics Viewer (IGV) software126 (Fig. 3e).
Electrophoresis mobility shift assay (EMSA)
Promoter sequences of LHCX2, LHCX3, and the alpha tubulin gene were PCR amplified from genomic DNA of P. tricornutum, with one of the primers biotinylated at the 5’ terminus (Supplementary Data 1), and then purified. Incubation of recombinant His-tagged AUREO1c with the probes, mobility analysis of probes, and their visualization was performed using the Light Shift Chemiluminescent EMSA Kit (20148, Thermo) according to the manufacturer’s instructions. Control samples that did not contain AUREO1c proteins were included to show the mobility of free probes. In additional samples, non-biotinylated probes were added to compete for binding by AUREO1c.
Accession numbers
For P. tricornutum genes, Phatr3 IDs of the ASM15095v2 genome assembly are provided as follows. AUREO1a: Phatr3_ J8113; AUREO1b: Phatr3_ J15977; AUREO1c: Phatr3_ J51933; LHCX2: Phatr3_EG02404 (Phatr2_54065); LHCX3: Phatr3_ J44733; RPS: Phatr3_ J10847; EF1α: Phatr3_ J18475; α tubulin: Phatr3_ J54534. For C. reinhardtii genes, v5.6 IDs of the ABCN02000000 genome assembly are provided as follows. LHCSR3.1: Cre08.g367500; LHCSR3.2: Cre08.g367400; RACK1, Cre06.g278222.
Raw transcriptome files from the sequencing platform have been uploaded to NCBI Sequence Read Archive (SRA; https://www.ncbi.nlm.nih.gov/sra). Accession numbers for each individual transcriptome are provided in Supplementary Data 3. No custom code was used to derive major conclusions in this study.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All generated and analyzed data from this study are included in the main figures, Supplementary Information, or have been uploaded to public repositories. All unique materials are readily available from the corresponding author on request. RNA sequencing data generated in this study have been deposited in the NCBI database under accession code. The accession number of sequence Read Archive (SRA) data are SAMN34393917, SAMN34393918, SAMN34393919 (for WT in growth light), SAMN34393932, SAMN34393933, SAMN34393934 (for aureo1c-1 in growth light), SAMN34393911, SAMN34393912, SAMN34393913 (for WT in high red light), SAMN34393926, SAMN34393927, SAMN34393928 (for aureo1c-1 in high red light), SAMN34393914, SAMN34393915, SAMN34393916 (for WT in high white light), SAMN34393929, SAMN34393930, SAMN34393931 (for aureo1c-1 in high white light), SAMN34393905, SAMN34393906, SAMN34393907 (for WT in high blue light 1), SAMN34393920, SAMN34393921, SAMN34393922 (for aureo1c-1 in high blue light 1), SAMN34393908, SAMN34393909, SAMN34393910 (for WT in high blue light 2), SAMN34393923, SAMN34393924, SAMN34393925 (for aureo1c-1 in high blue light 2). Accession numbers for each individual transcriptome are also provided in Supplementary Data 3. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE127 partner repository with the dataset identifier PXD045342 for Phaeodactylum tricornutum, PXD045344 for Chlamydomonas reinhardtii carried out in TAP medium, and PXD050435 for Chlamydomonas reinhardtii carried out in HS medium. The DAP-seq data used in this study are available in the NCBI database under accession code SAMN37389769 (incubating AUREO1c-GST fusion protein with DNA in vitro) and SAMN37389768 (incubating GST protein with DNA in vitro, control). Source data are provided with this paper.
References
Katz, M. E., Finkel, Z. V., Grzebyk, D., Knoll, A. H. & Falkowski, P. G. Evolutionary trajectories and biogeochemical impacts of marine eukaryotic phytoplankton. Annu. Rev. Ecol. Evol. Syst. 35, 523–556 (2004).
Ibarbalz, F. M. et al. Global trends in marine plankton diversity across kingdoms of life. Cell 179, 1084–1097.e21 (2019).
Field, C. B., Behrenfeld, M. J., Randerson, J. T. & Falkowski, P. Primary production of the biosphere: integrating terrestrial and oceanic components. Science 281, 237–240 (1998).
Seth, K. et al. Bioprospecting of fucoxanthin from diatoms—challenges and perspectives. Algal Res. 60, 102475 (2021).
Wang, W. et al. Structural basis for blue-green light harvesting and energy dissipation in diatoms. Science 363, https://doi.org/10.1126/science.aav0365 (2019).
Büchel, C. et al. in The Molecular Life of Diatoms (eds Falciatore, A. & Mock, T.) 397–422 (Springer International Publishing, 2022).
Croce, R. & van Amerongen, H. Natural strategies for photosynthetic light harvesting. Nat. Chem. Biol. 10, 492–501 (2014).
Moejes, F. W. et al. A systems-wide understanding of photosynthetic acclimation in algae and higher plants. J. Exp. Bot. 68, 2667–2681 (2017).
Bassi, R. & Dall’Osto, L. Dissipation of light energy absorbed in excess: the molecular mechanisms. Annu. Rev. Plant Biol. 72, 47–76 (2021).
Ruban, A. V. Nonphotochemical chlorophyll fluorescence quenching: mechanism and effectiveness in protecting plants from photodamage. Plant Physiol. 170, 1903–1916 (2016).
Muller, P., Li, X. P. & Niyogi, K. K. Non-photochemical quenching. A response to excess light energy. Plant Physiol. 125, 1558–1566 (2001).
Krause, G. H., Vernotte, C. & Briantais, J.-M. Photoinduced quenching of chlorophyll fluorescence in intact chloroplasts and algae. Resolution into two components. Biochim. Biophys. Acta 679, 116–124 (1982).
Niyogi, K. K., Grossman, A. R. & Bjorkman, O. Arabidopsis mutants define a central role for the xanthophyll cycle in the regulation of photosynthetic energy conversion. Plant Cell 10, 1121–1134 (1998).
Gagne, G. & Guertin, M. The early genetic response to light in the green unicellular alga Chlamydomonas eugametos grown under light/dark cycles involves genes that represent direct responses to light and photosynthesis. Plant Mol. Biol. 18, 429–445 (1992).
Li, X. P. et al. A pigment-binding protein essential for regulation of photosynthetic light harvesting. Nature 403, 391–395 (2000).
Peers, G. et al. An ancient light-harvesting protein is critical for the regulation of algal photosynthesis. Nature 462, 518–521 (2009).
Bailleul, B. et al. An atypical member of the light-harvesting complex stress-related protein family modulates diatom responses to light. Proc. Natl Acad. Sci. USA 107, 18214–18219 (2010).
Giovagnetti, V. & Ruban, A. V. The evolution of the photoprotective antenna proteins in oxygenic photosynthetic eukaryotes. Biochem. Soc. Trans. 46, 1263–1277 (2018).
Dinc, E. et al. LHCSR1 induces a fast and reversible pH-dependent fluorescence quenching in LHCII in Chlamydomonas reinhardtii cells. Proc. Natl Acad. Sci. USA 113, 7673–7678 (2016).
Buck, J. M. et al. Lhcx proteins provide photoprotection via thermal dissipation of absorbed light in the diatom Phaeodactylum tricornutum. Nat. Commun. 10, 4167 (2019).
Buck, J. M., Kroth, P. G. & Lepetit, B. Identification of sequence motifs in Lhcx proteins that confer qE-based photoprotection in the diatom Phaeodactylum tricornutum. Plant J. 108, 1721–1734 (2021).
Taddei, L. et al. Dynamic changes between two LHCX-related energy quenching sites control diatom photoacclimation. Plant Physiol. 177, 953–965 (2018).
Buck, J. M., Wünsch, M., Schober, A. F., Kroth, P. G. & Lepetit, B. Impact of Lhcx2 on acclimation to low iron conditions in the diatom phaeodactylum tricornutum. Front. Plant Sci. 13, 841058 (2022).
MacIntyre, H. L., Kana, T. M. & Geider, R. J. The effect of water motion on short-term rates of photosynthesis by marine phytoplankton. Trends Plant Sci. 5, 12–17 (2000).
Strzepek, R. F. & Harrison, P. J. Photosynthetic architecture differs in coastal and oceanic diatoms. Nature 431, 689–692 (2004).
Ruban, A. et al. The super-excess energy dissipation in diatom algae: comparative analysis with higher plants. Photosynth. Res. 82, 165–175 (2004).
Litchman, E. Growth rates of phytoplankton under fluctuating light. Freshw. Biol. 44, 223–235 (2000).
Richardson, K., Beardall, J. & Raven, J. A. Adaptation of unicellular algae to irradiance: an analysis of strategies. Freshw. Biol. 93, 157–191 (1983).
Wagner, H., Jakob, T. & Wilhelm, C. Balancing the energy flow from captured light to biomass under fluctuating light conditions. N. Phytol. 169, 95–108 (2006).
Zhou, L. et al. Photosynthesis acclimation under severely fluctuating light conditions allows faster growth of diatoms compared with dinoflagellates. BMC Plant Biol. 21, 164 (2021).
Lavaud, J. Fast regulation of photosynthesis in diatoms: mechanisms, evolution and ecophysiology. Funct. Plant Sci. Biotechnol. 1, 267–287 (2007).
Lavaud, J., Rousseau, B. & Etienne, A. L. General features of photoprotection by energy dissipation in planktonic diatoms (Bacillariophyceae). J. Phycol. 40, 130–137 (2004).
Lohr, M. & Wilhelm, C. Xanthophyll synthesis in diatoms: quantification of putative intermediates and comparison of pigment conversion kinetics with rate constants derived from a model. Planta 212, 382–391 (2001).
Olaizola, M., La Roche, J., Kolber, Z. & Falkowski, P. G. Non-photochemical fluorescence quenching and the diadinoxanthin cycle in a marine diatom. Photosynth. Res. 41, 357–370 (1994).
Lepetit, B. et al. The diatom Phaeodactylum tricornutum adjusts nonphotochemical fluorescence quenching capacity in response to dynamic light via fine-tuned Lhcx and xanthophyll cycle pigment synthesis. N. Phytol. 214, 205–218 (2017).
Zhu, S. H. & Green, B. R. Photoprotection in the diatom Thalassiosira pseudonana: role of LI818-like proteins in response to high light stress. Biochim. Biophys. Acta 1797, 1449–1457 (2010).
Giovagnetti, V. et al. Biochemical and molecular properties of LHCX1, the essential regulator of dynamic photoprotection in diatoms. Plant Physiol. 188, 509–525 (2022).
Nymark, M. et al. An integrated analysis of molecular acclimation to high light in the marine diatom Phaeodactylum tricornutum. PLoS ONE 4, e7743 (2009).
Taddei, L. et al. Multisignal control of expression of the LHCX protein family in the marine diatom Phaeodactylum tricornutum. J. Exp. Bot. 67, 3939–3951 (2016).
Ruiz-Sola, M. A. et al. Light-independent regulation of algal photoprotection by CO(2) availability. Nat. Commun. 14, 1977 (2023).
Chen, M., Chory, J. & Fankhauser, C. Light signal transduction in higher plants. Annu. Rev. Genet. 38, 87–117 (2004).
Christie, J. M. Phototropin blue-light receptors. Annu. Rev. Plant Biol. 58, 21–45 (2007).
Jarillo, J. A. et al. Phototropin-related NPL1 controls chloroplast relocation induced by blue light. Nature 410, 952–954 (2001).
Kagawa, T. et al. Arabidopsis NPL1: a phototropin homolog controlling the chloroplast high-light avoidance response. Science 291, 2138–2141 (2001).
Petroutsos, D. et al. A blue-light photoreceptor mediates the feedback regulation of photosynthesis. Nature 537, 563–566 (2016).
Allorent, G. et al. UV-B photoreceptor-mediated protection of the photosynthetic machinery in Chlamydomonas reinhardtii. Proc. Natl Acad. Sci. USA 113, 14864–14869 (2016).
Tilbrook, K. et al. UV-B perception and acclimation in Chlamydomonas reinhardtii. Plant Cell 28, 966–983 (2016).
Agarwal, A., Levitan, O., Cruz de Carvalho, H. & Falkowski, P. G. Light-dependent signal transduction in the marine diatom Phaeodactylum tricornutum. Proc. Natl Acad. Sci. USA 120, e2216286120 (2023).
Seydoux, C. et al. Impaired photoprotection in Phaeodactylum tricornutum KEA3 mutants reveals the proton regulatory circuit of diatoms light acclimation. N. Phytol. 234, 578–591 (2022).
Lepetit, B. et al. High light acclimation in the secondary plastids containing diatom Phaeodactylum tricornutum is triggered by the redox state of the plastoquinone pool. Plant Physiol. 161, 853–865 (2013).
Yang, W. et al. Knockout of a diatom cryptochrome by CRISPR/Cas9 causes an increase in light-harvesting protein levels and accumulation of fucoxanthin. Algal Res. 66, 102822 (2022).
Mann, M., Serif, M., Jakob, T., Kroth, P. G. & Wilhelm, C. PtAUREO1a and PtAUREO1b knockout mutants of the diatom Phaeodactylum tricornutum are blocked in photoacclimation to blue light. J. Plant Physiol. 217, 44–48 (2017).
Mann, M. et al. The aureochrome photoreceptor PtAUREO1a is a highly effective blue light switch in diatoms. iScience 23, 101730 (2020).
Coesel, S. et al. Diatom PtCPF1 is a new cryptochrome/photolyase family member with DNA repair and transcription regulation activity. EMBO Rep. 10, 655–661 (2009).
Juhas, M. et al. A novel cryptochrome in the diatom Phaeodactylum tricornutum influences the regulation of light-harvesting protein levels. FEBS J. 281, 2299–2311 (2014).
Schellenberger Costa, B. et al. Aureochrome 1a is involved in the photoacclimation of the diatom Phaeodactylum tricornutum. PLoS ONE 8, e74451 (2013).
Fortunato, A. E. et al. Diatom phytochromes reveal the existence of far-red-light-based sensing in the ocean. Plant Cell 28, 616–628 (2016).
Nymark, M., Sharma, A. K., Sparstad, T., Bones, A. M. & Winge, P. A CRISPR/Cas9 system adapted for gene editing in marine algae. Sci. Rep. 6, 24951 (2016).
Valle, K. C. et al. System responses to equal doses of photosynthetically usable radiation of blue, green, and red light in the marine diatom Phaeodactylum tricornutum. PLoS ONE 9, e114211 (2014).
Hao, T. B. et al. Light-harvesting protein Lhcx3 is essential for high light acclimation of Phaeodactylum tricornutum. AMB Express 8, 174 (2018).
Aro, E. M., Virgin, I. & Andersson, B. Photoinhibition of photosystem II. Inactivation, protein damage and turnover. Biochim. Biophys. Acta 1143, 113–134 (1993).
Maxwell, K. & Johnson, G. N. Chlorophyll fluorescence—a practical guide. J. Exp. Bot. 51, 659–668 (2000).
Jaubert, M. et al. in The Molecular Life of Diatoms (eds Falciatore, A. & Mock, T.) 607–640 (Springer International Publishing, 2022).
Kroth, P. G., Wilhelm, C. & Kottke, T. An update on aureochromes: phylogeny—mechanism—function. J. Plant Physiol. 217, 20–26 (2017).
Takahashi, F. et al. AUREOCHROME, a photoreceptor required for photomorphogenesis in stramenopiles. Proc. Natl Acad. Sci. USA 104, 19625–19630 (2007).
Poliner, E. et al. Aureochromes maintain polyunsaturated fatty acid content in Nannochloropsis oceanica. Plant Physiol. 189, 906–921 (2022).
Zhang, P. et al. Exploring a blue-light-sensing transcription factor to double the peak productivity of oil in Nannochloropsis oceanica. Nat. Commun. 13, 1664 (2022).
Coesel, S. N. et al. Diel transcriptional oscillations of light-sensitive regulatory elements in open-ocean eukaryotic plankton communities. Proc. Natl Acad. Sci. USA 118, https://doi.org/10.1073/pnas.2011038118 (2021).
Huysman, M. J. et al. AUREOCHROME1a-mediated induction of the diatom-specific cyclin dsCYC2 controls the onset of cell division in diatoms (Phaeodactylum tricornutum). Plant Cell 25, 215–228 (2013).
Ait-Mohamed, O. et al. PhaeoNet: a holistic RNAseq-based portrait of transcriptional coordination in the model diatom Phaeodactylum tricornutum. Front. Plant Sci. 11, 590949 (2020).
Schellenberger Costa, B. et al. Blue light is essential for high light acclimation and photoprotection in the diatom Phaeodactylum tricornutum. J. Exp. Bot. 64, 483–493 (2013).
Kasahara, M. et al. Photochemical properties of the flavin mononucleotide-binding domains of the phototropins from Arabidopsis, rice, and Chlamydomonas reinhardtii. Plant Physiol. 129, 762–773 (2002).
Bannister, S., Bohm, E., Zinn, T., Hellweg, T. & Kottke, T. Arguments for an additional long-lived intermediate in the photocycle of the full-length aureochrome 1c receptor: a time-resolved small-angle X-ray scattering study. Struct. Dyn. 6, 034701 (2019).
Bulankova, P. et al. Mitotic recombination between homologous chromosomes drives genomic diversity in diatoms. Curr. Biol. 31, 3221–3232.e9 (2021).
Johzuka-Hisatomi, Y., Terada, R. & Iida, S. Efficient transfer of base changes from a vector to the rice genome by homologous recombination: involvement of heteroduplex formation and mismatch correction. Nucleic Acids Res. 36, 4727–4735 (2008).
Redekop, P. et al. Transcriptional regulation of photoprotection in dark-to-light transition-More than just a matter of excess light energy. Sci. Adv. 8, eabn1832 (2022).
Banerjee, A., Herman, E., Kottke, T. & Essen, L. O. Structure of a native-like aureochrome 1a LOV ___domain dimer from Phaeodactylum tricornutum. Structure 24, 171–178 (2016).
Banerjee, A. et al. Allosteric communication between DNA-binding and light-responsive domains of diatom class I aureochromes. Nucleic Acids Res. 44, 5957–5970 (2016).
Heintz, U. & Schlichting, I. Blue light-induced LOV ___domain dimerization enhances the affinity of Aureochrome 1a for its target DNA sequence. Elife 5, e11860 (2016).
Herman, E., Sachse, M., Kroth, P. G. & Kottke, T. Blue-light-induced unfolding of the Jalpha helix allows for the dimerization of aureochrome-LOV from the diatom Phaeodactylum tricornutum. Biochemistry 52, 3094–3101 (2013).
Hisatomi, O., Nakatani, Y., Takeuchi, K., Takahashi, F. & Kataoka, H. Blue light-induced dimerization of monomeric aureochrome-1 enhances its affinity for the target sequence. J. Biol. Chem. 289, 17379–17391 (2014).
Briggs, W. R. & Christie, J. M. Phototropins 1 and 2: versatile plant blue-light receptors. Trends Plant Sci. 7, 204–210 (2002).
Koziol, A. G. et al. Tracing the evolution of the light-harvesting antennae in chlorophyll a/b-containing organisms. Plant Physiol. 143, 1802–1816 (2007).
O’Malley, R. C. et al. Cistrome and epicistrome features shape the regulatory DNA landscape. Cell 165, 1280–1292 (2016).
Aihara, Y., Fujimura-Kamada, K., Yamasaki, T. & Minagawa, J. Algal photoprotection is regulated by the E3 ligase CUL4-DDB1(DET1). Nat. Plants 5, 34–40 (2019).
Tokutsu, R., Fujimura-Kamada, K., Matsuo, T., Yamasaki, T. & Minagawa, J. The CONSTANS flowering complex controls the protective response of photosynthesis in the green alga Chlamydomonas. Nat. Commun. 10, 4099 (2019).
Gabilly, S. T. et al. Regulation of photoprotection gene expression in Chlamydomonas by a putative E3 ubiquitin ligase complex and a homolog of CONSTANS. Proc. Natl Acad. Sci. USA 116, 17556–17562 (2019).
Strassert, J. F. H., Irisarri, I., Williams, T. A. & Burki, F. A molecular timescale for eukaryote evolution with implications for the origin of red algal-derived plastids. Nat. Commun. 12, 1879 (2021).
Ghusinga, K. R., Jones, R. D., Jones, A. M. & Elston, T. C. Molecular switch architecture determines response properties of signaling pathways. Proc. Natl Acad. Sci. USA 118, https://doi.org/10.1073/pnas.2013401118 (2021).
Armbruster, U. et al. Ion antiport accelerates photosynthetic acclimation in fluctuating light environments. Nat. Commun. 5, 5439 (2014).
Kromdijk, J. et al. Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science 354, 857–861 (2016).
Christie, J. M. & Zurbriggen, M. D. Optogenetics in plants. N. Phytol. 229, 3108–3115 (2021).
Deisseroth, K. Optogenetics: 10 years of microbial opsins in neuroscience. Nat. Neurosci. 18, 1213–1225 (2015).
Boyden, E. S., Zhang, F., Bamberg, E., Nagel, G. & Deisseroth, K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci. 8, 1263–1268 (2005).
Chen, X. et al. An extraordinary stringent and sensitive light-switchable gene expression system for bacterial cells. Cell Res. 26, 854–857 (2016).
Li, X. et al. A single-component light sensor system allows highly tunable and direct activation of gene expression in bacterial cells. Nucleic Acids Res. 48, e33 (2020).
Kristensen, T. N., Ketola, T. & Kronholm, I. Adaptation to environmental stress at different timescales. Ann. N. Y. Acad. Sci. 1476, 5–22 (2020).
Litchman, E. & Klausmeier, C. A. Dissipation of light energy absorbed in excess: the molecular mechanisms. Ann. Rev. Ecol. Evol. Syst. 39, 615–639 (2008).
Morales, A. & Kaiser, E. Photosynthetic acclimation to fluctuating irradiance in plants. Front. Plant Sci. 11, 268 (2020).
Morel, A., Lazzara, L. & Gostan, J. Growth rate and quantum yield time response for a diatom to changing irradiances (energy and color). Limnol. Oceanogr. 32, 1066–1084 (1987).
Orefice, I. et al. Light-induced changes in the photosynthetic physiology and biochemistry in the diatom Skeletonema marinoi. Algal Res. 17, 1–13 (2016).
Zhang, R. et al. High-throughput genotyping of green algal mutants reveals random distribution of mutagenic insertion sites and endonucleolytic cleavage of transforming DNA. Plant Cell 26, 1398–1409 (2014).
Kropat, J. et al. A revised mineral nutrient supplement increases biomass and growth rate in Chlamydomonas reinhardtii. Plant J. 66, 770–780 (2011).
Sueoka, N. Mitotic replication of deoxyribonucleic acid in chlamydomonas reinhardi. Proc. Natl Acad. Sci. USA 46, 83–91 (1960).
Concordet, J. P. & Haeussler, M. CRISPOR: intuitive guide selection for CRISPR/Cas9 genome editing experiments and screens. Nucleic Acids Res. 46, W242–W245 (2018).
Slattery, S. S. et al. An expanded plasmid-based genetic toolbox enables Cas9 genome editing and stable maintenance of synthetic pathways in Phaeodactylum tricornutum. ACS Synth. Biol. 7, 328–338 (2018).
Karas, B. J. et al. Designer diatom episomes delivered by bacterial conjugation. Nat. Commun. 6, 6925 (2015).
Bai, Y. et al. Green diatom mutants reveal an intricate biosynthetic pathway of fucoxanthin. Proc. Natl Acad. Sci. USA 119, e2203708119 (2022).
Apt, K. E., Kroth-Pancic, P. G. & Grossman, A. R. Stable nuclear transformation of the diatom Phaeodactylum tricornutum. Mol. Gen. Genet. 252, 572–579 (1996).
Zhang, C. & Hu, H. High-efficiency nuclear transformation of the diatom Phaeodactylum tricornutum by electroporation. Mar. Genomics 16, 63–66 (2014).
Thompson, A. et al. Tandem mass tags: a novel quantification strategy for comparative analysis of complex protein mixtures by MS/MS. Anal. Chem. 75, 1895–1904 (2003).
Bilger, W. & Bjorkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 25, 173–185 (1990).
Fu, W., Magnusdottir, M., Brynjolfson, S., Palsson, B. O. & Paglia, G. UPLC-UV-MS(E) analysis for quantification and identification of major carotenoid and chlorophyll species in algae. Anal. Bioanal. Chem. 404, 3145–3154 (2012).
Zhu, M., Mori, M., Hwa, T. & Dai, X. Disruption of transcription-translation coordination in Escherichia coli leads to premature transcriptional termination. Nat. Microbiol. 4, 2347–2356 (2019).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402–408 (2001).
Chen, S., Zhou, Y., Chen, Y. & Gu, J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 34, i884–i890 (2018).
Bowler, C. et al. The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 456, 239–244 (2008).
Kim, D., Paggi, J. M., Park, C., Bennett, C. & Salzberg, S. L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 37, 907–915 (2019).
Li, H. et al. The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079 (2009).
Liao, Y., Smyth, G. K. & Shi, W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930 (2014).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Benjamini, Y. & Hochberg, Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. J. Biol. Chem. 57, 289–300 (1995).
Wu, T. et al. clusterProfiler 4.0: a universal enrichment tool for interpreting omics data. Innovation 2, 100141 (2021).
Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 72, 248–254 (1976).
Bartlett, A. et al. Mapping genome-wide transcription-factor binding sites using DAP-seq. Nat. Protoc. 12, 1659–1672 (2017).
Robinson, J. T. et al. Integrative genomics viewer. Nat. Biotechnol. 29, 24–26 (2011).
Perez-Riverol, Y. et al. The PRIDE database resources in 2022: a hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 50, D543–d552 (2022).
Acknowledgements
The authors thank these colleagues for helpful discussion: Hongtao Liu from Shenzhen University; Congming Lu from Shandong Agricultural University; Yizhong Yuan from the Carnegie Institution for Science; Deqiang Duanmu and Longlong Wang from Huazhong Agricultural University; Wenbin Zhou, Zefu Lu, Pan Liu, and Shaobo Wei from the Chinese Academy of Agricultural Sciences; Qiguang Xie from Henan University; Xiaojie Pang from Chinese Academy of Sciences; Mingzhu Fan, Yanlei Feng, Xiangdong Fu, Yinping Gao, Peng Liu, Fang Xiao, Yanxiao Zhang, and Jia Zheng from Westlake University. We thank Du Cao (Westlake University) for proofreading the manuscript. We also acknowledge the Instrumentation and Service Center for Molecular Sciences and the Biomedical Research Core Facilities at Westlake University for technical support. This work was supported by fundings listed below: National Key R&D Program of China (2019YFA0906300 to X.L.), National Natural Science Foundation of China (32150022 to X.L., 42106114 to H.Z., 42206115 to T.C.), National Key R&D Program of China (2022YFC3401800 to T.C.), Zhejiang Province Key R&D Program (2023SDXHDX0002 to X.L.), Zhejiang Natural Science Foundation (LR20C020002 to X.L.), Zhejiang Provincial Key Laboratory Construction Project (to X.L.), Westlake Research Center for Industries of the Future (WU2023C002 to X.L.), Westlake Education Foundation (WU2023B002 to X.L.), and the Westlake Center for Genome Editing (to X.L.).
Author information
Authors and Affiliations
Contributions
H.Z. and X.L. conceived the project. H.Z., L.T., and X.L. designed the research. H.Z., K.G., X.X., M.Z., T.C., Y.Y., J.S., J.C., J.Z., Y.J., and S.F. performed experiments and bioinformatic analyses; X.L. wrote the manuscript with contributions from all authors.
Corresponding author
Ethics declarations
Competing interests
X.L., H.Z., K.G., X.X., and T.C. have submitted a Chinese patent application for this work presented in the manuscript. The application number is 202410675502.8. The other authors declare that they have no competing interests.
Peer review
Peer review information
Nature Communications thanks Johann Lavaud, Dimitris Petroutsos and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhang, H., Xiong, X., Guo, K. et al. A rapid aureochrome opto-switch enables diatom acclimation to dynamic light. Nat Commun 15, 5578 (2024). https://doi.org/10.1038/s41467-024-49991-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-024-49991-7
This article is cited by
-
Uridine diphosphate glucose confers oxidative stress tolerance in microalgae
BMC Plant Biology (2025)
