
Abstract
The endosymbiont Candidatus Azoamicus ciliaticola was proposed to generate ATP for its eukaryotic host, an anaerobic ciliate of the Plagiopylea class, fulfilling a function analogous to mitochondria in other eukaryotic cells. The discovery of this respiratory endosymbiosis has major implications for both evolutionary history and ecology of microbial eukaryotes. However, with only a single species described, knowledge of its environmental distribution and diversity is limited. Here we report four complete, circular metagenome assembled genomes (cMAGs) representing respiratory endosymbionts inhabiting groundwater in California, Ohio, and Germany. These cMAGs form two lineages comprising a monophyletic clade within the uncharacterized gammaproteobacterial order UBA6186, enabling evolutionary analysis of their key protein complexes. Strikingly, all four cMAGs encode a cytochrome cbb3 oxidase, which indicates that these endosymbionts have the capacity for aerobic respiration. Accordingly, we detect these respiratory endosymbionts in diverse habitats worldwide, thus further expanding the ecological scope of this respiratory symbiosis.
Similar content being viewed by others


Introduction
Host beneficial endosymbionts (HBEs) are widespread in eukaryotes, and often evolve from a parasitic or pathogenic ancestor1. The best studied HBEs are nutritional symbionts of insects that provide their hosts with essential molecules (such as vitamins or cofactors), which the hosts cannot synthesize or obtain from their diet2,3. Another example are defensive endosymbionts that synthesize compounds that protect the host or its offspring against predators or pathogens4,5. Beyond insect hosts, intracellular endosymbionts also perform a wide range of beneficial functions in protists, such as photosynthesis, scavenging of hydrogen or other metabolic products, or complementing catabolic pathways (reviewed in ref.6). The recently discovered protist endosymbiont Ca. A. ciliaticola fulfills yet another function, using a denitrifying respiratory chain to generate ATP that can be supplied to its ciliate host7. The Plagiopylean ciliate host of Ca. A. ciliaticola was proposed to harbor only metabolically reduced organelles known as mitochondrion-related organelles (MROs)8,9, possibly in the form of hydrogenosomes. Thus the host is likely incapable of (aerobic) respiration, although it might still be able to generate ATP in the MROs, or through substrate level phosphorylation in the cytoplasm7,10. The endosymbiont Ca. A. ciliaticola also lacks the capability for aerobic respiration, restricting the ecological niche of its ciliate host to permanently anoxic habitats, such as the hypolimnion of a meromictic lake. Interestingly, virtually all organisms capable of denitrification are facultative anaerobes11,12, and therefore it was hypothesized that Ca. A. ciliaticola evolved from a predecessor that was capable of both aerobic respiration and denitrification7. To investigate this possibility, we searched for genomes of endosymbionts related to Ca. A. ciliaticola in publicly available environmental metagenomic sequencing datasets.
In this work we describe the discovery for four additional closed genomes representing respiratory endosymbionts. The five genomes form a monophyletic clade and have strongly conserved gene content and genomic features, indicating a shared function in their host. Using the expanded genomic coverage, we confidently place this endosymbiont clade in the UBA6186 order, which we accordingly propose to rename to Azoamicales. We show that the four newly retrieved genomes encode the capability for aerobic respiration in addition to respiration of nitrogen oxides, and that the genes required for both were acquired horizontally. Finally, we use the expanded genome coverage to show that the respiratory endosymbionts are globally distributed.
Results
Discovery and genomic features of novel endosymbiont genomes
We used the tlcA gene, which encodes for an ATP/ADP transporter that is crucial for the proposed ATP supplying function of Ca. A. ciliaticola, to identify genomes of putative respiratory endosymbionts. Using this strategy, we recovered one complete circular cMAG originating from groundwater samples taken in California13 and assembled three complete circular cMAGs from groundwater samples of a carbonate-rock-fracture aquifer taken in Germany (1 cMAG)14,15 and groundwater samples from Ohio (2 cMAGs)16. All four cMAGs share genomic features characteristic of obligate endosymbionts, including extreme genome reduction (284–373 kb genome size), low GC content (20–26 %) and a high protein coding density (91–92 %) (Fig. 1, Supplementary Table 1). Additionally, all four cMAGs contain a complete respiratory denitrification pathway (Fig. 1, Supplementary Data 2), but have undergone extensive loss of genes for the biosynthesis of vitamins, amino acids, and other essential metabolites (Supplementary Data 3). These genomic similarities strongly suggest that, like Ca. A ciliaticola, these cMAGs represent obligate respiratory endosymbionts that perform denitrification.

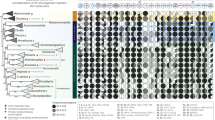
Concatenated marker gene phylogeny of the gammaproteobacterial orders Berkiellales, JABCZS01 and UBA6186. The expanded section shows the UBA6186 order, including the Azoamicaceae. The tree annotations indicate, from left to right, MAG completeness as estimated by Anvi’o (in %), genome length (in Mbp), presence (filled squares) or absence (empty squares) of the denitrification pathway, the electron transport chain complexes NADH dehydrogenase (I), succinate dehydrogenase (II), bc1 complex (III), cytochrome bd oxidase (IV - bd), cytochrome caa3 oxidase (IV - A), cytochrome cbb3 oxidase (IV - C), and ATP synthase (V), and presence of at least one copy of the tlcA gene encoding an ATP/ADP transporter. The genomic features of Azoamicaceae are indicated in red, while the features of the other UBA6186 genomes are shown in gray. Sequences were obtained from genomes included in the genome taxonomy database (GTDB) and genomic catalog of Earth’s microbiomes (GEM) databases. Black circles on the branches indicate bootstrap values higher than 90%.
The four cMAGs (hereafter: genomes) form a monophyletic clade with Ca. A. ciliaticola, within the UBA6186 order of the Gammaproteobacteria (Fig. 1, Supplementary Fig. 1, Supplementary Notes). The groundwater genomes belong to two distinct clades, with the two Ohio genomes forming a lineage with the lacustrine Ca. A. ciliaticola, and the California and Germany genomes forming a novel lineage (Fig. 1). Based on the pairwise average amino acid identity (AAI) and average nucleotide identity (ANI) values17,18 between the five genomes (Supplementary Data 4) we propose that they represent five species grouped in two genera within the Candidatus Azoamicaceae family (hereafter: Azoamicaceae). The two genera are the previously established Candidatus Azoamicus (hereafter: Azoamicus) and a novel genus, for which we propose the name Candidatus Azosocius (hereafter: Azosocius; see Supplementary Notes for discussion of species and genera delineation). We propose the names Candidatus Azoamicus viridis and Candidatus Azoamicus soli for the two Ohio genomes and Candidatus Azosocius agrarius and Candidatus Azosocius aquiferis for the California and Germany genomes, respectively. Candidatus Azosocius agrarius is the type species of the Azosocius genus.
The genomes of the Azosocius species are nearly identical in size (352 and 353 kpb for Ca. A. agrarius and Ca. A. aquiferis, respectively) and they are larger than those of Ca. A. soli (284 kpb) and Ca. A. ciliaticola (293 kbp); however, the largest genome belonged to Ca. A. viridis (374 kbp). A comparative genomic analysis of the five Azoamicaceae genomes revealed that their collective protein coding gene content amounts to only 470 genes (Fig. 2a), of which 254 are shared between all five genomes (68−87 % of the protein coding gene complement of individual genomes). This conserved set includes the genes for an electron transport chain and ATP synthase (Complexes I, II, III, and V), a complete denitrification pathway for reduction of nitrate to dinitrogen gas, ATP/ADP transporter, iron sulfur cluster biosynthesis, molybdopterin cofactor biosynthesis, and cellular information processing (Fig. 2d, Supplementary Data 2). 43 additional genes are present in multiple (2-4) genomes across both Azoamicus and Azosocius lineages (Fig. 2a), indicating ongoing loss of genes after the lineages diverged. Pseudogene prediction revealed 3-8 putative pseudogenes in the Azoamicaceae genomes (Supplementary Data 5), providing further support for ongoing genome erosion. In addition to the genes shared between the two lineages, there are 107 genes unique to Azoamicus and 66 genes unique to Azosocius (Fig. 2b), as further discussed in the Supplementary Notes. Analysis of single nucleotide variants (SNVs) revealed only 0 − 65 SNVs in the groundwater Azoamicaceae genomes (Supplementary Data 6). However, this limited genomic variability may be a consequence of the low coverage of the Azoamicaceae in the source datasets, as the more deeply sequenced metagenomes of Ca. A. ciliaticola7 showed higher variation (74 − 2856 SNVs). Nonetheless, these analyzes indicate that strain differentiation might exist in these microbial symbionts, with a so-far unknown effect on the symbiotic association with its host.

a UpSet plot showing the coding sequence (CDS) content shared between, or unique to, the individual Azoamicaceae genomes. Each vertical bar represents the number of genes in an intersecting set between the genomes. The filled circles connected by lines indicate which genomes contribute genes to the intersecting set. Sets are ordered by the number of genomes contributing, and then the number of genes in the set. Vertical bars are colored according to the broad classification of genes that can be found in Supplementary Data 2. Total coding sequence numbers for the genomes are Ca. A aquiferis 352; Ca. A agrarius 347; Ca. A soli 299; Ca. A ciliaticola 311; Ca. A viridis 378 (b, inset) Distribution of the gene content shown in panel A, summarized by genus. Aa: Azoamicus, As: Azosocius. (c, inset) Distribution of the CDS content shown in panel A summarized by the number of genomes containing each gene. (d) Key pathways in the Azoamicaceae genomes that could enable generation of ATP for the host cell from both denitrification and aerobic respiration. Boxes indicate protein complexes or pathways, with background shading corresponding to the categories in panels (a–c). Filled and empty circles indicate the presence/absence of a complex or pathway in the genome. Roman numerals in the boxes representing the electron transport chain complexes indicate: (I) NADH dehydrogenase, (II) succinate dehydrogenase, (III) bc1 complex, (IV) cytochrome cbb3 oxidase, (V) ATP synthase.
Presence and transcription of high affinity cytochrome c oxidase in groundwater Azoamicaceae
Notably, among the genes conserved in the four genomes is an operon consisting of ccoNO(Q)P, encoding for a high affinity cytochrome cbb3 oxygen reductase (complex IV)19. The cbb3 operon is complete in three of the four genomes, only Ca. A. soli appears to have lost the ccoQ gene that encodes for the small non-catalytic subunit20. The cytochrome cbb3 oxygen reductases were initially thought to be restricted to Proteobacteria, but have since also been found in genomes of organisms from other bacterial phyla21. In organisms encoding a cytochrome cbb3 oxidase, small c4 and c5 cytochromes can form a branch point in the respiratory chain, allowing electrons from complex III to flow to either the cbb3 oxidase during aerobic respiration or a nitrite reductase during denitrification. All four Azoamicaceae genomes encode the diheme cytochrome c4 that acts as the natural electron donor to the cytochome cbb3 oxidase in Vibrio cholerae22 and Neisseria meningitidis23, as well as the c5 monoheme cytochrome that shuttles electrons from complex III to nitrite reductase in Neisseria meningitidis23. These small cytochromes are also present in Ca. A. ciliaticola, which lacks the cbb3 oxidase, strongly suggesting that Ca. A. ciliaticola has lost its terminal oxidase secondarily. This is further supported by the presence of a cbb3 oxidase in closely related Ca. A. viridis and Ca. A. soli genomes from the Azoamicus genus (Fig. 1).
We confirmed the transcription of the ccoNOQP operon in Ca. A aquiferis using data from 18 groundwater metatranscriptome datasets from the Hainich Critical Zone Exploratory (CZE)24, from which the Ca. A aquiferis genome was assembled (Supplementary Data 7). Sufficient reads matching Ca. A. aquiferis genes for analysis of genome wide transcription patterns (327−1138 reads) could be recovered from seven of these datasets, which represent two wells sampled at two time points. The overall observed transcription pattern was very similar in all seven datasets (Fig. 3, Supplementary Data 7). As previously observed for Ca. A. ciliaticola, the nirK, norB, and nosZ genes were amongst the highest transcribed genes for Ca. A aquiferis in all groundwater samples, with transcription of the other genes of the denitrification pathway lower but still detectable7. The catalytic subunit of the cytochrome cbb3 oxidase (encoded by ccoN) was also consistently highly transcribed, at a similar level as e.g. the narG gene (encoding for the catalytic subunit of nitrate reductase). The ccoO and ccoP genes encoding the other core subunits of the cytochrome cbb3 oxidase were also highly transcribed in some but not all datasets (Fig. 3, Supplementary Data 5). Interestingly, high ccoNOP gene transcription was detected in samples with markedly different dissolved oxygen concentrations15. Samples from well H5-2 originate from typically anoxic groundwater in low permeable marlstones and dissolved oxygen was not detected at the time of RNA sampling25. In contrast, groundwater samples from the permeable fracture aquifer accessed at well H4-1 contained ~6 mg/L dissolved oxygen at the time of RNA sampling25, which is typical for this sampling site15. The comparably high levels of transcription of the cytochrome cbb3 oxidase operon in oxic and anoxic groundwater at both timepoints (Fig. 3) may indicate that the terminal oxidase is constitutively expressed in Ca. A. aquiferis, as previously observed for cco1 of Pseudomonas aeruginosa26.

Heatmap depicting the gene expression of 352 protein coding genes (columns) in the genome of Ca. A. aquiferis across seven metatranscriptome samples (rows) taken from anoxic groundwater of well H5-2 and oxic groundwater of well H4-1 at the Hainich critical zone exploratory (CZE) during two sampling campaigns. The genes are arranged in order according to their position in the Ca. A. aquiferis genome, with highly expressed genes and other selected genes annotated above the heatmap. The gene expression was normalized to “transcripts per thousand”, and then log2 transformed to highlight the genome-wide expression pattern. Ef-Tu: Elongation factor Tu; Ef-G: Elongation factor G; RP S12 & L31: Ribosomal proteins S12 and L31. Accession numbers of the datasets are available in the Method section.
The presence of a cytochrome cbb3 oxidase operon in Azoamicaceae symbionts suggests that like their free-living counterparts, symbiotic denitrifying bacteria also typically have the capacity to respire oxygen in addition to nitrogen oxides. However, oxygen respiration can be secondarily lost as demonstrated by the example of Ca. A. ciliaticola. In the case of Azoamicaceae symbionts, the presence and expression of a terminal oxidase raises the compelling possibility that Azoamicaceae respiratory endosymbionts respire oxygen, instead of, or in addition to, host mitochondria.
Evolutionary history of respiratory enzymes in Azoamicaceae
Given the importance of nitrate- and oxygen-respiring genes for the function of the Azoamicaceae respiratory endosymbionts, we used phylogenetic analyzes to investigate the evolutionary history of those genes. We assume vertical inheritance when Azoamicaceae genes form a well-supported clade with their UBA6186 relatives, whereas genes that are absent from the entire UBA6186 group, are considered more likely to be horizontally acquired by the Azoamicaceae. All the core complexes of the electron transport chain (complex I, II, III and V) are monophyletic and appear to be vertically inherited from the UBA6186 clade (Fig. 1, Supplementary Fig. 2). In contrast, the cytochrome cbb3 oxidase (complex IV) appears to be horizontally transferred from the Pseudomonadales, with the closest homologs present in Alcanivoraceae and Berkiellales (Fig. 4a). The horizontal acquisition of cbb3 oxidase by the Azoamicaceae is supported by the observation that none of the UBA6186 genomes, and only a single genome in the sister clade Berkiellales, contain genes for a cytochrome cbb3 oxidase. Instead, both clades typically encode a cytochrome caa3 oxidase and a cytochrome bd-type alternative oxidase, which are both absent from the five Azoamicaceae genomes (Fig. 1). The key genes encoding for the core enzymes of the denitrification pathway (narG, nirK, norB and nosZ) also appear to be horizontally acquired, from four different bacterial donor lineages (Supplementary Figs. 3–6). The four gene clusters are located at three distinct loci in the Azoamicaceae genomes, with the genes encoding for nitrite reductase and nitric oxide reductase adjacent to each other (Supplementary Data 2). The two most closely related sequences to the Azoamicaceae narG gene, encoding for the nitrate reductase, are encoded in genomes of Gammaproteobacteria of the PIVX01 and JACQPQ01 orders. However, it appears that these Gammaproteobacteria likely obtained this gene horizontally from an alphaproteobacterial lineage (Supplementary Fig. 3). The nirK gene, encoding for the nitrite reductase is most closely related to uncultured members of the Ignavibacteraceae family and was likely horizontally acquired from a member of this clade (Supplementary Fig. 4). The provenance of the nitric oxide reductase and nitrous oxide reductase cannot be conclusively established from the phylogenetic analyzes (Supplementary Figs. 5 and 6), but the Azoamicaceae norB gene (encoding for nitric oxide reductase), and to a lesser extent the nosZ gene (encoding for nitrous oxide reductase) are related to those found in predatory bacteria of the Bdellovibrionota and Myxococcota phyla, allowing for the possibility that these taxa might have acted as intermediate carriers of these genes. Interestingly, these phylogenetic analyses of the origin of the respiratory genes suggest that the evolutionary model proposed by Graf et al.7 likely needs to be revised. Unlike originally suggested, the denitrification pathway was most likely absent from the predecessor of the Azoamicaceae and only acquired laterally at a later stage. It is possible that the capability to respire nitrogen oxides in addition to oxygen provided the host a competitive advantage for the colonization of micro-oxic and anoxic niches.

a Phylogenetic tree based on amino acid sequences of the catalytic subunit of the cbb3 cytochrome c oxidase (ccoN). Color strip indicates the GTDB assigned order within the class Gammaproteobacteria (or the phylum Myxococcota_A) of the respective genome containing the ccoN gene. Collapsed wedges contain gammaproteobacterial sequences primarily originating from Pseudomonales and Burkholderiales. b Phylogenetic tree based on amino acid sequences of the ATP/ADP transporter tlcA showing three independent origins for the Azoamicaceae genes. Both Azosocius genomes contain two copies (As1 & As2), and the three Azoamicus genomes contain third distinct tlcA copy (Aa). Color strip indicates the GTDB assigned taxonomy at the phylum or class (for Pseudomonadota) of the genome containing the tlcA gene. Sequences were obtained from genomes included in the genome taxonomy database and genomic catalog of Earth’s microbiomes databases. Black circles on the branches indicate bootstrap values higher than 80% (a) or 90% (b).
Unlike the respiratory genes discussed above, the tlcA gene encoding an ATP/ADP transporter, is not monophyletic in the Azoamicaceae (Fig. 4b) and seems to have been acquired by the Azoamicaceae or the larger UBA6186 order multiple times independently. Both Azosocius genomes contain one copy that was acquired by the UBA6186 order from a Chlamydiota organism and vertically inherited by Azosocius, but likely lost in Azoamicus. The Azosocius genomes additionally contain a second copy likely directly acquired from an alphaproteobacterium in the Rickettsiales order. The three Azoamicus genomes contain a single distinct tlcA copy, that also likely originated from a Rickettsiales bacterium7 and is shared with one of the UBA6186 genomes, hinting at the possibility of vertical inheritance and loss in Azosocius (Fig. 4b).
Genetic potential for carbon metabolism and cofactor biosynthesis in Azoamicaceae
While the genes encoding the key enzymes of the respiratory pathway are highly conserved across the Azoamicaceae, there is less consistency regarding the distribution of genes involved in carbon metabolism across the four genomes (Fig. 2d). Three of the four genomes encode genes for both the oxidation of malate to pyruvate (maeA) as well as to oxaloacetate (mdh). The Ca. A. ciliaticola genome only encodes mdh, which was highly transcribed in situ, leading to the suggestion that malate, which is likely provided by the host, acts as the primary electron donor for denitrification7. The role of malate as an important metabolite is further supported by the Ca. A. aquiferis transcription data analyzed here, showing that mdh is the third highest transcribed gene on average, across seven datasets. Conversely, no transcription of maeA could be detected in 6 out of 7 datasets (Fig. 3, Supplementary Data 7). Surprisingly, the smallest of the five genomes (Ca. A. soli) does not encode either of these enzymes for malate oxidation (Fig. 2d), although it does retain the anion permease proposed to import malate in Ca. A. ciliaticola (yflS, Supplementary Data 2)7. From the available data, it can not be excluded that Ca. A. soli oxidizes malate using host-encoded malate dehydrogenase, but it is also possible that succinate is oxidized by complex II and used as an electron donor instead (Fig. 2d).
Beyond the potential for malate and succinate oxidation, three of the genomes also encode 2-oxoacid:ferredoxin reductase (korAB) and succinyl-CoA synthetase (sucCD) that can oxidize 2-oxoglutarate to succinyl-CoA, and ultimately to succinate for further oxidation to fumarate using complex II. The sucCD genes are lacking in Ca. A. soli as well as in Ca. A aquiferis, the latter of which also appears to have lost the korAB genes (Fig. 2d, Fig. 5), suggesting use of only malate and (possibly) succinate as electron donors, or replacement of korAB and/or sucCD by host proteins.

Circular genome maps of Azoamicus (a) and Azosocius (b) genomes, with rings (inside to outside) indicating GC skew, GC content, reverse strand genes, forward strand genes. rRNA and tRNA genes are indicated in black, protein coding genes encoded in multiple genomes of a genus are indicated in gray. Protein coding genes differentially lost in Azoamicus are indicated in pink, and differentially lost in Azosocius in orange. The cytochrome cbb3 oxidase and associated genes are highlighted in blue, and the ATP/ADP transporter tlcA genes in purple.
In addition to genes involved in respiration, other genes conserved in all 5 genomes include those involved in the biosynthesis of metal cofactors. These include the sufBCDSU genes for iron-sulfur cluster biosynthesis, as well as the moaABCDE, moeAB, and mobA genes for the assembly of the molybdopterin cofactor required for nitrate reductase (Fig. 2d). In contrast, the modABC genes encoding a transporter for molybdenum are lost from the two Azosocius genomes. Interestingly, a near complete gene set for heme biosynthesis (hemABCDEJLN and glnS) remains in only the Ca. A. aquiferis genome, although hemH is missing. On the other hand, the cytochrome maturation system encoded by ccmABCEFH was found only in Ca. A. viridis. Notably, Ca. A. viridis also retained the capacity for fatty acid biosynthesis from pyruvate. Malonyl-CoA required for fatty acid biosynthesis can be formed from malate by the activities of malic enzyme (maeA), pyruvate dehydrogenase (aceEF, lpdA), and acetyl-CoA carboxylase (accABCD) (Fig. 5). The latter two complexes are only retained in Ca. A. viridis. Fatty acids can then be synthesized from malonyl-CoA by the enzymes of the type II fatty acid biosynthesis pathway (fabADFGHV) (Fig. 5)27. However, the genes encoding the membrane proteins required to convert fatty acids to phosphatidic acid are missing from Ca. A. viridis. Apart from two genes (cdsA, pgsA) no other genes for synthesis of phosphatidylglycerol or phosphatidylethanolamine were found. Instead, the genome encodes the same mla pathway thought to be used for lipid import in the other Azoamicaceae7 (Supplementary Data 2). Ca. A. viridis additionally encodes the inner membrane component of the LPS export system (lptBGF), as well as several of the steps needed to synthesize lipid-IVA (lpxABCDK). However, the genome is lacking lpxH, a gene considered essential for biosynthesis of the Lipid A moiety, as well as key genes of the heptose biosynthesis pathway and the outer membrane component of the export system (lptADE), and it seems unlikely that Ca. A. viridis can synthesize LPS for a typical outer cell membrane. It is peculiar that the Lipid A biosynthesis pathway has been retained in the strongly reduced genome of Ca. A. viridis, suggesting that it may have a yet unidentified role in the symbiosis.
Plagiopylean ciliates as putative hosts of the Azoamicaceae
To identify the potential hosts of the groundwater Azoamicaceae we searched for 18S rRNA gene sequences of Plagiopylean ciliates, the known host of Ca. A. ciliaticola, in samples from which genomes of the groundwater symbionts were recovered. The source metagenomic data for both Azosocius genomes contained two closely related full-length 18S rRNA gene sequences of Plagiopylean ciliates belonging to the same order (Odontostomatida) as the host of Ca. A. ciliaticola (Supplementary Fig. 7a). The combined source datasets for Ca. A. viridis and Ca. A. soli did not yield a full length Plagiopylean 18S rRNA gene sequence, but targeted assembly of reads mapping to CONThreeP 18S rRNA genes resulted in recovery of a fragment representing a partial Plagiopylean ciliate 18S rRNA gene sequence (Supplementary Fig. 7). As the branch containing the putative Azosocius hosts in the de novo calculated 18S rRNA gene tree (Supplementary Fig. 7a) has low support, we additionally placed the assembled putative host 18S rRNA gene sequences into a precalculated tree containing only the reference sequences. This approach gave the same topology, suggesting that the added sequences did not affect the branching pattern of the de novo calculated 18S rRNA gene tree (Supplementary Fig. 7b).
Presence and distribution of Azoamicaceae in global amplicon datasets
In addition to comparative genomics, the newly recovered genomes also provide an opportunity to assess the distribution of Azoamicaceae respiratory endosymbionts in diverse ecosystems across the world. For this, we searched public 16S rRNA gene amplicon datasets for the presence of the five endosymbiont 16S rRNA gene sequences. We identified 998 unique datasets containing putative Azoamicaceae sequences ( ≥ 3 reads at ≥ 95 % sequence identity). These datasets encompass samples from all continents, suggesting a global distribution of Azoamicaceae endosymbionts (Fig. 6). Such global distribution is congruent with the previously estimated early origin of this symbiosis ( ~ 106–213 Mya7) based on predictions of dispersal of microorganisms28,29.

Detection of the Azoamicaceae 16S rRNA gene sequences in publicly available 16S rRNA gene amplicon sequencing datasets shows a global distribution across all continents. Points represent amplicon datasets, and are colored according to their affiliation with the Azoamicaceae genome that have > 95% 16S rRNA gene identity to reads in the dataset. The hotspots in the global north likely reflect sampling bias.
It is interesting to note, that despite all four of our new genomes being from groundwater, only 18 samples (2%) containing Azoamicaceae reads were annotated as ‘groundwater metagenomes’ in our global survey. However, the quality of sample type annotation of 16S rRNA gene amplicon datasets is highly variable, and samples of other categories (such as “freshwater metagenome” or “soil metagenome”) might encompass more groundwater samples. In any case, the two most common sample types in which putative Azoamicaceae sequences were found were aquatic/freshwater and wastewater/activated sludge (Supplementary Data 8). The previously described Ca. A. cilaticola genome was indeed recovered from anoxic lake water, and our analyzes suggest that symbiont sequences most closely related ( ≥ 95 % identity) to Ca. A. ciliaticola are by far the most common and present in 808 of the 998 datasets (81 %). Relatives of Ca. A. agrarius were second most abundant and found in 143 datasets (14 %; Supplementary Data 8). The Azoamicaceae sequences were generally rare within the examined datasets, with Azoamicaceae reads accounting for ≦ 0.1 % of reads in 90 % of the retrieved datasets. This is consistent with the presence of endosymbionts constrained by host abundance, and may partly explain why the Azoamicaceae have only recently been discovered. Reads related to the Azoamicaceae accounted for ≥ 1 % of the reads in only 10 out of 998 datasets, which represent wastewater/activated sludge (6), freshwater lake (2), human gut (1), and groundwater (1). In general, our results indicate that wastewater is a widespread and probably important habitat for the symbionts that warrants future attention. Even though the five currently known genomes do not represent the full diversity of the Azoamicaceae, they already suggest a ubiquitous presence of the Azoamicaceae symbionts in diverse ecosystems around the world.
Discussion
The four Azoamicaceae genomes described in this study greatly expand our understanding of the diversity and distribution of respiratory endosymbiosis. We show that Azoamicaceae are globally distributed, and have the genomic potential for both denitrification and aerobic respiration. Of the metabolic capacities encoded in the new endosymbiont genomes it is undoubtedly the capacity to respire oxygen that has the most profound physiological and ecological implications. Retention of the cbb3 terminal oxidase in these extremely reduced genomes, as well as the comparatively high ccoNOP transcription in Ca. A. aquiferis, strongly implies an active role in endosymbiont and host physiology, by potentially conveying the capacity to respire oxygen to its host. Furthermore, the presence of the cbb3 oxidase can explain the broad distribution of the symbiont in seasonally or permanently oxic environments, thus considerably expanding the ecological role for this symbiosis. In fact, the capacity for aerobic respiration (or oxygen detoxification) may have facilitated the dispersal of the (presumably anaerobic) host across oxic environments. Endosymbionts are common in anaerobic ciliates, and syntrophic endosymbionts have been proposed to play a role in the transition of ciliates to anaerobiosis30,31. Given their potential for aerobic respiration, the Azoamicaceae may represent an interesting example of a ciliate endosymbiont facilitating a secondary adaptation, this time to a (micro)aerobic lifestyle.
We propose that Plagiopylean ciliates are the putative hosts of the groundwater Azoamicaceae. Indeed, based on a molecular analysis, Ciliophora were abundant in the wells of the Hainich CZE32, from which Ca. A. aquiferis was retrieved. We could reconstruct full length Plagiopylea 18S rRNA gene sequences from the metagenomes containing Ca. A. aquiferis as well as Ca. A. agrarius. A shorter Plagiopylea 18S rRNA gene fragment from the metagenome containing both Ca. A. viridis and Ca. A. soli could also be recovered. Interestingly, the phylogeny of 18S rRNA sequences (Supplementary Fig. 7) is congruent with the genome phylogeny of their respective putative symbionts (Fig. 1) and thus could be indicative of long-term vertical inheritance of the symbionts in the Odontostomatida order. This is further supported by the extremely reduced nature of the symbiont genomes that is consistent with an older, likely vertically inherited, symbiosis6. However, more data on the host identity are needed to confirm an exclusively vertical inheritance of the Azoamicaceae symbionts, as some protist hosts are able to replace their symbionts33,34. Additionally, while an apparent vertical inheritance of protist endosymbionts can be observed over short evolutionary time35, examples of long term vertical inheritance are more rare, possibly due to eventual replacement of protist symbionts6, or simply undersampling36. Our comparative genomics of the five Azoamicaceae genomes shows a varying degree of genome erosion, as observed in insect HBEs37,38. The Azoamicaceae seem to converge on a minimal gene set encoding the genes required for ATP generation using a denitrifying and oxygen respiring respiratory chain, with the conserved core genes comprising over 80% of the protein complement of the smallest genomes. The loss of biosynthetic pathways essential for the respiratory function, such as heme biosynthesis and cytochrome c maturation, strongly suggests that host proteins are targeted to the endosymbiont. Host protein targeting has previously been observed in rare cases in insect HBEs39,40 in the trypanosomatid Angomonas deanei41, and more extensively in the chromatophore of Paulinella chromatophora42 and in the nitroplast (formerly Candidatus Atelocyanobacterium thalassa) of Braarudosphaera bigelowii43. These observations have blurred the boundaries between HBEs and organelles36,44, and the extant Azoamicaceae may represent another snapshot of the transition between the two. While comparative genomic analyzes can provide a great insight into the evolutionary history and metabolic potential of these enigmatic endosymbionts, the establishment of an experimentally tractable laboratory culture of the host is essential to test these exciting hypotheses.
Method
Database searching and cMAG curation
The 16S rRNA sequence and nitrate reductase sequence of Ca. A. ciliaticola were used to search public genome databases. This led to the identification of the Candidatus Azosocius agrarius cMAG consisting of a single circular contig with 100 bp overlap (CP066692), binned from a groundwater sample taken in Modesto, CA, USA (37.661915 N, 121.114554 W) in 2018 (SAMN15459604)13. For this study, we removed the overlap and started the contig at the putative origin of replication, resulting in a contig of 352,496 bp. The fasta file with the modified cMAG is provided as Supplementary Data 1.
To identify further samples containing respiratory endosymbionts in groundwater, we used the 16S rRNA gene of Ca. A. agrarius to query publicly available 16S rRNA gene amplicon sequencing data sets using the web search tool IMNGS45. This search led to matches with > 95% sequence identity in amplicon datasets from a groundwater aquifer located near Hainich national park in Germany, through the long term AquaDiva project (Supplementary Data 8). We screened the assembled metagenomes from the AquaDiva project (51.119338 N, 10.469198 W; Bioproject PRJEB36523)14 for contigs matching the Ca. A. agrarius cMAG using BLASTn46 (version 2.6.0) with a minimum length cutoff of 3000 bp. We iteratively mapped the trimmed reads from datasets ERR3858113, ERR3858114, ERR3858115 on the resulting 13 contigs using coverM (version 0.7.0) (https://github.com/wwood/CoverM), followed by reassembly of the mapping reads using Spades47 (version 3.15) with the –isolate flag, resulting in a circular contig of 353051 bp after three iterations, as confirmed by visualization in Bandage48 (version 0.9.0). This circular contig represents the cMAG of Candidatus Azosocius aquiferis and is deposited to genbank under accession number NCBI BioSample SAMN39831648 and NCBI GenBank accession GCA_043390185.1.
For further endosymbiont discovery we constructed a database of proteins encoded by tlcA gene, encoding the NTT transporter required for the exchange of ATP and ADP with the host, and screened sequencing runs using a blast score ratio (BSR) approach as previously described49,50. We then queried the SRA for groundwater datasets, and selected large and medium scale BioProjects to screen. We screened a combined 256 sequencing runs from BioProjects PRJNA640378, PRJEB36505, PRJEB28738, PRJNA530103, PRJNA268031, PRJNA292723, PRJEB32173, PRJEB14718, PRJNA513876, and PRJNA512237 encompassing groundwater samples from Australia, Saudi Arabia, Germany, and USA (Colorado, Tennessee, California and Ohio). While we found evidence for the presence of putative endosymbionts (at low abundance) in several datasets, we only managed to reconstruct additional cMAGs, which was the quality threshold we set for this study, from sequencing runs from bioproject PRJNA51223716. cMAG reconstruction for Ca. A. viridis and Ca. A. soli was performed by first co-assembling trimmed reads from datasets SRR8863434, SRR8863435, and SRR8863439 using MEGAHIT51 (version 1.2.9). The resulting contigs were searched using BLASTn46(version 2.6.0) using the three other cMAGs as query, keeping hits with a minimum length cutoff of 3000 bp. This approach yielded 3 contigs, a circular contig representing the cMAG of Candidatus Azoamicus soli, and two contigs forming the MAG of Candidatus Azoamicus viridis. The Ca. A. viridis MAG was then circularized as described above for Candidatus A. aquiferis. The cMAG of Candidatus Azoamicus soli is available via NCBI BioSample SAMN39831885 and NCBI GenBank accession GCA_043390205.1 and the cMAG of Candidatus Azoamicus viridis is available via NCBI BioSample SAMN39831884 and NCBI GenBank accession GCA_043390165.1.
Genome annotation and comparative genomics
cMAGs were analyzed using anvi’o52 (version 7.1), with gene calling by prodigal53 (version 2.6.3). Annotation of protein coding genes was transferred from Ca. A. ciliaticola where appropriate based on bidirectional best hit DIAMOND (version 2.0.14.152)54, and the genes were annotated using KEGG55, COG56 and PFAM57 annotation as integrated in anvi’o, followed by manual curation of the annotation. The final annotation is provided as Supplementary Data 2. The UpSet plot58 of the cMAG gene content was generated from a presence absence matrix of the genes content using the UpSetR R package (version 1.4.0)59.
The average nucleotide identity between the cMAGs was calculated using fastANI17 (version 1.33) with a fragment length of 1000 basepairs. Average amino acid identity was calculated using ezAAI60 (version 1.2.2) with default settings. 16S rRNA genes were extracted from the cMAGs using anvi’o (development version)52, and pairwise identity was calculated using BLASTn (version 2.6.0)46.
Metabolic prediction of the cMAGs was done using the anvi-estimate-metabolism program (https://anvio.org/help/main/programs/anvi-estimate-metabolism/) as integrated in anvi’o52 (development version) using KEGG modules annotation.
Pseudogene and single nucleotide variant detection
Pseudogenes in the Azoamicaceae genomes were detected using pseudofinder (version 1.1.0)61. The Azoamicaceae genomes were reannotated using prokka (version 1.14.6)62 due to required genbank file format input. Predicted proteins and intergenic regions were used as BLASTp and BLASTx (version 2.12.0 + )46 queries against the NCBI-nr protein database (downloaded 11 June 2024). This resulted in 13–16 predicted pseudogenes per genome. The gene coordinates of predicted pseudogenes were compared with the anvi’o annotation, and pseudogenes consistent across both annotation methods (3–8 per genome) were added to Supplementary Data 5. Single nucleotide variation in the five Azoamicaceae genomes was assessed using inStrain (version 1.9.0)63 after read mapping with coverM (version 0.7.0) (https://github.com/wwood/CoverM) requiring mapped reads to have > 95 % identity over > 80 % of the read length. Results for analyzed dataset and reference pairs are summarized in Supplementary Data 6.
Metatranscriptome analysis
The 18 metatranscriptome sequencing read datasets in BioProject PRJEB28738 were retrieved from NCBI and trimmed using the cutadapt (version 3.7)64 wrapper trim-galore (version 0.6.10) (https://github.com/FelixKrueger/TrimGalore) with default settings and automatic adapter detection. The trimmed reads were mapped against all gene sequences in the Ca. A. aquiferis genome using coverM (version 0.7.0) (https://github.com/wwood/CoverM), with minimum identity 95 % and minimum length fraction 80 %, exporting the read counts per gene sequence. Based on total reads mapped, seven datasets were selected for analysis (Supplementary Data 7). Read counts matching protein coding genes were used for the calculation of transcript per million (TPM) values65 and genes were ranked according to average TPM across the seven datasets. For visualization of the transcriptome in a heatmap (Fig. 3), transcript abundance was recalculated as “transcript per thousand”, following the same procedure as for TPM but using a 103 scaling factor instead of 106 to account for the number of reads mapped to Ca. A. aquiferis. Datasets included in the heatmap are ERR2809165 (H52_1), ERR2809163 (H52_2), ERR2809153 (H52_3), ERR2809156 (H41_1), ERR2809155 (H41_2), ERR2809154 (H41_3), and ERR2809148 (H41_4). Samples H5-2_1, H5-2_2, and H5-2_3 represent replicated metatranscriptome data from the same well (H5-2) from sampling campaign PNK69. Sample H4-1_4 represents a metatranscriptomic data set from three pooled RNA preparations from well H4-1 from from sampling campaign PNK66.
Global detection of Azoamicaceae
The 16S rRNA genes from the four cMAGs described here, and the previously published Candidatus Azoamicus ciliaticola, were used to screen 16S rRNA gene amplicon datasets for matches using the IMNGS web search tool45. 998 Datasets with more than 3 reads matching any of the five cMAGs at ≥ 95 % identity were retained for further analysis. The dataset IDs were used to retrieve sample metadata from NCBI, and sample coordinates were manually standardized. Samples with coordinates were plotted on world map (Fig. 6) in R using packages maps (https://CRAN.R-project.org/package=maps) and mapdata (https://CRAN.R-project.org/package=mapdata). For 210 of the 998 datasets no coordinates were provided and these are therefore not included in Fig. 6. To assess environmental distribution of the Azoamicaceae, the 998 datasets were grouped by their dataset description (defined upon dataset submission to INSDC databases).
Putative host detection
To gain insight into potential host organisms for the organisms represented by the cMAGs, we attempted to reconstruct full length 18S rRNA gene sequences from their source datasets using Phyloflash (version 3.4.2)66. One 18S rRNA gene sequence was from each of ERR3858113, ERR3858114, ERR3858115 (source data for Ca. A. aquiferis). These three sequences were identical, and belonged to a ciliate of the Plagiopylea class. Four 18S rRNA gene sequences were retrieved from SRR12113421 (source data for Ca. A. agrarius). One of the four sequences obtained from SRR12113421 belonged to a ciliate of the Plagiopylea class, closely related to the sequences obtained from ERR3858113, ERR3858114, ERR3858115. No full length 18S rRNA gene sequences were retrieved from datasets SRR8863434, SRR8863435, and SRR8863439 (source data for Ca. A. viridis and Ca. A. soli), but reads matching CONthreeP 18S sequences were detectable using PhyloFlash. Fasta sequences of the reads matching CONthreeP were extracted from the PhyloFlash output of SRR8863434, SRR8863435, and SRR8863439, combined, and assembled using idba-ud (version 1.1.3)67, resulting in one contig representing a partial Plagiopylean 18S rRNA gene sequence.
18S rRNA gene phylogeny
The four recovered (partial) putative host 18S rRNA gene sequences were used as input for the web based Silva alignment, classification, and tree service (Silva-ACT https://www.arb-silva.de/aligner/). The maximum number of neighbor sequences was set to 100 (maximum) and the identity was set to 75%. Alignment was done with the SINA aligner (version 1.2.12)68 with the variability profile for Eukaryota. A de novo tree with neighbor sequences was computed using Fasttree69 (no version provided in Silva-ACT web interface) with the GTR model and Gamma likelihood rate model with the stringent positional variability filter for Eukarya. The resulting tree was visualized in iToL70. To support the obtained topology, a second tree was generated using the Silva-ACT web interface, by placing the four putative host sequences in the tree of neighbors using RaxML71 (no version provided in Silva-ACT web interface) and the resulting tree was also visualized in iToL.
Phylogenetic analyzes of individual and concatenated amino acid sequences
Phylogenetic analysis of protein coding genes (nuoG, sdhA, qcrB, atpA, ccoN, narG, nirK, norB, nosZ, tlcA) was done using protein sequences obtained from genomes in the genome taxonomy database (GTDB, version 207)72 and the genomic catalog of Earth’s microbiomes73. Sequences were aligned using MUSCLE74 (version 3.8.31), and phylogenetic trees were calculated using IQ-tree75 (version 2.2.0) with automatic model selection using ModelFinder76 (integrated in IQ-tree, no separate version provided) and ultrafast bootstrapping (1000 replicates) with UFBoot277 (integrated in IQ-tree, no separate version provided). Models selected by ModelFinder were: NuoG - Q.pfam+R10; SdhA - LG + I + I + R10; QcrB - LG + F + I + I + R10; AtpA - Q.yeast+I + I + R10; CcoN - Q.pfam+I + I + R10; NarG - Q.yeast+I + I + R7; NirK - Q.pfam+I + I + R9; NorB - Q.yeast+F + I + I + R6; NosZ - LG + F + I + I + R7; TlcA - Q.yeast+F + R10).
Concatenated marker gene phylogenies were calculated based on protein sequences identified using hmmer78 (version 3.3.2) with the Bacteria_71 HMM set included in Anvi’o52 (version 7.1). The anvi-get-sequences-for-hmm-hits program was used to retrieve sequences for HMM hits, align them using MUSCLE74 (version 3.8.31), concatenate alignments, and write a partition file. The concatenated alignments and corresponding partition files were used to calculate phylogenies using IQ-tree75 (version 2.2.0) with automatic model selection using ModelFinder76 (integrated in IQ-tree, no separate version provided) and ultrafast bootstrapping (1000 replicates) with UFBoot277 (integrated in IQ-tree, no separate version provided). All data for the phylogenetic analyzes are available on figshare at https://figshare.com/projects/Groundwater_Azoamicaceae_phylogenies/205756.
Etymology
Description of ‘Candidatus Azoamicus viridis’ sp. nov
‘Candidatus Azoamicus viridis’ (L. masc. adj. viridis, green pertaining to Greene county, where the sample containing the species was taken). A bacterial species identified by metagenomic analyzes. This species includes all bacteria with genomes that show ≥ 95% average nucleotide identity to the type genome for the species to which is available via NCBI BioSample SAMN39831884 and NCBI GenBank accession GCA_043390165.1.
Description of ‘Candidatus Azoamicus soli’ sp. nov
‘Candidatus Azoamicus soli’ (L. masc. n. soli, of the earth). A bacterial species identified by metagenomic analyzes. This species includes all bacteria with genomes that show ≥ 95% average nucleotide identity to the type genome for the species to which is available via NCBI BioSample SAMN39831885 and NCBI GenBank accession GCA_043390205.1.
Description of ‘Candidatus Azosocius’ gen. nov
‘Candidatus Azosocius’ (N.L. masc. n. Azosocius, combines the prefix azo- (N. L., pertaining to nitrogen) with socius (Latin, masc. n., associate); thus giving azosocius (‘associate that pertains to nitrogen’). A bacterial genus identified by metagenomic analyzes and delineated according to Relative Evolutionary Distance by the Genome Taxonomy Database (GTDB). The type species of the genus is ‘Candidatus Azosocius agrarius’.
Description of ‘Candidatus Azosocius agrarius’ sp. nov
‘Candidatus Azosocius agrarius’ (L. masc. adj. agrarius, of the soil). A bacterial species identified by metagenomic analyzes. This species includes all bacteria with genomes that show ≥ 95% average nucleotide identity to the type genome for the species to which is available via NCBI BioSample SAMN15435421 and NCBI GenBank accession GCA_016432505.1.
Description of ‘Candidatus Azosocius aquiferis’ sp. nov
‘Candidatus Azosocius aquiferis’ (N.L. gen. masc. n. aquiferis, of an aquifer). A bacterial species identified by metagenomic analyzes. This species includes all bacteria with genomes that show ≥ 95% average nucleotide identity to the type genome for the species to which is available via NCBI BioSample SAMN39831648 and NCBI GenBank accession GCA_043390185.1.
Description of ‘Candidatus Azoamicaceae’ fam. nov
‘Candidatus Azoamicaceae’ (A.zo.a.mi.ca.ce.ae. N.L. masc. n. Azoamicus. type genus of the family; N.L. suff. –aceae to denote a family; N.L. fem. pl. n. Azoamicaceae, the family of the genus Azoamicus). The description of the family ‘Candidatus Azoamicaceae’ is the same as that of the genus ‘Candidatus Azoamicus’. The type genus is ‘Candidatus Azoamicus’.
Description of ‘Candidatus Azoamicales’ ord. nov
‘Candidatus Azoamicales’ (A.zo.a.mi.ca.les. N.L. masc. n. Azoamicus. type genus of the order; N.L. suff. –ales to denote an order; N.L. fem. pl. n. Azoamicales, the order of the genus Azoamicus). A bacterial order identified by metagenomic analyzes and delineated according to Relative Evolutionary Distance by the Genome Taxonomy Database (GTDB). Phylogenetic analyzes in this work have shown that the previously published ‘Candidatus Azoamicus ciliaticola’ is assigned to an uncharacterized order with provisional designation UBA6186. Consequently, we propose to rename the UBA6186 order to ‘Candidatus Azoamicales’. the type genus of the order is ‘Candidatus Azoamicus’. The order is assigned to the class Gammaproteobacteria.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The sequencing data used to construct the symbiont genomes described in this study is publicly available in Genbank under BioProject numbers PRJEB36523 and PRJNA512237. The successfully reconstructed genomes are available in Genbank under Bioproject number PRJNA1073475 and accession numbers GCA_043390165.1, GCA_043390205.1, and GCA_043390185.1. The previously reconstructed genome that we identified as Candidatus Azosocius agrarius is available in Genbank under accession number GCA_016432505.1. Metatranscriptome sequencing data used was publicly available in Genbank under BioProject accession number PRJEB28738. 16S rRNA gene amplicon data was accessed through the IMNGS web server (https://www.imngs.org/). Data files for phylogenetic trees discussed in the manuscript, as well as the anvi’o generated genome annotation files used to generate Supplementary Data 2 are available on figshare at https://doi.org/10.6084/m9.figshare.c.7510170.v1. Source data for all other figures can be found in Supplementary Data files 1-8.
References
McCutcheon, J. P., Boyd, B. M. & Dale, C. The life of an insect endosymbiont from the cradle to the grave. Curr. Biol. 29, R485–R495 (2019).
Douglas, A. E. et al. Nutritional interactions in insect-microbial symbioses: aphids and their symbiotic bacteria Buchnera. https://doi.org/10.1146/annurev.ento.43.1.17 (2003).
Moran, N. A., Plague, G. R., Sandström, J. P. & Wilcox, J. L. A genomic perspective on nutrient provisioning by bacterial symbionts of insects. Proc. Natl Acad. Sci. USA 100, 14543–14548 (2003).
Brownlie, J. C. & Johnson, K. N. Symbiont-mediated protection in insect hosts. Trends Microbiol 17, 348–354 (2009).
Van Arnam, E. B., Currie, C. R. & Clardy, J. Defense contracts: molecular protection in insect-microbe symbioses. Chem. Soc. Rev. 47, 1638–1651 (2018).
Husnik, F. et al. Bacterial and archaeal symbioses with protists. Curr. Biol. 31, R862–R877 (2021).
Graf, J. S. et al. Anaerobic endosymbiont generates energy for ciliate host by denitrification. Nature 591, 445–450 (2021).
Müller, M. et al. Biochemistry and evolution of anaerobic energy metabolism in eukaryotes. Microbiol. Mol. Biol. Rev. 76, 444–495 (2012).
Stairs, C. W., Leger, M. M. & Roger, A. J. Diversity and origins of anaerobic metabolism in mitochondria and related organelles. Philos. Trans. R. Soc. Lond. B Biol. Sci. 370, 20140326 (2015).
Embley, T. M. et al. Multiple origins of anaerobic ciliates with hydrogenosomes within the radiation of aerobic ciliates. Proc. R. Soc. Lond. Ser. B: Biol. Sci. 262, 87–93 (1997).
Chen, J. & Strous, M. Denitrification and aerobic respiration, hybrid electron transport chains and co-evolution. Biochim. Biophys. Acta 1827, 136–144 (2013).
Kuypers, M. M. M., Marchant, H. K. & Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 16, 263–276 (2018).
He, C. et al. Genome-resolved metagenomics reveals site-specific diversity of episymbiotic CPR bacteria and DPANN archaea in groundwater ecosystems. Nat. Microbiol 6, 354–365 (2021).
Overholt, W. A. et al. Carbon fixation rates in groundwater similar to those in oligotrophic marine systems. Nat. Geosci. 15, 561–567 (2022).
Küsel, K. et al. How deep can surface signals be traced in the critical zone? merging biodiversity with biogeochemistry research in a central German muschelkalk landscape. Front. Earth Sci. 4, 32 (2016).
Danczak, R. E., Johnston, M. D., Kenah, C., Slattery, M. & Wilkins, M. J. Capability for arsenic mobilization in groundwater is distributed across broad phylogenetic lineages. PLoS One 14, e0221694 (2019).
Jain, C., Rodriguez-R, L. M., Phillippy, A. M., Konstantinidis, K. T. & Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 9, 5114 (2018).
Barco, R. A. et al. A genus definition for bacteria and archaea based on a standard genome relatedness index. MBio 11, 02475-19 (2020).
Pitcher, R. S. & Watmough, N. J. The bacterial cytochrome cbb3 oxidases. Biochim. Biophys. Acta 1655, 388–399 (2004).
Kohlstaedt, M., Buschmann, S., Langer, J. D., Xie, H. & Michel, H. Subunit CcoQ is involved in the assembly of the Cbb3-type cytochrome c oxidases from Pseudomonas stutzeri ZoBell but not required for their activity. Biochim. Biophys. Acta Bioenerg. 1858, 231–238 (2017).
Ducluzeau, A.-L., Ouchane, S. & Nitschke, W. The cbb3 oxidases are an ancient innovation of the ___domain bacteria. Mol. Biol. Evol. 25, 1158–1166 (2008).
Chang, H.-Y. et al. The diheme cytochrome c(4) from Vibrio cholerae is a natural electron donor to the respiratory cbb(3) oxygen reductase. Biochemistry 49, 7494–7503 (2010).
Deeudom, M., Koomey, M. & Moir, J. W. B. Roles of c-type cytochromes in respiration in Neisseria meningitidis. Microbiology 154, 2857–2864 (2008).
Wegner, C.-E. et al. Biogeochemical regimes in shallow aquifers reflect the metabolic coupling of the elements nitrogen, sulfur, and carbon. Appl. Environ. Microbiol. 85, e02346-18 (2019).
Kohlhepp, B. et al. Aquifer configuration and geostructural links control the groundwater quality in thin-bedded carbonate–siliciclastic alternations of the Hainich CZE, central Germany. Hydrol. Earth Syst. Sci. 21, 6091–6116 (2017).
Kawakami, T., Kuroki, M., Ishii, M., Igarashi, Y. & Arai, H. Differential expression of multiple terminal oxidases for aerobic respiration in Pseudomonas aeruginosa. Environ. Microbiol. 12, 1399–1412 (2010).
Zhang, Y.-M. & Rock, C. O. Membrane lipid homeostasis in bacteria. Nat. Rev. Microbiol. 6, 222–233 (2008).
Martiny, J. B. H. et al. Microbial biogeography: putting microorganisms on the map. Nat. Rev. Microbiol. 4, 102–112 (2006).
Nemergut Diana, R. et al. Patterns and processes of microbial community assembly. Microbiol. Mol. Biol. Rev. 77, 342–356 (2013).
Rotterová, J., Edgcomb, V. P., Čepička, I. & Beinart, R. Anaerobic ciliates as a model group for studying symbioses in oxygen-depleted environments. J. Eukaryot. Microbiol. 69, e12912 (2022).
Rotterová, J. et al. Genomics of new ciliate lineages provides insight into the evolution of obligate anaerobiosis. Curr. Biol. 30, 2037–2050.e6 (2020).
Herrmann, M. et al. Complex food webs coincide with high genetic potential for chemolithoautotrophy in fractured bedrock groundwater. Water Res 170, 115306 (2020).
Boscaro, V., Husnik, F., Vannini, C. & Keeling, P. J. Symbionts of the ciliate Euplotes: diversity, patterns and potential as models for bacteria-eukaryote endosymbioses. Proc. Biol. Sci. 286, 20190693 (2019).
Boscaro, V. et al. All essential endosymbionts of the ciliate Euplotes are cyclically replaced. Curr. Biol. 32, R826–R827 (2022).
Lind, A. E. et al. Genomes of two archaeal endosymbionts show convergent adaptations to an intracellular lifestyle. ISME J. 12, 2655–2667 (2018).
Husnik, F. & Keeling, P. J. The fate of obligate endosymbionts: reduction, integration, or extinction. Curr. Opin. Genet. Dev. 58-59, 1–8 (2019).
Moran, N. A., McLaughlin, H. J. & Sorek, R. The dynamics and time scale of ongoing genomic erosion in symbiotic bacteria. Science 323, 379–382 (2009).
McCutcheon, J. P. & Moran, N. A. Extreme genome reduction in symbiotic bacteria. Nat. Rev. Microbiol. 10, 13–26 (2011).
Nakabachi, A., Ishida, K., Hongoh, Y., Ohkuma, M. & Miyagishima, S.-Y. Aphid gene of bacterial origin encodes a protein transported to an obligate endosymbiont. Curr. Biol. 24, R640–R641 (2014).
Bublitz, D. C. et al. Peptidoglycan production by an insect-bacterial mosaic. Cell 179, 703–712.e7 (2019).
Morales, J. et al. Host-symbiont interactions in Angomonas deanei include the evolution of a host-derived dynamin ring around the endosymbiont division site. Curr. Biol. 33, 28–40.e7 (2023).
Nowack, E. C. M. & Grossman, A. R. Trafficking of protein into the recently established photosynthetic organelles of Paulinella chromatophora. Proc. Natl Acad. Sci. USA. 109, 5340–5345 (2012).
Coale, T. H. et al. Nitrogen-fixing organelle in a marine alga. Science 384, 217–222 (2024).
McCutcheon, J. P. The genomics and cell biology of host-beneficial intracellular infections. Annu. Rev. Cell Dev. Biol. 37, 115–142 (2021).
Lagkouvardos, I. et al. IMNGS: A comprehensive open resource of processed 16S rRNA microbial profiles for ecology and diversity studies. Sci. Rep. 6, 33721 (2016).
Altschul, S. F., Gish, W., Miller, W., Myers, E. W. & Lipman, D. J. Basic local alignment search tool. J. Mol. Biol. 215, 403–410 (1990).
Bankevich, A. et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 19, 455–477 (2012).
Wick, R. R., Schultz, M. B., Zobel, J. & Holt, K. E. Bandage: interactive visualization of de novo genome assemblies. Bioinformatics 31, 3350–3352 (2015).
Rasko, D. A., Myers, G. S. A. & Ravel, J. Visualization of comparative genomic analyses by BLAST score ratio. BMC Bioinforma. 6, 2 (2005).
Speth, D. R. & Orphan, V. J. Metabolic marker gene mining provides insight in global mcrA diversity and, coupled with targeted genome reconstruction, sheds further light on metabolic potential of the Methanomassiliicoccales. PeerJ 6, e5614 (2018).
Li, D., Liu, C.-M., Luo, R., Sadakane, K. & Lam, T.-W. MEGAHIT: an ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 31, 1674–1676 (2015).
Eren, A. M. et al. Community-led, integrated, reproducible multi-omics with anvi’o. Nat. Microbiol 6, 3–6 (2021).
Hyatt, D. et al. Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinforma. 11, 119 (2010).
Hernández-Salmerón, J. E. & Moreno-Hagelsieb, G. Progress in quickly finding orthologs as reciprocal best hits: comparing blast, last, diamond and MMseqs2. BMC Genomics 21, 741 (2020).
Kanehisa, M. & Goto, S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res 28, 27–30 (2000).
Galperin, M. Y., Makarova, K. S., Wolf, Y. I. & Koonin, E. V. Expanded microbial genome coverage and improved protein family annotation in the COG database. Nucleic Acids Res 43, D261–D269 (2015).
El-Gebali, S. et al. The Pfam protein families database in 2019. Nucleic Acids Res 47, D427–D432 (2019).
Lex, A., Gehlenborg, N., Strobelt, H., Vuillemot, R. & Pfister, H. UpSet: visualization of intersecting sets. IEEE Trans. Vis. Comput. Graph. 20, 1983–1992 (2014).
Conway, J. R., Lex, A. & Gehlenborg, N. UpSetR: an R package for the visualization of intersecting sets and their properties. Bioinformatics 33, 2938–2940 (2017).
Kim, D., Park, S. & Chun, J. Introducing EzAAI: a pipeline for high throughput calculations of prokaryotic average amino acid identity. J. Microbiol. 59, 476–480 (2021).
Syberg-Olsen, M. J., Garber, A. I., Keeling, P. J., McCutcheon, J. P. & Husnik, F. Pseudofinder: detection of pseudogenes in prokaryotic genomes. Mol. Biol. Evol. 39, msac153 (2022).
Seemann, T. Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069 (2014).
Olm, M. R. et al. inStrain profiles population microdiversity from metagenomic data and sensitively detects shared microbial strains. Nat. Biotechnol. 39, 727–736 (2021).
Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 17, 10–12 (2011).
Li, B., Ruotti, V., Stewart, R. M., Thomson, J. A. & Dewey, C. N. RNA-Seq gene expression estimation with read mapping uncertainty. Bioinformatics 26, 493–500 (2010).
Gruber-Vodicka, H. R., Seah, B. K. B. & Pruesse, E. phyloFlash: rapid small-subunit rRNA profiling and targeted assembly from metagenomes. mSystems 5, 00920-20 (2020).
Peng, Y., Leung, H. C. M., Yiu, S. M. & Chin, F. Y. L. IDBA-UD: a de novo assembler for single-cell and metagenomic sequencing data with highly uneven depth. Bioinformatics 28, 1420–1428 (2012).
Pruesse, E., Peplies, J. & Glöckner, F. O. SINA: accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 28, 1823–1829 (2012).
Price, M. N., Dehal, P. S. & Arkin, A. P. FastTree 2–approximately maximum-likelihood trees for large alignments. PLoS One 5, e9490 (2010).
Letunic, I. & Bork, P. Interactive Tree Of Life (iTOL) v4: recent updates and new developments. Nucleic Acids Res 47, W256–W259 (2019).
Stamatakis, A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313 (2014).
Parks, D. H. et al. A standardized bacterial taxonomy based on genome phylogeny substantially revises the tree of life. Nat. Biotechnol. 36, 996–1004 (2018).
Nayfach, S. et al. A genomic catalog of Earth’s microbiomes. Nat. Biotechnol. 39, 499–509 (2020).
Edgar, R. C. MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC Bioinforma. 5, 113 (2004).
Nguyen, L.-T., Schmidt, H. A., von Haeseler, A. & Minh, B. Q. IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 32, 268–274 (2015).
Kalyaanamoorthy, S., Minh, B. Q., Wong, T. K. F., von Haeseler, A. & Jermiin, L. S. ModelFinder: fast model selection for accurate phylogenetic estimates. Nat. Methods 14, 587–589 (2017).
Hoang, D. T., Chernomor, O., von Haeseler, A., Minh, B. Q. & Vinh, L. S. UFBoot2: improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 35, 518–522 (2018).
Eddy, S. R. Profile hidden Markov models. Bioinformatics 14, 755–763 (1998).
Acknowledgements
We thank Falko Gutmann and Robert Lehmann for assistance with sampling groundwaters at the Hainich CZE as part of the Collaborative Research Center AquaDiva. This study used publicly available data from Bioprojects PRJNA640378, PRJEB36523, and PRJNA512237 and we thank the authors of those studies for making their data available. This study was financially supported by the Max Planck Gesellschaft. This research was further supported by the Deutsche Forschungsgemeinschaft (DFG) under Germany’s Excellence Strategy—EXC 2051, Project-ID 390713860 (to K.K.) and the Collaborative Research Center AquaDiva (CRC 1076 AquaDiva - Project-ID 218627073, to K.K) of the Friedrich Schiller-University Jena.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
D.R.S. and J.M. designed research. D.R.S. and L.M.Z. organized and performed sampling at the Hainich critical zone exploratory (CZE) through the Collaborative Research Center (CRC) AquaDiva and analyzed field sampling data. J.S.G performed initial 16S rRNA gene surveys and identified the Ca. Azosocius agrarius genome. K.K. and W.A.O. contributed data from previous sampling campaigns by CRC AquaDiva at the Hainich CZE. D.R.S. processed, synthesized, and analyzed sequence data. D.R.S. and J.M. interpreted genomic data. D.R.S. and J.M. wrote the manuscript with contributions from all authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Speth, D.R., Zeller, L.M., Graf, J.S. et al. Genetic potential for aerobic respiration and denitrification in globally distributed respiratory endosymbionts. Nat Commun 15, 9682 (2024). https://doi.org/10.1038/s41467-024-54047-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-024-54047-x
