
Abstract
Ubiquitin-specific proteases (USPs) represent the largest class of human deubiquitinases (DUBs) and comprise its phylogenetically most distant members USP53 and USP54, which are annotated as catalytically inactive pseudoenzymes. Conspicuously, mutations within the USP ___domain of USP53 cause progressive familial intrahepatic cholestasis. Here, we report the discovery that USP53 and USP54 are active DUBs with high specificity for K63-linked polyubiquitin. We demonstrate how USP53 mutations abrogate catalytic activity, implicating loss of DUB activity in USP53-mediated pathology. Depletion of USP53 increases K63-linked ubiquitination of tricellular junction components. Assays with substrate-bound polyubiquitin reveal that USP54 cleaves within K63-linked chains, whereas USP53 can en bloc deubiquitinate substrate proteins in a K63-linkage-dependent manner. Biochemical and structural analyses uncover underlying K63-specific S2 ubiquitin-binding sites within their catalytic domains. Collectively, our work revises the annotation of USP53 and USP54, provides reagents and a mechanistic framework to investigate K63-linked polyubiquitin decoding and establishes K63-linkage-directed deubiquitination as a new DUB activity.

Similar content being viewed by others

Main
The covalent attachment of the small protein ubiquitin to substrates and to other ubiquitin moieties facilitates the formation of a diverse array of post-translational modifications; this attachment is characterized by structural and functional heterogeneity and collectively termed the ‘ubiquitin code’ (refs. 1,2). In addition to monoubiquitination, polyubiquitin chains of eight linkages exist, with K48-linked and K63-linked chains being the most abundant in human cells1,3. While the attachment of K48-linked ubiquitin chains to substrates commonly facilitates proteasomal degradation, predominantly nondegradative roles have been described for K63-linked chains, including in intracellular trafficking, innate immune signaling and genome maintenance4,5,6. Ubiquitin chain length encodes an additional layer of information1,2,7,8,9,10,11,12,13. However, in contrast to K11-linked and K48-linked polyubiquitin chains9,14,15,16,17, molecular mechanisms for length-dependent decoding of K63-linked chains have remained elusive.
Deubiquitinating enzymes (DUBs) edit ubiquitin chains or cleave ubiquitin–substrate linkages, thereby regulating the ubiquitination status of proteins to determine their stability, localization, activity and function1,18,19,20,21. The human genome encodes approximately 100 DUBs, which can be classified into ubiquitin linkage-specific and nonspecific enzymes11,15,22,23,24. High ubiquitin linkage specificity has been demonstrated for members of the ovarian tumor protease (OTU; diverse linkages)15, MINDY (K48)22, ZUFSP (K63)6,11,23,25 and JAMM (K63)26,27,28 families. In contrast, DUBs of the ubiquitin C-terminal hydrolase (UCH), Josephin and ubiquitin-specific protease (USP) families generally show poor discrimination of linkages24. Of those, USPs form the largest and most diverse DUB family with ~60 members in humans29.
USP53 and USP54 are poorly characterized USP family members30,31. They possess high homology within their predicted catalytic domains but show otherwise very low sequence homology to all other USP members20. Notably, both proteins have been reported to be catalytically inactive20,32 because of the absence of strongly conserved residues before their catalytic histidines32, the inability of bacterially expressed USP53 to hydrolyze a monoubiquitin substrate33 and the lack of reactivity of cellular USP53 with a ubiquitin probe30. However, we were intrigued by recent reports that identified loss or biallelic mutations in USP53 as the cause for progressive familial intrahepatic cholestasis, a hereditary liver disorder in children34,35,36,37. This phenotype and proposed cellular roles linked to USP53 and USP54 have not been reconciled with their annotation as pseudoenzymes31. In addition, the lack of an in vitro assessment of catalytic activity and unavailable structural information have complicated insights into pathology-related molecular mechanisms.
Here, we report USP53 and USP54 to be active DUBs with specific cleavage activities on K63-linked ubiquitin chains, revising their annotation as inactive USP family members. Biochemical experiments and a crystal structure of USP54 in complex with a K63-linked diubiquitin probe uncover cryptic S2 ubiquitin sites within the USP domains of both enzymes, underlying efficient cleavage within longer K63-linked chains by USP54. USP53 catalyzes K63-linkage-directed en bloc deubiquitination, a DUB activity previously not observed, which is driven by a K63-specific S2 site within its catalytic ___domain. Taken together, our data establish distinct molecular mechanisms of length-dependent decoding of K63-linked polyubiquitin chains and mechanistically connect the loss of USP53 activity to pediatric cholestasis.
Results
USP53 and USP54 are active DUBs with K63-linked polyubiquitin specificity
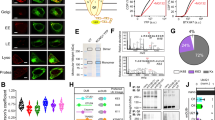
Activity-based probes are commonly used for the identification, activity profiling and structural analysis of DUBs11,22,23,38. We recently used propargylamide (PA)-based probes39,40, whose C-terminal warhead can form a vinyl thioether with the catalytic cysteines of active enzymes, to characterize ubiquitin and ubiquitin-like protein (Ubl) cross-reactivity of DUBs (Extended Data Fig. 1a,b)41. Consistent with previous studies, the ubiquitin probe facilitated the enrichment of many DUBs and HECT family E3 ligases38. Surprisingly, we also detected prominent enrichment of USP54 (Fig. 1a) even though this enzyme and its close homolog USP53 are widely reported to be catalytically inactive20,30,33. On the basis of the catalytic domains, both enzymes are the phylogenetically most distinct members of the USP family of DUBs and cluster with the deSUMOylase USPL1 (Fig. 1b)20.

a, Proteomics analysis of ubiquitin–PA probe-labeled proteins in HeLa cell lysate. The volcano plot shows the enrichment (fold change) of proteins detected in HA pulldowns from lysate treated with HA–ubiquitin–PA probe in comparison to HA–UFM1–PA as a control. Identified proteins are shown as dots, HECT E3 ligases are shown in yellow, DUBs are shown in red and USP54 is shown in blue. b, Human DUB families are shown as boxes with numbers of members given in brackets. USP53 and USP54 are annotated as inactive and comprise the most distantly related catalytic domains within the USP family, with highest similarity to the deSUMOylase USPL1. c, Domain architectures of human full-length (FL) and catalytic ___domain (CD) constructs of USP53 and USP54 used in this study. d, Probe reactivity assay with recombinant catalytic domains. HA–ubiquitin–PA was incubated with wild-type (WT) USP5320–383 and USP5421–369 or inactive mutants of USP5320–383 and USP5421–385. Probe reactivity was analyzed by SDS–PAGE and Coomassie staining. e, Ubiquitin and Ubl RhoG cleavage assay. USP5320–383 (200 nM, top) or USP5421–369 (200 nM, bottom) was added to ubiquitin–RhoG (Ub–RhoG), SUMO1–RhoG (S1–RhoG), SUMO2–RhoG (S2–RhoG), NEDD8–RhoG or ISG15–RhoG (I15–RhoG) and the fluorescence was recorded. Data are shown as the average of technical triplicates. f, Gel-based ubiquitin chain cleavage assay. Specifically linked tetraubiquitin chains (2 µM) were incubated with USP5320–383 (3 µM) or USP5421–369 (300 nM). Samples were taken after the indicated time points and cleavage activity was analyzed by SDS–PAGE and Coomassie staining. diUb, diubiquitin; triUb, triubiquitin; tetraUb, tetraubiquitin.
To test for the potential activity of USP53 and USP54, we bacterially expressed and purified their catalytic domains (Fig. 1c). Using a panel of probes including ubiquitin, different SUMO orthologs and other Ubls (Extended Data Fig. 1c,d), we observed specific reactivity with HA–ubiquitin–PA for both USP53 and USP54 (Extended Data Fig. 1c). This reactivity depended on the presence of the predicted catalytic cysteines (Fig. 1d). To directly test for ubiquitin C-terminal hydrolase activity, we used a panel of fluorogenic reagents derived from ubiquitin and Ubls (Extended Data Fig. 1e). We observed concentration-dependent and specific cleavage of ubiquitin–RhoG (rhodamine G) but not of SUMO1, SUMO2, ISG15 or NEDD8 reagents by both USP53 and USP54 (Fig. 1e and Extended Data Fig. 1f).
With the notable exception of CYLD, USP DUBs generally show unspecific cleavage or only moderate selectivity toward differently linked ubiquitin chains24,42,43. To analyze cleavage activity on isopeptide linkages, we assayed a tetraubiquitin panel. To our surprise, USP53 and USP54 cleaved K63-linked chains with remarkable specificity (Fig. 1f). This finding was highly unexpected as no other human USP DUB shows specific cleavage activity of a single linkage. For USP54, no cleavage was observed for any other linkage, whereas USP53 minimally cleaved K11-linked and K48-linked tetraubiquitin chains at longer time points. Collectively, these experiments demonstrate that human USP53 and USP54 are active enzymes with a specificity for K63-linked polyubiquitin (Fig. 1 and Extended Data Fig. 1).
Mutations in USP53 cause loss of catalytic activity
Recently, 14 reports have linked pediatric cholestasis to biallelic mutations in the USP53 gene (Supplementary Table 1)34,35,36,37. However, the falsely presumed enzymatic inactivity of the USP53 protein has so far prevented a mechanistic analysis of the underlying pathologies. We compiled all missense mutations in USP53 and found a clustering within its catalytic ___domain (Fig. 2a). We obtained the USP53 catalytic ___domain with the disease-associated substitution R99S through bacterial expression and found it to be completely inactive toward K63-linked triubiquitin chains (Fig. 2b) and ubiquitin–RhoG (Fig. 2c and Extended Data Fig. 2a). Consistently, USP54 with an equivalent substitution of R100A was virtually inactive in the same assays (Fig. 2b and Extended Data Fig. 2b). In addition, we tested the G31S, C303Y and H132Y USP53 substitutions in the same assays (Fig. 2c,d and Extended Data Fig. 2) and found them to be strongly impaired in catalysis, particularly in ubiquitin chain cleavage (Fig. 2d). All mutant proteins were folded as assessed in thermal shift assays (TSAs) (Extended Data Fig. 2c,d) and were capable of recognizing ubiquitin, as evident from reduced but retained reactivity toward a ubiquitin probe (Extended Data Fig. 2e–g). These data demonstrate that disease-associated mutations abrogate the enzymatic activity of USP53. More broadly, the data implicate the loss of USP53 DUB activity as the cause of USP53-mediated pathology.

a, Schematic of human USP53 with amino acid substitutions associated with cholestasis or hearing loss. Single amino acid changes cluster in the catalytic ___domain. b, Gel-based ubiquitin chain cleavage assay. K63-linked triubiquitin (3 µM) was incubated with USP5320–383 (2 µM, left) or USP5421–369 (300 nM, right). Cleavage was analyzed by SDS–PAGE and Coomassie staining. c, Ubiquitin–RhoG cleavage assay. Ubiquitin–RhoG was incubated with USP5320–383 (WT or indicated mutants; raw data in Extended Data Fig. 2a) and the fluorescence was recorded. Observed rate constants (kobs, shown as the mean ± s.e.m.) were plotted over enzyme concentrations to obtain catalytic efficiencies (shown as the mean ± s.e.m.). d, Gel-based ubiquitin chain cleavage assay with additional USP53 mutations, shown as in b. e, Fluorescence-based triubiquitin cleavage assay. A K63-linked triubiquitin substrate (3 µM) was used, in which the proximal Ub1–75-CA was conjugated to maleimide–TAMRA (triubiquitin–TMR). Cleavage by USP5320–383 (3 µM) or USP5421–369 (300 nM) was analyzed by SDS–PAGE, in-gel fluorescence and Coomassie staining. The major cleavage position is indicated with an arrow. S2, S1 and S1′ ubiquitin-binding sites in the DUB were assigned consistent with the observed cleavage products. f, Schematic of ubiquitin substrates used in g. Cleavage sites are indicated by arrows. Ubiquitin-binding sites, which lead to cleavage when engaged, are given for the respective substrates. TAMRA is shown as a purple star. Structures of the substrates are shown in Extended Data Fig. 3. g, Fluorescence polarization cleavage assays. Substrates were incubated with USP5320–383 (top) or USP5421–369 (bottom) and fluorescence anisotropy was recorded (raw data in Extended Data Fig. 3i). Observed rate constants (kobs, shown as the mean ± s.e.m.) were plotted over enzyme concentrations to obtain catalytic efficiencies (shown as the mean ± s.e.m.).
Ubiquitin chain length decoding by USP53 and USP54
Strikingly, diubiquitin chains accumulated during K63-linked polyubiquitin chain cleavage by both enzymes (Figs. 1f and 2b), as confirmed in further time-resolved assays (Extended Data Fig. 3a). These data suggest an unusual ubiquitin chain length preference of both enzymes. The linkage specificity of DUBs is typically facilitated by S1 and S1′ ubiquitin-binding sites, with the preferred cleavage of longer chains being facilitated through additional ubiquitin-binding sites (Fig. 2e)15,22. To determine whether USP53 and USP54 contain S2′ or S2 sites, we prepared fluorescently labeled K63-linked triubiquitin substrates allowing the separate detection of fluorescent species (Extended Data Fig. 3b–e). As we found USP54 activity to be impaired by the presence of the FlAsH dye (Extended Data Fig. 3c), we generated K63-linked triubiquitin–TAMRA in which the proximal ubiquitin is fluorescently labeled through a C-terminal cysteine (Extended Data Fig. 3d,e). Assaying the cleavage of the reagent by USP53 and USP54 at early time points revealed preferential conversion into nonfluorescent diubiquitin and fluorescent ubiquitin–TAMRA (Fig. 2e). This cleavage pattern suggested the existence of an S2 ubiquitin-binding site in both enzymes, as depicted in Fig. 2e and Extended Data Fig. 3b. A potential fourth ubiquitin-binding site was excluded upon assaying an equivalent K63-linked tetraubiquitin–TAMRA reagent (Extended Data Fig. 3f,g).
We next aimed to quantify the contributions of the S1, S1′ and S2 ubiquitin-binding sites to USP53 and USP54 activity by using monoubiquitin–KGTAMRA together with K63-linked diubiquitin and triubiquitin reagents in fluorescence polarization assays (Fig. 2f and Extended Data Fig. 3h). Cleavage activity was measured as the decline in anisotropy over time (Extended Data Fig. 3i) and was quantified through observed rate constants, yielding catalytic efficiencies of USP53 and USP54 for each substrate (Fig. 2g). USP53 showed poor cleavage of ubiquitin–KGTAMRA and an even lower turnover of the diubiquitin substrate. Addition of a third ubiquitin led to an 18-fold increase in catalytic efficiency (Fig. 2g). This behavior could be explained by competing and catalytically unproductive binding of the monoubiquitin and diubiquitin substrates to a high-affinity S2 binding site. In contrast, USP54 showed an even poorer cleavage of ubiquitin–KGTAMRA but a more than 20-fold increase in catalytic efficiency for diubiquitin–TAMRA. Additional engagement of the S2 site by triubiquitin–TAMRA led to a further increase in catalytic efficiency (Fig. 2g), consistent with the results of the tetraubiquitin panel (Fig. 1f).
USP53 displays linkage-directed deubiquitination activity
While DUB activity is typically assayed with free chains, the majority of cellular polyubiquitin is conjugated to protein substrates1,2. To test for distinct activity profiles of USP53 and USP54 in the context of protein-bound ubiquitin chains, we generated a panel of ubiquitinated model substrates including isopeptide-linked monoubiquitinated GFP and GFP conjugated to K48-linked or K63-linked chains of different length (Fig. 3a). The isopeptide linkage of ubiquitin to lysine acylation using conjugating enzymes (LACE)-tagged GFP was generated with recently reported methodology44 and chains were subsequently assembled. Consistent with the fluorescence polarization assays, USP54 did not turn over GFP–ubiquitin but shortened K63-linked ubiquitin chains to leave a single ubiquitin on GFP (Fig. 3b). This finding highlights the importance of its S1′ ubiquitin-binding site. Substrate turnover increased with chain length, consistent with the identified S2 site.

a,b, Cleavage assay with isopeptide-linked, K63-linked monoubiquitinated or polyubiquitinated substrates. GFP–ubiquitin substrates (1 µM) were incubated with USP5320–383 (500 nM; a) or USP5421–369 (300 nM; b). In-gel fluorescence was used to visualize GFP species, while Coomassie staining of the same gel was used to visualize ubiquitin chains. Arrows indicate cleavage sites. c, USP53 depletion analysis. CaCo-2 cells stably transduced as indicated were analyzed by western blotting after treatment with doxycycline for 72 h. d, Schematic illustration of the diglycine ubiquitinome profiling to identify USP53 substrates. e, Volcano plot showing log2 fold changes of diglycine (ubiquitinated) peptides upon depletion of USP53 in CaCo-2 cells. Proteins linked to phenotypes similar to USP53 mutations are indicated in red. Diglycine sites were unambiguously identified. f, Model linking USP53-associated phenotypes to USP53-modulated ubiquitination of tricellular tight junction proteins (with domains, ubiquitination sites and residue numbers given). PM, plasma membrane. g, MARVELD2 ubiquitination analysis through OtUBD pulldown. CaCo-2 shRNA5 (USP53) cells were treated as in c, lysates were enriched with the high-affinity ubiquitin binder OtUBD and samples were analyzed by western blotting. The triangle highlights MARVELD2 modified with two ubiquitin moieties, emerging upon USP53 depletion. h, MARVELD2 polyubiquitination analysis with K63-specific tUIMRap80 (Rx3A7) TUBE pulldown. i, Analysis of protein levels for samples processed in h. j. Illustration of the workflow used to generate eluates of OtUBD pulldowns for UbiCRest assays. Acidic conditions allowed the elution of ubiquitinated proteins, with the biotinylated OtUBD reagent being retained on streptavidin beads. k, UbiCRest assay. OtUBD eluates prepared as described in j were treated with the indicated DUBs for 1 h at 37 °C and analyzed by western blot. l, UbiCRest assay as in k with USP5320–383 (0.5, 2 and 5 µM) and USP5421–369 (0.3, 1 and 3 µM), showing en bloc deubiquitination of cellular MARVELD2 by recombinant USP53. The results after 2-h incubation are shown in Extended Data Fig. 4k.
A strikingly different picture emerged for USP53, which preferentially cleaved polyubiquitin completely off GFP in a K63-linkage-dependent manner as evident from gel-based cleavage assays (Fig. 3a and Extended Data Fig. 4a–d). We followed the conversion of the GFP substrates using in-gel fluorescence and observed the emergence of free GFP. Coomassie staining of the same gel visualized free ubiquitin chains generated by USP53. Importantly, GFP–diubiquitin was converted into GFP and free diubiquitin (Fig. 3a). The earliest time points of the GFP–triubiquitin and GFP–tetraubiquitin assays showed the emergence of free triubiquitin and tetraubiquitin, respectably, which were later broken down into shorter chains consistent with the tetraubiquitin panel. The formation of free diubiquitin and triubiquitin chains was validated by intact protein mass spectrometry, which unequivocally confirmed the surprising en bloc deubiquitination activity of USP53 (Extended Data Fig. 4a-b). The emergence of low amounts of monoubiquitinated GFP in GFP–triubiquitin and GFP–tetraubiquitin assays demonstrates that substrate deubiquitination is not exclusive and cleavage in substrate-bound chains can take place in K63-linked chains longer than two ubiquitin moieties. Importantly, GFP conjugated to K48-linked diubiquitin or triubiquitin chains was not cleaved under identical conditions (Extended Data Fig. 4c,d), demonstrating linkage specificity of this activity.
Previously reported ubiquitin linkage-specific DUBs were described to operate strictly within ubiquitin chains. To our knowledge, an enzyme for which ubiquitin chains of a defined linkage direct deubiquitination activity for the preferred en bloc removal of this modification from substrates has not been described2,14,16,17,20,45,46. Our data strengthen the hypothesis that the en bloc deubiquitination activity of USP53 is facilitated by ubiquitin binding to both the S1 and S2 sites (Extended Data Fig. 4d)20,45. Taken together, these results show that USP53 and USP54 are active DUBs with intriguing K63 linkage and chain length specificities and that USP53 possesses unique K63-linkage-directed deubiquitination activity.
Tricellular tight junction proteins are cellular substrates of USP53
To validate this activity in a cellular context and investigate possible consequences related to USP53 loss, we generated stably transduced CaCo-2 cell lines carrying a doxycycline-inducible short hairpin RNA (shRNA) against USP53. CaCo-2 is a widely used human model cell line for the study of epithelial cell barriers and USP53 shRNA induction led to effective depletion of the USP53 protein (Fig. 3c). We next assessed the effect of USP53 depletion on the ubiquitination status of proteins by ubiquitinome analysis, carried out by enrichment of diglycine-modified peptides and data-independent acquisition (DIA) mass spectrometry analysis (Fig. 3d)3. The top four ubiquitination sites, whose abundance increased upon USP53 depletion, included K412 of MARVELD2 (also termed tricellulin) and K638 of LSR (also termed angulin-1) (Fig. 3e). Markedly, these two sites are within the cytoplasmic domains of MARVELD2 and LSR, for which genetic mutations in humans lead to the same phenotypes as USP53 mutation (Fig. 3f)36,47. Both membrane-spanning proteins are core components of specialized cell–cell contact sites termed tricellular tight junctions48, with LSR recruiting MARVELD2 to tricellular tight junctions49, and are essential for epithelial barrier function47,50,51.
To corroborate ubiquitination, we used the high-affinity ubiquitin-binding ___domain OtUBD52,53 in a denaturing pulldown of cellular ubiquitin (Extended Data Fig. 4e,f). Blotting for MARVELD2 revealed its increased ubiquitination upon USP53 depletion (Fig. 3g). We observed the strongest change in a species whose molecular weight fits to MARVELD2 modified with two ubiquitin moieties, which we assigned as K63 diubiquitination on the basis ofour biochemical data (Figs. 1f and 3a and Extended Data Fig. 4c). The same change in MARVELD2 ubiquitination was observed using a cell line in which USP53 was depleted through a different shRNA sequence but was not observed in parental CaCo-2 cells upon treatment with doxycycline (Extended Data Fig. 4g,h).
To directly assess the regulated ubiquitin linkage type, while remaining at endogenous ubiquitin levels, we analyzed the same samples with tandem ubiquitin-binding entities (TUBEs) (Extended Data Fig. 4e,f)54. Their avidity-based mechanism entails that longer chains are enriched more efficiently than short chains. Pulldown of polyubiquitinated proteins with a high-affinity nonselective 4xUBAUBQLN1 reagent showed only a modest increase in overall polyubiquitinated MARVELD2 upon USP53 depletion (Extended Data Fig. 4i,j). However, enrichment with a K63-specific tUIMRap80 reagent55 revealed a strong increase in K63-linked polyubiquitinated MARVELD2 (Fig. 3h). Importantly, global levels of MARVELD2 remained unchanged (Fig. 3i), which supports the notion that nondegradative ubiquitination is being regulated by USP53.
To test whether USP53 can deubiquitinate MARVELD2, we devised a workflow in which ubiquitinated proteins are enriched and then separated from the high-affinity OtUBD reagent by elution under acidic conditions. Subsequent elevation of the pH in the eluates and incubation with recombinant DUBs allowed UbiCRest assays15 (Fig. 3j). We observed that the nonspecific DUB USP2 removed all modifications on MARVELD2, confirming ubiquitination. The K63-specific DUB AMSH* but not the K48-specific DUB OTUB1* increased the amount of monoubiquitinated MARVELD2, consistent with cleavage within K63-linked chains (Fig. 3k). In contrast, incubation with USP53 but not USP54 led to an increase in free MARVELD2 and a concomitant decrease in the K63-linked diubiquitinated species, whereas the monoubiquitinated species remained unchanged (Fig. 3l and Extended Data Fig. 4k). These results are fully consistent with the biochemically identified K63-linkage-directed en bloc deubiquitination activity and, in sum, validate MARVELD2 as a cellular substrate of USP53.
A covalent USP54~K63-linked diubiquitin–PA complex for crystallization
We next set out to investigate the mechanism of K63 selectivity and how these DUBs recognize chains. To visualize the additional S2 binding site, we aimed to solve a crystal structure of USP53 or USP54 in complex with K63-linked diubiquitin–PA. We prepared this isopeptide-linked probe by enzymatically assembling ubiquitin K63R onto ubiquitin–PA (Fig. 4a). A similarly designed K48-linked diubiquitin probe comprising a triazole linkage was previously used for the identification of a K48-specific S2 ubiquitin-binding site of a severe acute respiratory syndrome coronavirus (SARS-CoV) papain-like protease (PLpro) enzyme16,56. We used our probe alongside ubiquitin–PA to assess ubiquitin engagement in the prepared complexes (Fig. 4b). In thermal stability assays, diubiquitin–PA binding stabilized USP53 and USP54 more than ubiquitin–PA, indicating occupation of the S2 site (Fig. 4c and Extended Data Fig. 5a).

a, Schematic of the generation of a K63-linked diubiquitin probe, which was enzymatically assembled from ubiquitin K63R and ubiquitin–PA. IEC, ion-exchange chromatography. b, Schematic of catalytic DUB domains after reaction with ubiquitin–PA or diubiquitin–PA probes, illustrating ubiquitin engagement in samples used in c,d. S1′, S1 and S2 ubiquitin-binding sites are labeled. The active site cysteine is depicted as a star. c, Protein stability assessment. USP5320–368 and USP5421–369 were labeled with ubiquitin–PA or diubiquitin–PA probes and the stability of protein samples was analyzed by thermal shift analysis. Melting temperatures (Tm) are shown for technical replicates, indicating the contribution of the S2 site. d, Purified DUB samples. USP5421–369 and USP5421–369 conjugated to ubiquitin–PA and USP5421–369 conjugated to K63-linked diubiquitin–PA were analyzed by SDS–PAGE and Coomassie staining. e, Crystal structure of USP54 conjugated to K63-linked diubiquitin–PA. In the observed complex, two USP54 molecules (blue and red) engage two K63-linked diubiquitin–PA molecules (gold and wheat) crosswise. The two isopeptide bonds between the diubiquitin–PA molecules are highlighted and are shown as sticks. f, SEC–MALS experiment of the catalytic ___domain of USP5421–369 alone (blue), in complex with ubiquitin–PA (brown) or in complex with K63-linked diubiquitin–PA (yellow), demonstrating monomeric species in solution. g, Schematic depiction of the organization of USP54 conjugated to K63-linked diubiquitin–PA in the crystal and in solution.
We prepared and purified several probe-conjugated DUB samples for extensive crystallization trials (Fig. 4d and Extended Data Fig. 5b,c). Following construct optimization, we obtained crystals for the most stable complex, USP54 conjugated to diubiquitin–PA (Fig. 4c,d), allowing the determination of a structure to 2.5-Å resolution by X-ray crystallography (Supplementary Table 2). Its asymmetric unit contained four copies of USP54 conjugated to diubiquitin–PA (Extended Data Fig. 5d,e), of which two copies formed an identical arrangement (Extended Data Fig. 5f). Excellent electron density covering the entire complex revealed that the two ubiquitin moieties of the probe engaged two different USP54 copies with symmetrical binding in trans configuration (Fig. 4e and Extended Data Fig. 5e). To assess the solution state of this complex, size-exclusion chromatography with multiangle light scattering (SEC–MALS) measurements were performed, proving that all USP54 samples are monomeric in solution (Fig. 4f). These results allowed the assignment of the dimeric arrangement of USP54 conjugated to diubiquitin–PA as a crystal artifact, likely derived from stabilization of the finger ___domain of USP54. We propose that, in solution, the complex of USP54 conjugated to diubiquitin–PA exists as a monomer and that USP54 engages K63-linked ubiquitin chains through its S1 and the S2 sites in cis configuration (Fig. 4g).
USP54 contains a USP catalytic ___domain with a weakened S1 site
USP54 adopts the USP DUB catalytic ___domain fold consisting of hand, thumb and finger subdomains (Extended Data Fig. 6a). A model for the solution state illustrated binding of the distal ubiquitin to an S2 site located at the back of the USP fingers subdomain (Fig. 5a). An identical orientation was observed for all four USP54 molecules within the asymmetric unit (Extended Data Fig. 6b). Moreover, we found the C-terminal residues of the ubiquitin in the S2 site to be in close proximity to the K63 side chain of the S1 ubiquitin (Fig. 5a). This is consistent with an in-solution structure in which the isopeptide bond is placed on top of the fingers without the need for reorientation of the ubiquitin moieties.

a, Solution arrangement of USP54 conjugated to K63-linked diubiquitin–PA. Cartoon representation depicting USP5421–369 (gray) with ubiquitin moieties bound in its S1 (wheat) and S2 (gold) binding sites. Zinc atoms are shown as gray spheres. b, Close-up view of the catalytic triad. Indicated distances are visualized by dotted lines. c, Sequence alignment of residues forming BL2 in USP DUBs. Catalytic histidines are colored red, while conserved residues are colored green. Residues or gaps in USP54 and USP53, which are unique across the entire human USP family, are highlighted with a box. Residues strongly conserved in the USP DUB family, which led to the initial mischaracterization of USP53 and USP54 as inactive, are highlighted in bold. Secondary-structure elements and numbering are according to USP54. d, Left, close-up view of BL1 and BL2 of USP54. Right, superposition with the corresponding residues in USP2 (PDB 2IBI, yellow) and USP14 (PDB 2AYO, purple), illustrating the atypical shortening of both loops in USP54. e, Left, close-up view of the unique loop of USP54 close to the catalytic cysteine. Right, superposition of the Cys loop with the corresponding residues in USP2 (yellow) and USP12 (PDB 5L8W, magenta). f, Sequence alignment of residues forming the Cys loop in USP DUBs (annotation as in c). Sequences of USP53 and USP54 constructs used to study the Cys loop are shown below with changes marked in violet. g, Gel-based ubiquitin chain cleavage assay. K63-linked triubiquitin chains (3 µM) were incubated with USP5320–383 (3 µM) or USP5421–369 (300 nM) as either WT or Cys loop mutant (labeled as LGNT) (sequences in f) for the indicated time points. Cleavage activity was analyzed by SDS–PAGE and Coomassie staining.
We next analyzed the USP54 catalytic ___domain and its engagement of the S1 ubiquitin. A canonical catalytic triad was positioned in an active conformation, with the warhead of the diubiquitin–PA probe covalently linked to the catalytic cysteine (Fig. 5b). The cleavage activity on K63-linked ubiquitin chains was abolished for catalytic cysteine and histidine mutants (Extended Data Fig. 6c,d). Moreover, the structure revealed three coordinated zinc ions, which comprised two positions within the thumb subdomain in addition to the common position at the tip of the fingers (Extended Data Fig. 6a). One of these was identified in human USP36 (ref. 41), whereas coordinating residues for the second zinc are not present in other human USP DUBs32. The positions of the three ions loosely resemble those in the USP ___domain within the yeast SAGA DUB module57,58.
Engagement of the S1 ubiquitin by USP DUBs typically relies on (1) hydrophobic contacts to the F4 patch by the frontside of the finger subdomain; (2) a hydrophobic contact of the I36 patch by blocking loop 1 (BL1); and (3) guiding of the ubiquitin C-terminal residues through the active site cleft by blocking loop 2 (BL2)41,59. In USP54, the S1 ubiquitin displayed a ~30° rotation in comparison to the common binding mode (Extended Data Fig. 7a) and differed in all three main interaction sites. Firstly, the hydrophobic residues normally contacting F4 were missing, leaving this patch largely solvent exposed (Extended Data Fig. 7b). Secondly, I36 of ubiquitin was solvent exposed (Extended Data Fig. 7c), as BL1 appeared absent, with corresponding residues adopting a unique helix (α7.5) oriented away from ubiquitin. This arrangement has not previously been observed for USP DUBs (Fig. 5c,d). Thirdly, BL2 of USP54 was truncated (Fig. 5c,d), leaving amide bonds in the ubiquitin C-terminal residues without the otherwise occurring zip-like hydrogen bonding (Extended Data Fig. 7d). Collectively, these differences, which appear to be shared by USP53 and USP54 according to the primary sequence, suggest a weakened S1 ubiquitin-binding site. This is consistent with the comparably weak activity of both DUBs on monoubiquitin substrates (Extended Data Fig. 1f and Fig. 2g).
Another feature of USP53 and USP54, which is unique among human USP DUBs, is an elongated Cys loop directly before the catalytic cysteine (Fig. 5e,f). The additional residues are flanked by two asparagine residues conserved in the USP family32, of which N35 acts as an oxyanion hole. Substitution of this residue in both USP53 and USP54 verified its importance for catalytic activity (Extended Data Fig. 6e). Moreover, amino acid changes in the unique Cys loop sequence (EPGQNSC) of both DUBs into the commonly observed sequence (LGNTC) completely abolished cleavage of K63-linked chains (Fig. 5g).
Mapping all cholestasis-related single amino acid substitutions in USP53 (Fig. 2a) on a homology model based on our USP54 structure illustrated that several substituted residues are either part of the above-described zinc fingers (explaining the reduced stability and impaired catalytic activity of the H132Y protein) or are located around the active site (Extended Data Fig. 8a–e). The model indicates how G31S and C303T lead to structural perturbations in immediate vicinity of the catalytic residues (Extended Data Fig. 8d,e), explaining the impaired catalytic activity of USP53 proteins with these substitutions (Fig. 2c,d). A closer inspection of R99 in USP53 or of the equivalent residue R100 in the USP54 structure revealed critical roles in coordinating the unique Cys loop to the switching loop (Extended Data Fig. 8b,c). Taken together, these data demonstrate the importance of the unique active site arrangement in both USP DUBs for catalytic activity and enable the structural rationalization of USP53 mutations associated with disease (Supplementary Table 1).
A cryptic S2 site within the USP fingers engages K63-linked chains
The structure of USP54 conjugated to diubiquitin–PA uncovered an S2 ubiquitin-binding site within its USP ___domain (Figs. 5a and 6a and Extended Data Fig. 8f), consistent with biochemical assay data. This interface is centered on the backside of the fingers subdomain and is governed by hydrophobic interactions with the I44 patch of ubiquitin. This main protein interaction site of ubiquitin comprises I44 in addition to L8, V70 and H68 (ref. 1). The central residue in USP54 at the contact site is F161 (Fig. 6b), which corresponds to Y160 in USP53, suggesting similar ubiquitin engagement. Notably, these hydrophobic residues on the backside of the USP fingers ___domain are strictly conserved in USP53 and USP54 orthologs (Extended Data Fig. 8g,h), whereas other human USP DUBs have varying and typically polar residues at these positions (Fig. 6c).

a, Structure of USP54 conjugated to K63-linked diubiquitin–PA. b, Interface of ubiquitin and USP54 S2 site. c, Sequence alignment of residues forming the S2 site in USP54 and USP53. Cys-x-x-Cys motifs at the tip of the fingers (blue), conserved residues (green), residues unique in USP53 and USP54 within the human USP family (box) and residues annotated in b (arrows) are highlighted. d, Gel-based polyubiquitin cleavage assays. K63-linked ubiquitin chains (2 µM) were incubated with WT USP5421–369 or the S2 site mutant F161K (both at 300 nM). Substrate consumption was quantified by densitometry, normalized to initial intensities. Data are shown as the average ± s.d. of three independent replicates. e, Gel-based polyubiquitin cleavage assays of USP53, shown as in d. WT USP5320–383 or the S2 site mutant Y160K was used (2 µM). f, Catalytic efficiencies of USP54 proteins obtained from fluorescence polarization assays (substrates shown in Fig. 2d). Raw data are shown in Extended Data Fig. 9d,e and the catalytic efficiencies of WT protein are repeated from Fig. 2g. Data are shown as the mean ± s.e.m. g, Catalytic efficiencies of USP53 proteins, analyzed as in f. Efficient catalysis of USP53 is dependent on its S2 site. h, Schematic of ubiquitin processing by DUBs. K63-linkage-directed deubiquitination by USP53 bridges canonical DUB categories. The structural mechanisms for polyubiquitin length and linkage specificity in DUBs are shown in Extended Data Fig. 10.
We next sought to assess the importance of the S2 site for cleavage activity on K63-linked ubiquitin chains. An F161K substitution abrogated binding of the distal ubiquitin through disruption of the S2 site (Extended Data Fig. 9a–c). This substitution led to a decrease in cleavage of triubiquitin and tetraubiquitin chains by USP54 (Fig. 6d), visualized through quantitative gel-based cleavage assays. Importantly, the substitution did not influence the cleavage of K63-linked diubiquitin, demonstrating that binding of ubiquitin chains to the S1 and S1′ sites is not affected by a change in the S2 site (Fig. 6d). Strikingly, an equivalent Y160K substitution in USP53 drastically reduced the cleavage of triubiquitin and tetraubiquitin chains (Fig. 6e), indicating that efficient catalysis for USP53 is driven through the S2 site.
To confirm these findings, we assessed both mutant proteins in fluorescence polarization assays (Fig. 6f,g and Extended Data Fig. 9d,e). Consistently, amino acid changes impacting the S2 sites of USP53 or USP54 did not change the catalytic activities on monoubiquitin and diubiquitin substrates. However, triubiquitin cleavage activities were dampened. USP53 Y160K showed cleavage activity barely above background (Fig. 6g), proving the critical importance of the S2 site for catalysis. The more dramatic effect of the amino acid changes in USP53 compared to USP54 is consistent with the higher activity of USP54 on diubiquitin substrates.
Collectively, these data demonstrate that both USP53 and USP54 possess an additional K63-linkage-specific S2 ubiquitin-binding site embedded into their USP catalytic domains. Engagement of the ubiquitin I44 patch by the back of their fingers subdomains is unique within the USP family of DUBs and mechanistically explains their preference for K63-linked ubiquitin chains.
Discussion
We here revise the annotation of USP53 and USP54 from inactive pseudo-DUBs32 to active DUBs and report in USP53 the discovery of K63 ubiquitin linkage-directed deubiquitination activity. This unprecedented mode of human DUB activity expands the previously established categories of canonical USP DUBs, which typically cleave isopeptide linkages promiscuously, and of linkage-specific DUBs, which edit chains but do not deubiquitinate substrates (Fig. 6h)20. In USP53, recognition of K63-linked chains through an S2 site embedded within its catalytic ___domain is a prerequisite for efficient catalysis. USP53 shares with its homolog USP54 a weakened S1 ubiquitin-binding site through truncations in BL1 and BL2, which explain their comparably poor reactivity on monoubiquitin substrates.
Catalysis driven by an S2 ubiquitin-binding site has previously been reported for PLpro enzymes of SARS-CoV16 and SARS-CoV-2 (ref. 17), albeit with K48 linkage specificity. These enzymes possess N-terminal extensions to their USP-like catalytic ___domain to recognize ISG15 and K48-linked polyubiquitin (Extended Data Fig. 10), for which hydrophobic recognition of the distal ubiquitin is critical60. While K48-linkage-directed substrate deubiquitination by these enzymes has been evaluated in vitro45,60, evidence for this activity on cellular substrates is currently not available. Structural16 and biochemical45 studies on SARS-CoV PLpro concluded that cleavage within K48-linked polyubiquitin is preferred over K48-linkage-directed substrate deubiquitination. This is in contrast to USP53, which, when evaluated with purified substrate-bound ubiquitin chains, preferred the en bloc removal of K63-linked polyubiquitin over cleavage within the chain (Fig. 3a and Extended Data Fig. 4a,b). Moreover, cellular investigation revealed changes in K63-linked ubiquitination of MARVELD2, the emergence of a diubiquitin-modified protein and changes in ubiquitination site abundance in MARVDEL2 and LSR upon USP53 deletion (Fig. 3). These findings are consistent with the unique activity we characterize and, together, set it apart from canonical linkage-specific DUBs. Importantly, deubiquitination by USP53 is greatly accelerated by K63-linked but not by K48-linked polyubiquitin, as expected from the structure-guided biochemical analysis of its K63-specific S2 site (Fig. 6). At high concentrations or long incubation times, low cleavage activity on other linkage types can be detected for USP53. The same has also been observed for the M1/K63-specific enzyme CYLD42 and is in line with the typically broad catalytic scope of USP family members. This explains why linkage-directed deubiquitination activity could emerge in a USP-fold enzyme with weakened S1 sites and an added S2 site. Investigation of a potential S1′ site in future work could illuminate molecular determinants for en bloc substrate deubiquitination versus cleavage within chains.
En bloc removal of ubiquitin chains from substrates has previously been demonstrated for proteasome-resident DUBs PSMD14 and USP14, which is independent of the chain linkage type20,46,61. Notably, the mechanism of K63-linkage-directed deubiquitination of USP53 is also distinct from the established activity of USP5 (ref. 62), which drives efficient cleavage of free ubiquitin chains through an S1′ site outside of its catalytic ___domain and irrespective of the linkage type. Thus, the mechanism of USP53 functionally expands the described DUB activities in human cells.
USP54 is also an atypical USP DUB family member with K63 specificity and a preference for longer chains. Preferred cleavage of longer chains has been reported for (1) ZUFSP for K63 linkages6,11,23,25, with a yet unknown mechanism; (2) MINDY DUBs with K48-specific S2′ through S4′ ubiquitin sites14,22; (3) the SARS-CoV PLpro enzyme with a K48-specific S2 site in the N-terminal extension of its catalytic ___domain16,17; and (4) OTUD2 with a K11-specific S2 site15. The mode of ubiquitin chain recognition by USP54 is distinct from all abovementioned examples and from the K63-specific DUBs AMSH26 and CYLD42 (Extended Data Fig. 9f), which contain S1′ but not S2 sites within their catalytic domains (Extended Data Fig. 10). While K63-linked chains of 2–4 ubiquitin moieties exist on substrate proteins in cells7 and can be recognized in a length-dependent manner12, how K63 polyubiquitin chain length can be decoded by DUBs on the molecular level was previously not known. These findings will guide the assessment of ubiquitin-dependent roles of cellular, full-length USP54 and stimulate research into the chain-length-dependent fine-tuning of ubiquitin signaling.
Mutations in USP53 have clinically been related to bile acid transport disorders, a subgroup of cholestasis63 also comprising cases with mutations in TJP2 (tight junction protein 2)64 and in LSR36. Some persons with congenital USP53 alterations also presented with hearing loss34,37, consistent with the mouse USP53 phenotype30. Notably, deafness is also caused by mutations in genes encoding cell–cell junction proteins including claudins, ILDR1 and the MARVELD family members occludin and tricellulin (MARVELD2)47,51. USP53 was shown to localize to cell–cell barriers in mouse inner ear cells and to interact with TJP1 and TJP2 (ref. 30). Consistent with our biochemical and structural data and the proposed model for USP53 activity, we found elevated ubiquitination on the two core components of tricellular junctions MARVELD2 and LSR upon knockdown of USP53. The combined use of diglycine proteomics, different ubiquitin enrichment techniques and UbiCRest-like assays validates these as USP53 substrates and synergizes with recent advances in the assessment of ubiquitin chain architecture3,7,65 These substrates are supported by coinciding phenotypes of deafness and cholestasis36,47, consistent with the USP53 mouse phenotype30,47. Collectively, the results converge into a model in which loss of the USP53 catalytic activity reported herein causes pediatric cholestasis (Fig. 3).
Our work critically guides the investigation of ubiquitin-dependent cellular roles of USP53 and USP54. Our structural insights will aid the molecular investigation of both enzymes and provide a framework for the understanding of USP53 disease-causing mutations. Moreover, our results demonstrate how ubiquitin chains can lead to en bloc protein deubiquitination by a human DUB in a chain-linkage-dependent manner. This unique activity enables the deubiquitination of polyubiquitinated proteins while preserving monoubiquitin modifications on the same substrate. Lastly, our data stimulate the re-evaluation of known DUBs with isopeptide-linked polyubiquitin substrates, which represent endogenous structures more closely and complement frequently used panels of free ubiquitin chains, in search for perhaps other enzymes featuring linkage-directed deubiquitination activity.
Methods
Cloning and constructs
Catalytic ___domain constructs of human USP53 (residues 20–368 and 20–383; UniProt Q70EK8) and USP54 (residues 21–369 and 21–385; UniProt Q70EL1) were cloned from Escherichia coli codon-optimized DNA into the pOPINK vector for bacterial expression (N-terminal His6-GST-3C-tag) with In-Fusion cloning (Takara Clonetech). Site-directed mutagenesis was carried out through PCR according to the QuikChange protocol using Phusion polymerase (New England BioLabs (NEB)).
DNA sequences coding for human ubiquitin (Uniprot P0CG47) or human Ubls SUMO1, SUMO2, SUMO4, SUMO5, ISG15, UFM1, FUBI, URM1 and DWNN (UniProt P63165, P61956, Q6EEV6, G2XKQ0, P05161, P61960, P62861, Q9BTM9 and Q7Z6E9, respectively) were cloned into the pTXB1 vector (NEB) with N-terminal HA tags as described previously41. Ubiquitin and ubiquitin variants were expressed from pET17b.
Constructs for mouse E1 (mE1), UBE2R1, UBE2V1, UBE2N, UBE2S-UBD, NleL, UBE2L3/UbcH7, AMSH* and OTUB1* were used as described previously66. UBE2S and UBE2N were cloned from plasmids encoding UBE2S-UBD and UBE2N into pOPINK.
Enhanced monomeric GFP was cloned from a previously described vector67 with a C-terminal LACE tag (GSGPRKVIKMESEE) (GFP-C-LACE-tag) into pOPINK.
To obtain shRNA-encoding vectors, the Tet-pLKO-puro plasmid was sequentially digested by AgeI and EcoRI followed by ligation with annealed sense and antisense shRNA oligonucleotides using T4 DNA Ligase (NEB). The following sequences were used:
shRNA1 (USP53), forward: CCGGGGTCACATTGATATGAAATATCTCGAGATATTTCATATCAATGTGACCTTTTTG
shRNA1 (USP53), reverse: AATTCAAAAAGGTCACATTGATATGAAATATCTCGAGATATTTCATATCAATGTGACC
shRNA5 (USP53), forward: CCGGCACTTCCCTCAGATAACATAACTCGAGTTATGTTATCTGAGGGAAGTGTTTTTG
shRNA5 (USP53), reverse: AATTCAAAAACACTTCCCTCAGATAACATAACTCGAGTTATGTTATCTGAGGGAAGTG
shRNA nontargeting control (NTC), forward: CCGGAGTGAACGAGTAAGATAACCCTCGAGGGTTATCTTACTCGTTCACTTTTTTG
shRNA NTC, reverse: AATTCAAAAAAGTGAACGAGTAAGATAACCCTCGAGGGTTATCTTACTCGTTCACT
Constructs for ubiquitin enrichment reagents were cloned into pOPINB adding N-terminal Avi tags for the pan-polyubiquitin–TUBE (4xUBAUBQLN1) and the K63-linked polyubiquitin–TUBE (tUIMRap80/Rx3A7) and an N-terminal cysteine for OtUBD.
Protein expression
Rosetta 2(DE3)pLacI cells were used for bacterial protein expressions after transformation with the respective plasmids (apart from ubiquitin enrichment proteins, which were expressed in BL21(DE3) cells). The 2xTY medium supplemented with appropriate antibiotics was inoculated in a 1:100 ratio from an overnight culture and cells were grown shaking at 37 °C until an optical density of 0.7–1.0 was reached. After cooling to 18 °C, protein expression was induced by the addition of 0.5 mM IPTG and conducted overnight at 18 °C. Harvested bacterial cells were frozen and stored at −80 °C until further use.
Protein purification
USP53 and USP54 were purified by Ni-NTA affinity purification, reverse Ni-NTA affinity purification, anion-exchange chromatography and SEC. Bacterial cells were resuspended in buffer N (20 mM NaH2PO4 pH 8.0, 300 mM NaCl, 20 mM imidazole, 4 mM β-mercaptoethanol (β-ME) supplemented with DNAseI, lysozyme, EDTA-free protease inhibitor cocktail (one tablet in 100 ml, Roche) and 1 mM PMSF) and lysed by sonication (55% amplitude; 5 s on, 10 s off) on ice. After pelleting, the lysate was loaded onto a Ni-NTA FF column (Cytiva) and eluted with buffer N containing 500 mM imidazole. Protein-containing fractions were pooled, His-3C protease was added and the samples were dialyzed overnight against buffer N. After reverse Ni-NTA, the flowthrough was dialyzed overnight against LS buffer (25 mM Tris pH 8.5, 50 mM NaCl and 4 mM β-ME) and subjected to anion-exchange chromatography using a Resource Q (ResQ) column (GE Healthcare) with an elution gradient ranging from 50 to 500 mM NaCl in LS buffer. Peaks containing the protein of interest were pooled, concentrated and either subjected to SEC using buffer S (20 mM Tris pH 8.0, 100 mM NaCl and 4 mM β-ME) on a HiLoad 16/600 Superdex 75pg column (GE Healthcare) or buffer-exchanged into buffer S.
Initially, a USP54 construct spanning amino acids 21–385 was expressed, which was found to be C-terminally truncated during purification. The mass of the main species matched to amino acids 21–369. A corresponding construct was generated and used for all subsequent experiments as indicated.
Purification of probe-labeled USP54 was performed similar to apo USP54. After Ni-NTA and reverse Ni-NTA purification, the protein solution was buffer-exchanged into 20 mM Tris pH 8.0, 100 mM NaCl and 5 mM DTT with the probe added in a 1:3 molar ratio and incubated overnight at 4 °C. A pure complex of USP54 conjugated to diubiquitin–PA was obtained from anion-exchange chromatography and SEC, which were performed as described for unlabeled USP54 (Extended Data Fig. 5).
OtUBD and Avi-tagged TUBE reagents were purified by Ni-NTA and SEC as described above with the adjustments of 50 mM NaH2PO4 in buffer N, a different SEC buffer (described below) and no protease inhibitor cocktail.
GFP-C-LACE-tag, UBE2N and UBE2S were purified by Ni-NTA purification, reverse Ni-NTA purification and SEC as described above for USP53. Purification protocols for GST fusion proteins of UBE2R, UBE2V, UBE2S-UBD, UBE2L/UBCH7 and NleL68,69, as well as for AMSH* and OTUB1*70, were adopted from the literature. mE1 was purified by Ni-NTA, ResQ and SEC. His-3C protease was obtained from Dortmund Protein Facility.
Ubiquitin, ubiquitin variants and M1-linked ubiquitin chains were purified by acidic precipitation, cation-exchange chromatography and SEC71. Cell pellets were resuspended in 20 mM Tris pH 7.4 and 2 mM EDTA and lysed by sonication (55% amplitude; 10 s on, 10 s off) on ice. The cleared lysate was mixed with 0.5% (v/v) perchloric acid on ice to induce protein precipitation. The remaining soluble protein was dialyzed against buffer F (50 mM sodium acetate pH 4.5) and subjected to cation-exchange chromatography using a HiPrep SP FF 16/10 column (GE Healthcare) with an elution gradient from 0 to 1,000 mM NaCl in buffer F. Fractions containing ubiquitin were further purified on a HiLoad 16/600 Superdex 75pg column in PBS.
Protein concentrations were determined by absorption measurements at 280 nm (Nanodrop) using calculated extinction coefficients and molecular weights. Aliquoted protein samples were snap-frozen in liquid nitrogen and stored at −80 °C until further use.
Generation of ubiquitin-based and Ubl-based probes
Ubiquitin and Ubl probes with PA and VS warheads were prepared as described previously through direct conversion of C-terminal protein thioesters with small-molecule amines41,72.
Enzymatic assembly of linkage-specific ubiquitin chains
K6-linked, K11-linked and K48-linked tetraubiquitin chains and K63-linked diubiquitin, triubiquitin and tetraubiquitin chains were assembled enzymatically from ubiquitin as described previously66. Reactions were performed at 37 °C, monitored by intact protein mass spectrometry and stopped with 5 mM DTT. Typical reaction times ranged from 1 to 8 h. For purification, reaction mixtures were diluted in buffer F and subjected to cation-exchange chromatography using a Resource S (ResS) column with an elution gradient from 0 to 1,000 mM NaCl in buffer F and SEC on a HiLoad 16/600 Superdex 75pg column in PBS. Enzymatically assembled K29-linked and K33-linked tetraubiquitin chains were purchased from Bio-Techne (UC-83-025 and UC-103-025).
Enzymatic assembly reactions were also used for the generation of ubiquitin chains containing amino acid changes. Details are provided below.
Generation of fluorescent TAMRA–maleimide-labeled, K63-linked ubiquitin chains
Ubiquitin–KGTAMRA was obtained as reported previously72. Introduction of a C-terminal cysteine and alanine to a shortened ubiquitin (Ub1–75-CA) enabled fluorescent labeling of the proximal ubiquitin in polyubiquitin chains by TAMRA–maleimide as follows: K63-linked diubiquitin chains containing Ub1–75-CA as proximal ubiquitin (UbK63R–Ub1–75-CA) were enzymatically assembled with 900 µM UbK63R and 1.3 mM Ub1–75-CA for 3 h at 37 °C in K63 assembly buffer (40 mM Tris pH 8.5, 10 mM MgCl2, 1 µM E1, 8 µM UbEV1, 8 µM UbE2N and 10 mM adenosine triphosohate). K63-linked triubiquitin chains containing Ub1–75-CA as proximal ubiquitin (Ub2–Ub1–75-CA) were obtained by using 180 µM K63-linked diubiquitin and 1,800 µM Ub1–75-CA in K63 assembly buffer to favor formation of triubiquitin over tetraubiquitin. Reactions were incubated at 37 °C until diubiquitin was consumed (monitored by protein mass spectrometry). To obtain K63-linked tetraubiquitin chains containing Ub1–75-CA as proximal ubiquitin (Ub3–Ub1–75-CA), 80 µM Ub2–Ub1–75-CA was combined with 500 µM UbK63R in K63 assembly buffer and incubated for 2 h at 37 °C.
The resulting substrates were purified on a ResS column with an elution gradient from 0 to 1,000 mM NaCl in buffer F (50 mM sodium acetate pH 4.5). Next, UbK63R–Ub1–75-CA, Ub2–Ub1–75-CA and UbK63R–Ub2–Ub1–75-CA were buffer-exchanged into labeling buffer (1× PBS and 1 mM TCEP) and fluorescently labeled by the addition of a 1.1 molar equivalent of TAMRA–maleimide. After 1-h incubation at room temperature (RT), the reaction was stopped with 5 mM DTT. Excess dye was removed by dialysis and SEC on a HiLoad 16/600 Superdex 75pg column in PBS. Samples were protected from light from the labeling reaction onward.
Generation of K63-linked diubiquitin–PA
K63-linked diubiquitin–PA was obtained by mixing 750 µM ubiquitin–PA with 800 µM UbK63R for 16 h at RT in K63 assembly buffer. Purification on a ResS column with an elution gradient from 0 to 1,000 mM NaCl in buffer F and on a HiLoad 16/600 Superdex 75pg column in buffer A yielded pure K63-linked diubiquitin–PA.
Crystallization
Crystallization screens were set up by a mosquito HTS robot (TTP Labtech) in 96-well sitting drop vapor diffusion plates of Medical Research Council (MRC) format (Molecular Dimensions) and were incubated at 20 °C. Commercially available plates were used for coarse screening, whereas reservoir solutions of fine-screening plates were prepared with a Dragonfly robot (TTP Labtech). The crystal used for structure determination was grown in a 500 + 500-nl drop prepared from 13 mg ml−1 USP54 conjugated to K63-linked diubiquitin–PA in 20 mM Tris pH 8.0, 100 mM NaCl and 4 mM DTT and a reservoir containing 100 mM HEPES pH 6.8, 27.2% (v/v) PEG400 and 200 mM CaCl2. Cryoprotection was achieved by soaking in 100 mM HEPES pH 7.3, 30% (v/v) PEG400 and 200 mM CaCl2 followed by vitrification in liquid nitrogen.
Data collection, structure solution and refinement
Diffraction data for crystals of USP54 conjugated to K63-linked diubiquitin–PA were collected at 100 K at the Swiss Light Source (SLS, Paul Scherrer Institute) on beamline PX2. Datasets were integrated using DIALS73 and scaled using AIMLESS74. Positions and identities of the zinc atoms and all protein chains were obtained from an anomalous dataset (collected at a wavelength of 1.2823 Å) using the CRANK2 pipeline75. The structure was solved by molecular replacement from a native dataset (collected at a wavelength of 1.000 Å) using MR Phaser76 with a truncated AlphaFold model of USP54 (AF-Q70EL1-F1) and the structure of free ubiquitin (Protein Data Bank (PDB) 1UBQ) as search models, whose relative orientations were obtained from the CRANK2 output. The final model was obtained by multiple cycles of structure building in Coot77 and refinement with phenix.refine78. Data collection and refinement statistics are shown in Supplementary Table 2. Data were deposited to the PDB under accession code 8C61. Interface characteristics were determined from the Proteins, Interfaces, Structures and Assemblies webserver (https://www.ebi.ac.uk/pdbe/pisa/).
SEC–MALS
SEC–MALS measurements were performed on an Agilent Infinity II high-performance liquid chromatography (HPLC) system coupled to a Wyatt DAWN NEON 8 angle detector, an Optilab online refractive index detector and an Agilent VWD ultraviolet detector. Protein solutions (60 µl at 2 mg ml−1) were injected and separated on a Superdex 75 Increase 10/300 column (Cytiva) with a flow of 0.5 ml min−1 in 20 mM Tris pH 8.0 and 100 mM NaCl. Masses were determined with the program Astra (Wyatt Technology) using BSA for calibration.
Analytical protein labeling
DUBs and probes were mixed in a 1:3 ratio in buffer A (20 mM Tris pH 7.7, 100 mM NaCl and 5 mM DTT) (Extended Data Fig. 2e,f) or buffer B (1× PBS and 5 mM DTT) (Fig. 1d and Extended Data Fig. 1c) to final concentrations of 2–3.5 µM DUB and 6–10.5 µM probe, followed by incubation for 1 h at RT (Extended Data Fig. 2e,f) or 37 °C (Fig. 1d and Extended Data Fig. 1c).
Ubiquitin–RhoG and Ubl–RhoG cleavage assay
For DUB activity assays using fluorogenic ubiquitin–RhoG and Ubl–RhoG substrates, solutions of 2× DUB (final concentrations: 2 nM–4 µM) with 2× ubiquitin–RhoG and Ubl–RhoG (final concentration: 50 nM) were prepared in 20 mM Tris pH 7.7, 100 mM NaCl, 5 mM DTT and 0.05 mg ml−1 BSA. Then, 10 µl of 2× DUB solution were added in triplicate to black 384-well low-volume nonbinding surface plates (Greiner). Reactions were started by the addition of 10 µl of 2× ubiquitin and Ubl RhoG to each well. Substrate cleavage was monitored by fluorescence measurement of liberated RhoG (excitation: 492 nm, emission: 525 nm) on a TECAN Spark plate reader for 1 h at 25–30 °C.
Fluorescence polarization activity assay
The cleavage of fluorescent ubiquitin chains of different lengths was monitored by fluorescence polarization measurements in black 384-well low-volume nonbinding surface plates (Greiner) on a TECAN Spark plate reader for 2 h at 25 °C (excitation: 535 nm, emission: 595 nm). Substrate and enzyme solutions were prepared separately in FP buffer (20 mM Tris pH 7.7, 100 mM NaCl, 5 mM DTT and 0.1 mg ml−1 BSA). Then, 10 µl of 2× substrate solution (final concentration: 50 nM) was added to the plates and preincubated for 10 min. Reactions were started by addition of 10 µl of 2× enzyme solution (final concentrations: 0.0625–4 µM) to each well. Observed rate constants (kobs) were calculated from averaged technical triplicates using GraphPad Prism and the function ‘plateau followed by one phase decay’. Catalytic efficiencies were obtained as the slope from linear fitting of kobs plotted against enzyme concentration.
Gel-based ubiquitin chain cleavage assay
Ubiquitin chains as 2× substrate dilutions (final concentration: 2–4 µM) and 2× enzyme solutions (final concentration: 0.3–3 µM) were prepared in buffer A (Figs. 2b,d, 5g and 6d,e and Extended Data Fig. 3a,c) or buffer B (Figs. 1f and 2e and Extended Data Figs. 3e,g and 6c) and equilibrated to reaction temperature. Reactions were started by mixing substrate and enzyme solutions in a 1:1 ratio, followed by incubation at 37 °C for most experiments and at RT for experiments shown in Figs. 2e and 5g and Extended Data Fig. 3c,e. Samples were taken at the indicated time points and substrate cleavage was analyzed by SDS–PAGE on NuPAGE Bis–Tris gradient gels (4–12%, Invitrogen) and Coomassie staining. Cleavage of fluorescent TAMRA-labeled ubiquitin chains was additionally visualized by in-gel fluorescence using the Alexa546 channel on a Chemidoc MP Imaging System (Bio-Rad). Protein levels were quantified by densitometry with the program ImageJ. For quantitative evaluation of DUB cleavage activity, the intensities for remaining substrate levels were normalized to the intensities for initial substrate levels. Averaged ratio and s.d. values were calculated from 3–4 independent cleavage assays.
GFP–ubiquitin cleavage assay
GFP-tagged diubiquitin, triubiquitin and tetraubiquitin substrates with K63 or K48 linkages were assembled from ubiquitinated, LACE-tagged GFP44. Purified GFP–ubiquitin substrates (final concentration: 1 µM) and enzyme solutions (final concentration: 0.3 µM for USP54 and 0.5 µM for USP53) were prepared in 20 mM Tris pH 7.7, 100 mM NaCl and 5 mM DTT as 2× dilutions. Solutions were preheated to the reaction temperature of 37 °C and mixed in a 1:1 ratio. Samples were taken at indicated time points and subsequently analyzed by intact protein mass spectrometry or SDS–PAGE. Protein bands on gels were visualized by in-gel fluorescence using the Alexa 488 channel and subsequent Coomassie staining.
Intact protein mass spectrometry
Protein samples were diluted to ~0.5–1 mg ml−1 in PBS and desalted on an AdvanceBio desalting reverse-phase cartridge (Agilent) on an Agilent 1260 II Infinity system before analysis on a mass-selective detection mass analyzer as described previously41. Spectral deconvolution was performed with the program ProMass (Enovatia).
Protein stability measurements
The thermal stability of apo and probe-labeled proteins was determined by a TSA. Samples were diluted in TSA assay buffer (20 mM HEPES pH 7.5, 100 mM NaCl and 5 mM DTT) and probe labeling was conducted with a 3× excess of probe overnight at 4 °C. From these, 2× protein solutions (6 µM) were prepared and mixed with SYPRO Orange dye (6×, in TSA assay buffer) in a 1:1 ratio. USP54F161K-containing proteins were purified before analysis and were assayed in a PBS–Tris pH 8.0 buffer mixture. Samples were added to white 96-well PCR plates (Bio-Rad) and a 20–90 °C gradient was applied (increment: 0.3 °C, held for 5 s before read) during which the fluorescence intensity was monitored with a CFX connect real-time PCR system (Bio-Rad) using the fluorescence resonance energy transfer channel (excitation: 450–490 nm, emission: 560–580 nm). Data were analyzed with CFX Maestro.
Lentivirus generation
Lenti-X 293T cells were transfected using PEI and a 4:3:2 ratio of Tet-pLKO-puro-shRNA:pPAX2:pMD2.G plasmids in Opti-MEM medium and the medium was exchanged on the next day. Viral supernatant was then collected 48 h, 72 h and 96 h after transfection. The pooled supernatant was centrifuged for 5 min and subsequently filtered. The filtered supernatant was concentrated using concentrating filter units (Amicon Ultra-15, 100-kDa cutoff, Sigma-Aldrich) for 30 min to 1 h at 4,000g and stored at −80 °C.
Cell culture, transfection and transduction
HeLa cells were obtained from the Leibniz Institute DSMZ (German Collection of Microorganisms and Cell Cultures). Human embryonic kidney Lenti-X 293T cells were obtained from Takara Bio and CaCo-2 cells were obtained from the American Type Culture Collection. Cells were cultivated in a humidified incubator at 37 °C and 5% CO2 in DMEM supplemented with either 10% (HeLa and Lenti-X 293 T) or 20% (CaCo-2) FBS and penicillin–streptomycin. Cells tested negative for Mycoplasma. For the creation of doxycycline-inducible knockdown cell lines, the medium was changed to fresh culture medium containing 8 μg ml−1 polybrene (Sigma-Aldrich). Concentrated lentivirus was added dropwise. Then, 24 h after infection, virus-containing medium was exchanged to fresh medium. Cell selection was started 48 h after infection and carried out by gradually increasing puromycin (12–16 μg ml−1) over the course of 10 days. Knockdown was confirmed by western blot after treatment with 2 μg ml−1 doxycycline for 72 h and medium exchange every 24 h. Cells were harvested with RIPA buffer (50 mM Tris HCl pH 8, 150 mM NaCl, 1% IGEPAL CA-630, 0.5% sodium deoxycholate and 0.1% SDS).
Mass spectrometric analysis of active DUBs
Identification of HA–ubiquitin–PA probe-labeled proteins by mass spectrometry was carried out as described previously41. In short, HeLa cell lysate was incubated with HA–ubiquitin–PA or with HA–UFM1–PA as control. Probe-labeled proteins were enriched by HA pulldown and subsequently identified by mass spectrometry.
Western blotting
Proteins were transferred to a nitrocellulose membrane using a Trans-Blot Turbo system (Bio-Rad) at 1.0 A, 25 V, 30 min for ubiquitin chain analysis or 1.3 A, 25 V, 10 min for all other blots. Membranes were blocked with 5% (w/v) nonfat milk in PBS-T buffer and incubated with indicated primary antibodies (anti-USP53, 1:1,000, Sigma-Aldrich, HPA035844; anti-MARVELD2, 1:500, Thermo Fisher Scientific, 700191; anti-tubulin, 1:4,000, Sigma-Aldrich, T6199; anti-GAPDH, 1:10,000, Thermo Fisher Scientific, AM4300) overnight at 4 °C. Membranes were then incubated with the respective secondary antibody (anti-mouse, 1:5,000, Sigma-Aldrich, NXA931; anti-rabbit, 1:5,000, Sigma-Aldrich, GENA934) coupled to horseradish peroxidase. The chemiluminescent reaction was initiated using a Clarity Western enhanced chemiluminescence substrate and images were taken on a ChemiDoc MP Imaging System (Bio-Rad).
Ubiquitinome sample preparation
CaCo-2 cells stably transduced with USP53 shRNA5 were seeded in eight 10-cm dishes, with each dish giving rise to one analyzed sample. Knockdown of USP53 was induced in four dishes using 2 μg ml−1 doxycycline 24 h after seeding and ultrapure water was used as the control in the other four dishes. The medium was changed every 24 h to medium with the respective treatment solution. Then, 72 h after induction of the knockdown, cells were washed thrice in ice-cold PBS and lysed in lysis buffer (50 mM Tris HCl pH 8.5 and 1% (w/v) sodium deoxycholate). Cells were scraped off the dish and proteins were denatured at 95 °C for 10 min. Cell lysates were snap-frozen in liquid nitrogen. Cells were lysed further using sonication in a Bioruptor Pico (Diagenode). Protein concentrations were measured using the BCA assay (Thermo Fisher Scientific). Diglycine-modified peptides were enriched by immunoprecipitation using the PTMScan ubiquitin remnant motif (K-ε-GG) antibody–bead conjugate (kit 5562, Cell Signaling Technology) starting from 800 µg of total protein according to the manufacturer’s protocol. Unbound peptides were removed by washing and the captured peptides were eluted with a low-pH buffer. Eluted peptides were analyzed by nanoflow LC–MS/MS.
Nanoflow LC–MS/MS
Mass spectrometry was performed on a Vanquish Neo ultra-HPLC System (Thermo Fisher Scientific) coupled to an Exploris 480 Orbitrap mass spectrometer (Thermo Fisher Scientific), operating in positive mode and equipped with a nanospray source. Peptide mixtures were separated on a CSH130 2.5-µm reverse-phase column (Waters; column dimensions 30 cm × 75 µm, packed in-house) using a linear gradient from 0% to 40% B (A = 0.1% formic acid in water; B = 80 % (v/v) acetonitrile in water, with 0.1% formic acid) in 80 min and at a constant flow rate of 300 nl min−1. The column eluent was directly sprayed into the electrospray ionization source of the mass spectrometer. All spectra were recorded in DIA mode at a resolution of 120,000 for full scans in the scan range from 350 to 1,650 m/z. The maximum injection time was set to 50 ms (automatic gain control (AGC) target: 4 × 105). For MS2 acquisition, the mass range was set to 336–1,391 m/z with dynamic isolation windows ranging from 7 to 82 m/z with a window overlap of 1 m/z. The orbitrap resolution for MS2 scans was set to 30,000. The maximum injection time was at 54 ms (AGC target: 5 × 104; normalized AGC target: 100%). A single LC–MS/MS run was performed for all immunoprecipitated peptide material from one sample.
Ubiquitinome data analysis
DIA raw data files were analyzed with the Spectronaut Pulsar X software package (version 17.0.221202, www.biognosys.com), using directDIA for DIA analysis including MaxLFQ as the label-free quantitation method and Spectronaut’s IDPicker algorithm for protein inference. The Q-value cutoff at the precursor and protein level was set to 0.01. All imputation of missing values was disabled. Data were further analyzed in Perseus (version 2.0.7.0). Peptide mass spectral intensity values were log2-transformed. Ubiquitinated peptides were filtered for sites that were quantified in all samples and in which the modified lysine was unambiguously identified. Increased ubiquitination sites upon USP53 knockdown were determined with a significance threshold of −log10(P value) ≥ 1 and a fold change threshold of log2(change in abundance) ≥ 1.
Preparation of ubiquitin enrichment reagents
OtUBD with an N-terminal cysteine (111 µM) in SEC buffer (20 mM Tris, 100 mM NaCl, 5 mM EDTA and 4 mM TCEP, pH 8.5) was incubated after fresh addition of TCEP (one equivalent) for 30 min. Biotin–PEG2–iodoacetamide (ten equivalents) was then added and the reaction mixture was incubated at 22 °C for 4 h. The protein was purified by a five-step sequential dialysis into storage buffer (20 mM Tris, 100 mM NaCl and 5% glycerol, pH 8). The protein concentration of OtUBD was determined by a DC assay (Bio-Rad). Purified TUBE proteins were biotinylated as described previously79. Protein concentrations of TUBE proteins were determined by a microBCA protein assay (Thermo Fisher Scientific). The identity and purity of all biotinylated proteins were checked by SDS–PAGE and intact protein mass spectrometry. Samples were aliquoted and stored at −80 °C.
Enrichment of endogenously ubiquitinated proteins
CaCo-2 cells (stably transduced with USP53 shRNA where indicated) were treated as described above for the ubiquitinome analysis. Then, 72 h after induction of the knockdown, cells were washed twice with ice-cold PBS and lysed in urea lysis buffer (8 M urea, 50 mM Tris HCl pH 8, 150 mM NaCl, 1% IGEPAL, 2 mM EDTA, 5% glycerol, 1× EDTA-free protease inhibitor cocktail, 2 µM MG132, 1 mM PMSF, 10 µM PR619, 20 mM N-ethylmaleimide and 4 mM 1,10-phenanthroline) and harvested by scraping. Directly after lysis, the obtained suspension was diluted 1:1 for OtUBD and 1:3 for pan-polyubiquitin and K63-linked polyubiquitin enrichment experiments with dilution buffer (urea lysis buffer devoid of urea). Cells were homogenized using sonication for 12 s (3 s on and 3 s off) at 10% amplitude and centrifuged for 10 min at 14,000g and 4 °C.
In parallel, 17 nmol of the biotinylated ubiquitin-binding entities were immobilized on 240 µl of high-capacity neutravidin agarose bead slurry (Pierce, Thermo Fisher Scientific) for 1 h at 4 °C with rotation. Excess reagent was washed away with ice-cold PBS. The beads were then equilibrated in diluted lysis buffer before being split up in two tubes.
For each condition, an equal amount of 8–10 mg of protein (as determined per Bradford assay, in 2 ml of diluted lysis buffer, derived from two confluent 10-cm dishes per condition) was added to the preimmobilized pulldown reagents and incubated for 2 h at 4 °C with rotation. Beads were pelleted by centrifugation at 500g for 1 min and the supernatant was removed. The resin was washed once with diluted lysis buffer, once with high-salt buffer (50 mM Tris HCl pH 8 and 1 M NaCl) and once with ice-cold PBS. The pulldown reagent and bound proteins were eluted using 2× LDS sample buffer with 25 mM DTT and boiling for 5 min at 95 °C. After pelleting of the beads at 500g for 1 min, the supernatant was transferred and analyzed by SDS–PAGE and western blotting as described above.
UbiCRest assays
Ubiquitinated proteins were isolated as described above using the OtUBD pulldown reagent. After the abovementioned washing steps, beads were additionally washed twice with water. Subsequently, ubiquitinated proteins were eluted from OtUBD using 50 µl of a 100 mM glycine solution at pH 2.5 with an incubation time of 5 min at RT. Beads were pelleted by centrifugation at 500g for 1 min. The supernatant was transferred to a new tube and brought immediately to a neutral pH with 5 µl of 1 M Tris at pH 9.0. This step was repeated once; the resulting supernatants were combined and supplemented with final concentrations of 5 mM DTT, 5 mM MgCl2 and 100 mM NaCl.
For UbiCRest assays, supernatants were incubated with USP2 (1 µM), OTUB1* (0.2 µM) or AMSH* (0.2 µM) for 1 h at 37 °C. The reactions were stopped by adding 4× LDS sample buffer (supplemented with 50 mM DTT) and visualized by SDS–PAGE and western blotting as described above.
To assess the activity of the catalytic domains of USP53 or USP54 on ubiquitinated proteins, enzymes were added to the supernatants at final concentrations of 0.5, 2 and 5 µM or 0.3, 1 and 3 µM, respectively. Samples were incubated for 1–2 h at 37 °C and analyzed as described above.
Statistics and reproducibility
Results were consistently observed in at least two, typically three independent experiments. For the statistical testing of quantitative data for the ubiquitin probe pulldown (Fig. 1a) and diglycine proteomics (Fig. 3e), two-sided t-tests were used as implemented in Perseus using 250 randomizations, a false discovery rate of 0.05 and an s0 of 0.1.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Coordinates and structure factors for the crystal structure of USP54 conjugated to K63-linked diubiquitin–PA were deposited to the PDB under accession code 8C61. Mass spectrometry raw data were deposited to the ProteomeXchange Consortium through the PRIDE partner repository under accession codes PXD038455 and PXD054748. Coordinates of other structures were obtained from the PDB under accession codes 1NBF, 1UBQ, 2AYO, 2IBI, 2ZNV, 3WXF, 3WXG, 3ZNZ, 4BOS, 5E6J, 5L8W, 6FGE, 6UPU and 7NPI. Source data are provided with this paper.
References
Komander, D. & Rape, M. The ubiquitin code. Annu. Rev. Biochem. 81, 203–229 (2012).
Dikic, I. & Schulman, B. A.An expanded lexicon for the ubiquitin code. Nat. Rev. Mol. Cell Biol. 24, 273–287 (2023).
Kim, W. et al. Systematic and quantitative assessment of the ubiquitin-modified proteome. Mol. Cell 44, 325–340 (2011).
Mukhopadhyay, D. & Riezman, H. Proteasome-independent functions of ubiquitin in endocytosis and signaling. Science 315, 201–205 (2007).
Pruneda, J. N. et al. The molecular basis for ubiquitin and ubiquitin-like specificities in bacterial effector proteases. Mol. Cell 63, 261–276 (2016).
Haahr, P. et al. ZUFSP deubiquitylates K63-linked polyubiquitin chains to promote genome stability. Mol. Cell 70, 165–174 (2018).
Tsuchiya, H. et al. Ub-ProT reveals global length and composition of protein ubiquitylation in cells. Nat. Commun. 9, 524 (2018).
Lutz, J., Hollmuller, E., Scheffner, M., Marx, A. & Stengel, F. The length of a ubiquitin chain: a general factor for selective recognition by ubiquitin-binding proteins. Angew. Chem. Int. Ed. Engl. 59, 12371–12375 (2020).
Lu, Y., Lee, B. H., King, R. W., Finley, D. & Kirschner, M. W. Substrate degradation by the proteasome: a single-molecule kinetic analysis. Science 348, 1250834 (2015).
Ikeda, F., Crosetto, N. & Dikic, I. What determines the specificity and outcomes of ubiquitin signaling? Cell 143, 677–681 (2010).
Kwasna, D. et al. Discovery and characterization of ZUFSP/ZUP1, a distinct deubiquitinase class important for genome stability. Mol. Cell 70, 150–164 (2018).
Waltho, A. et al. K48- and K63-linked ubiquitin chain interactome reveals branch- and length-specific ubiquitin interactors. Life Sci. Alliance 7, e202402740 (2024).
Swatek, K. N. & Komander, D. Ubiquitin modifications. Cell Res. 26, 399–422 (2016).
Abdul Rehman, S. A. et al. Mechanism of activation and regulation of deubiquitinase activity in MINDY1 and MINDY2. Mol. Cell 81, 4176–4190 (2021).
Mevissen, T. E. et al. OTU deubiquitinases reveal mechanisms of linkage specificity and enable ubiquitin chain restriction analysis. Cell 154, 169–184 (2013).
Bekes, M. et al. Recognition of Lys48-linked di-ubiquitin and deubiquitinating activities of the SARS coronavirus papain-like protease. Mol. Cell 62, 572–585 (2016).
Shin, D. et al. Papain-like protease regulates SARS-CoV-2 viral spread and innate immunity. Nature 587, 657–662 (2020).
Beck, D. B. et al. Linkage-specific deubiquitylation by OTUD5 defines an embryonic pathway intolerant to genomic variation. Sci. Adv. 7, eabe2116 (2021).
Damgaard, R. B. et al. The deubiquitinase OTULIN is an essential negative regulator of inflammation and autoimmunity. Cell 166, 1215–1230 (2016).
Clague, M. J., Urbe, S. & Komander, D. Breaking the chains: deubiquitylating enzyme specificity begets function. Nat. Rev. Mol. Cell Biol. 20, 338–352 (2019).
Tsou, W. L. et al. Systematic analysis of the physiological importance of deubiquitinating enzymes. PLoS ONE 7, e43112 (2012).
Abdul Rehman, S. A. et al. MINDY-1 is a member of an evolutionarily conserved and structurally distinct new family of deubiquitinating enzymes. Mol. Cell 63, 146–155 (2016).
Hewings, D. S. et al. Reactive-site-centric chemoproteomics identifies a distinct class of deubiquitinase enzymes. Nat. Commun. 9, 1162 (2018).
Faesen, A. C. et al. The differential modulation of USP activity by internal regulatory domains, interactors and eight ubiquitin chain types. Chem. Biol. 18, 1550–1561 (2011).
Hermanns, T. et al. A family of unconventional deubiquitinases with modular chain specificity determinants. Nat. Commun. 9, 799 (2018).
Sato, Y. et al. Structural basis for specific cleavage of Lys 63-linked polyubiquitin chains. Nature 455, 358–362 (2008).
Walden, M. et al. Metabolic control of BRISC–SHMT2 assembly regulates immune signalling. Nature 570, 194–199 (2019).
Rabl, J. et al. Structural basis of BRCC36 function in DNA repair and immune regulation. Mol. Cell 75, 483–497 (2019).
Mevissen, T. E. T. & Komander, D. Mechanisms of deubiquitinase specificity and regulation. Annu. Rev. Biochem. 86, 159–192 (2017).
Kazmierczak, M. et al. Progressive hearing loss in mice carrying a mutation in Usp53. J. Neurosci. 35, 15582–15598 (2015).
Fraile, J. M., Campos-Iglesias, D., Rodriguez, F., Espanol, Y. & Freije, J. M. The deubiquitinase USP54 is overexpressed in colorectal cancer stem cells and promotes intestinal tumorigenesis. Oncotarget 7, 74427–74434 (2016).
Ye, Y., Scheel, H., Hofmann, K. & Komander, D. Dissection of USP catalytic domains reveals five common insertion points. Mol. Biosyst. 5, 1797–1808 (2009).
Quesada, V. et al. Cloning and enzymatic analysis of 22 novel human ubiquitin-specific proteases. Biochem. Biophys. Res. Commun. 314, 54–62 (2004).
Alhebbi, H. et al. New paradigms of USP53 disease: normal GGT cholestasis, BRIC, cholangiopathy, and responsiveness to rifampicin. J. Hum. Genet 66, 151–159 (2021).
Bull, L. N. et al. Cholestasis due to USP53 deficiency. J. Pediatr. Gastroenterol. Nutr. 72, 667–673 (2021).
Maddirevula, S. et al. Identification of novel loci for pediatric cholestatic liver disease defined by KIF12, PPM1F, USP53, LSR, and WDR83OS pathogenic variants. Genet. Med. 21, 1164–1172 (2019).
Zhang, J. et al. Low-GGT intrahepatic cholestasis associated with biallelic USP53 variants: clinical, histological and ultrastructural characterization. Liver Int. 40, 1142–1150 (2020).
Pinto-Fernandez, A. et al. Comprehensive landscape of active deubiquitinating enzymes profiled by advanced chemoproteomics. Front. Chem. 7, 592 (2019).
Borodovsky, A. et al. A novel active site-directed probe specific for deubiquitylating enzymes reveals proteasome association of USP14. EMBO J. 20, 5187–5196 (2001).
Ekkebus, R. et al. On terminal alkynes that can react with active-site cysteine nucleophiles in proteases. J. Am. Chem. Soc. 135, 2867–2870 (2013).
O’Dea, R. et al. Molecular basis for ubiquitin/Fubi cross-reactivity in USP16 and USP36. Nat. Chem. Biol. 19, 1394–1405 (2023).
Sato, Y. et al. Structures of CYLD USP with Met1- or Lys63-linked diubiquitin reveal mechanisms for dual specificity. Nat. Struct. Mol. Biol. 22, 222–229 (2015).
Ritorto, M. S. et al. Screening of DUB activity and specificity by MALDI-TOF mass spectrometry. Nat. Commun. 5, 4763 (2014).
Hofmann, R., Akimoto, G., Wucherpfennig, T. G., Zeymer, C. & Bode, J. W. Lysine acylation using conjugating enzymes for site-specific modification and ubiquitination of recombinant proteins. Nat. Chem. 12, 1008–1015 (2020).
Bekes, M. et al. SARS hCoV papain-like protease is a unique Lys48 linkage-specific di-distributive deubiquitinating enzyme. Biochem. J. 468, 215–226 (2015).
Lee, B. H. et al. USP14 deubiquitinates proteasome-bound substrates that are ubiquitinated at multiple sites. Nature 532, 398–401 (2016).
Riazuddin, S. et al. Tricellulin is a tight-junction protein necessary for hearing. Am. J. Hum. Genet 79, 1040–1051 (2006).
Ikenouchi, J. et al. Tricellulin constitutes a novel barrier at tricellular contacts of epithelial cells. J. Cell Biol. 171, 939–945 (2005).
Masuda, S. et al. LSR defines cell corners for tricellular tight junction formation in epithelial cells. J. Cell Sci. 124, 548–555 (2011).
Kamitani, T. et al. Deletion of tricellulin causes progressive hearing loss associated with degeneration of cochlear hair cells. Sci. Rep. 5, 18402 (2015).
Higashi, T. et al. Analysis of the ‘angulin’ proteins LSR, ILDR1 and ILDR2—tricellulin recruitment, epithelial barrier function and implication in deafness pathogenesis. J. Cell Sci. 126, 966–977 (2013).
Zhang, M., Berk, J. M., Mehrtash, A. B., Kanyo, J. & Hochstrasser, M. A versatile new tool derived from a bacterial deubiquitylase to detect and purify ubiquitylated substrates and their interacting proteins. PLoS Biol. 20, e3001501 (2022).
Berk, J. M. et al. A deubiquitylase with an unusually high-affinity ubiquitin-binding ___domain from the scrub typhus pathogen Orientia tsutsugamushi. Nat. Commun. 11, 2343 (2020).
Hjerpe, R. et al. Efficient protection and isolation of ubiquitylated proteins using tandem ubiquitin-binding entities. EMBO Rep. 10, 1250–1258 (2009).
Sims, J. J. et al. Polyubiquitin-sensor proteins reveal localization and linkage-type dependence of cellular ubiquitin signaling. Nat. Methods 9, 303–309 (2012).
Flierman, D. et al. Non-hydrolyzable diubiquitin probes reveal linkage-specific reactivity of deubiquitylating enzymes mediated by S2 pockets. Cell Chem. Biol. 23, 472–482 (2016).
Samara, N. L. et al. Structural insights into the assembly and function of the SAGA deubiquitinating module. Science 328, 1025–1029 (2010).
Kohler, A., Zimmerman, E., Schneider, M., Hurt, E. & Zheng, N. Structural basis for assembly and activation of the heterotetrameric SAGA histone H2B deubiquitinase module. Cell 141, 606–617 (2010).
Hu, M. et al. Crystal structure of a UBP-family deubiquitinating enzyme in isolation and in complex with ubiquitin aldehyde. Cell 111, 1041–1054 (2002).
Klemm, T. et al. Mechanism and inhibition of the papain-like protease, PLpro, of SARS-CoV-2. EMBO J. 39, e106275 (2020).
Jonsson, E., Htet, Z. M., Bard, J. A. M., Dong, K. C. & Martin, A. Ubiquitin modulates 26S proteasome conformational dynamics and promotes substrate degradation. Sci. Adv. 8, eadd9520 (2022).
Reyes-Turcu, F. E. et al. The ubiquitin binding ___domain ZnF UBP recognizes the C-terminal diglycine motif of unanchored ubiquitin. Cell 124, 1197–1208 (2006).
Amendola, M. & Squires, J.E. Pediatric genetic cholestatic liver disease overview. In Gene Reviews (eds Adam, M.P. et al.) (University of Washington, 1993).
Sambrotta, M. et al. Mutations in TJP2 cause progressive cholestatic liver disease. Nat. Genet. 46, 326–328 (2014).
Swatek, K. N. et al. Insights into ubiquitin chain architecture using Ub-clipping. Nature 572, 533–537 (2019).
Michel, M. A., Komander, D. & Elliott, P. R. Enzymatic assembly of ubiquitin chains. Methods Mol. Biol. 1844, 73–84 (2018).
Gersch, M. et al. Distinct USP25 and USP28 oligomerization states regulate deubiquitinating activity. Mol. Cell 74, 436–451 (2019).
Bremm, A., Freund, S. M. & Komander, D. Lys11-linked ubiquitin chains adopt compact conformations and are preferentially hydrolyzed by the deubiquitinase Cezanne. Nat. Struct. Mol. Biol. 17, 939–947 (2010).
Hospenthal, M. K., Freund, S. M. & Komander, D. Assembly, analysis and architecture of atypical ubiquitin chains. Nat. Struct. Mol. Biol. 20, 555–565 (2013).
Michel, M. A. et al. Assembly and specific recognition of K29- and K33-linked polyubiquitin. Mol. Cell 58, 95–109 (2015).
Pickart, C. M. & Raasi, S. Controlled synthesis of polyubiquitin chains. Methods Enzymol. 399, 21–36 (2005).
Zhao, Z., O’Dea, R., Wendrich, K., Kazi, N. & Gersch, M. Native semisynthesis of isopeptide-linked substrates for specificity analysis of deubiquitinases and Ubl proteases. J. Am. Chem. Soc. 145, 20801–20812 (2023).
Beilsten-Edmands, J. et al. Scaling diffraction data in the DIALS software package: algorithms and new approaches for multi-crystal scaling. Acta Crystallogr. D Struct. Biol. 76, 385–399 (2020).
Evans, P. R. & Murshudov, G. N. How good are my data and what is the resolution? Acta Crystallogr. D Biol. Crystallogr. 69, 1204–1214 (2013).
Skubak, P. et al. A new MR-SAD algorithm for the automatic building of protein models from low-resolution X-ray data and a poor starting model. IUCrJ 5, 166–171 (2018).
McCoy, A. J. et al. Phaser crystallographic software. J. Appl. Crystallogr. 40, 658–674 (2007).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D Struct. Biol. 66, 486–501 (2010).
Adams, P. D. et al. The PHENIX software for automated determination of macromolecular structures. Methods 55, 94–106 (2011).
Fairhead, M. & Howarth, M. Site-specific biotinylation of purified proteins using BirA. Methods Mol. Biol. 1266, 171–184 (2015).
Acknowledgements
We thank the beamline scientists at the SLS for support during data collection and R. Gasper and P. Geue for support with crystallization and biophysics. We thank all staff at the Max Planck Institute Dortmund for excellent technical support and P. Janning for mass spectrometry support. We are grateful to all members of the M.G. lab for discussions, advice and reagents. We thank L.-M. Kaps, M. Grabow and M. Reichmann for support with biochemical assays and cell culture. We thank T. Sixma and J. Pruneda for comments on the manuscript. We are grateful to D. Komander (MRC Laboratory of Molecular Biology Cambridge and Walter and Eliza Hall Institute of Medical Research Melbourne), D. Alessi (MRC Protein Phosphorylation and Ubiquitylation Unit Dundee), D. Trono (École Polytechnique Fédérale de Lausanne) and D. Wiederschain for the gift of plasmids. Figure 3f, Extended Data Fig. 5b and the graphical abstract were created with BioRender.com. This work was funded by AstraZeneca, Merck KGaA, Pfizer and the Max Planck Society as part of the Chemical Genomics Center III (CGCIII-352S, to M.G.). Work in the M.G. lab is further supported by Deutsche Forschungsgemeinschaft (German Research Foundation; project GE 3110/1-1–Emmy Noether and 424228829–SFB1430, to M.G.) and by the State of North Rhine-Westphalia through the ‘Cancer Targeting Network’ initiative (NW21-062C, to M.G.).
Funding
Open access funding provided by Max Planck Society.
Author information
Authors and Affiliations
Contributions
K.W. and M.G. designed the study. K.W. performed all experiments unless noted otherwise. K.G. performed cellular experiments. K.W., S.R., N.H.K. and J.A.H. performed in vitro assays with mutant proteins. S.P. generated cell lines. S.F. provided ubiquitin enrichment reagents. R.O.D. contributed data in Fig. 1a and Extended Data Fig. 1d. K.B. and J.D. performed ubiquitinome analysis. All authors designed experiments, analyzed data and interpreted results. M.G. supervised the project and wrote the manuscript together with K.W. and with input from all authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Chemical Biology thanks Ivan Dikic, Monique Mulder and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 USP53 and USP54 are active DUBs.
a. Schematic of the workflow used to identify active DUBs. HeLa cell lysate was treated with HA-Ub-PA or HA-UFM1-PA probe. Labelled proteins were enriched and detected by mass spectrometry. b. Schematic of the probe reactivity assay. The C-terminal propargylamine (PA) warhead reacts with the catalytic cysteine of DUBs to a vinyl thioether, forming a covalent DUB~Ub-PA complex. c. Ub/Ubl specificity assayed through probe reactivity with recombinant catalytic domains. A panel of HA-Ub/Ubl-PA probes was incubated with wild-type USP5320-383 or wild-type USP5421-369. Probe reactivity was analyzed by SDS-PAGE and Coomassie staining. Uncropped versions of all gels are provided as Source Data. d. Intact protein mass spectrometry of probes used in panel c. Expected and found masses are given in Dalton. The star denotes an HA-SUMO41–92 species N-terminally truncated by four residues. All other probes have been characterized previously41. e. Schematic of the Ubiquitin-RhodamineG (Ub-RhoG) cleavage assay. DUBs cleave the amide bond between ubiquitin and RhoG and thereby liberate fluorescent RhoG. f. Ub-RhoG cleavage assay. Indicated concentrations of USP5320-383 (left) and USP5421-369 (right) or of the catalytic inactive mutants were added to Ub-RhoG and fluorescence was recorded. Data are shown as average of technical triplicates. CS enzymes denote the use of catalytic cysteine to serine mutated enzymes as in Fig. 1d. Of note, this activity is approximately 3 to 4 orders of magnitude weaker than for other USP DUBs including USP2, USP7, USP36 and USP4241.
Extended Data Fig. 2 Cholestasis-associated patient mutations of USP53 are catalytically impaired.
a. Ub-RhoG cleavage assay. Indicated concentrations of USP5320-383 (WT or patient mutants) were added to Ub-RhoG and fluorescence was recorded over time. Data are shown as average of technical triplicates. b. Ub-RhoG cleavage assay with USP5421-385, shown as in panel a. c-d. Protein stability assessment of USP5320-383 (panel c) or USP5421-369 (panel d) proteins by thermal shift analysis. Averaged fluorescence raw data are shown and melting temperatures (Tm) are plotted from technical triplicates (USP53) or quadruplicates (USP54). e-f. Probe reactivity assay. HA-Ub-VS was incubated with indicated USP5320-383 (panel e) or USP5421-369 (panel f) proteins. Probe reactivity was analyzed by SDS-PAGE and Coomassie staining. Uncropped versions of all gels are provided as Source Data. g. Schematic of the HA-Ub-VS probe reactivity assay.
Extended Data Fig. 3 An S2 site, but not an S3 site, underlies USP53 and USP54 polyubiquitin cleavage.
a. Time-resolved gel-based cleavage assays of K63-linked tetraubiquitin (2 µM) by USP5320-383 (4 µM, left) and USP5421-369 (300 nM, right). Cleavage activity was analyzed by SDS-PAGE and Coomassie staining. Uncropped versions of all gels are provided as Source Data. b. Schematic of expected products of a triubiquitin cleavage assay in which the reagent contains a non-cleavable, C-terminal fluorescent label. Ubiquitin binding sites in a DUB are shown, expected cleavage sites are indicated with arrows. The presence of an S2 site would lead to fluorescently labeled monoubiquitin, whereas the presence of an S2’ site would lead to fluorescently labeled diubiquitin. c. Gel-based cleavage assay with a FlAsH-based reagent. Native K63-linked triubiquitin chains, K63-linked triubiquitin with a C-terminal tetracysteine (TC) tag in the proximal Ubiquitin (Ub3-TC) and FlAsH-labeled triubiquitin (Ub3-TC-FlAsH, all at 3 µM) were incubated with USP5421-369 (300 nM) for indicated times. USP54 activity was impaired by the presence of the FlAsH dye. d. Schematic of the generation of the TAMRA-based, fluorescent K63-linked triubiquitin substrate. TMR, TAMRA. e. Gel-based cleavage assay with a TAMRA-based reagent. Equivalent assays as shown in panel c with K63-linked triubiquitin chains shown in panel d (3 µM substrate, 300 nM enzyme). f. Schematic of the generation of the TAMRA-based, fluorescent K63-linked tetraubiquitin substrate. g. Gel-based cleavage assay. Fluorescent K63-linked tetraubiquitin (3 µM) was incubated with USP5320-383 (4 µM) or USP5421-369 (300 nM). Cleavage activity was analyzed as in Fig. 2e. The equally formed fluorescent diubiquitin and monoubiquitin species demonstrate that neither USP53 nor USP54 possess S3 ubiquitin binding sites. h. Deconvoluted intact protein mass spectra of indicated reagents. Calculated and observed molecular weights are given in Dalton. i. Fluorescence polarization cleavage assays of USP5320-383 and USP5421-369 and indicated reagents. Catalytic efficiencies derived from these data are shown in Fig. 2g.
Extended Data Fig. 4 Validation and specificity analysis of linkage-directed deubiquitination activity in USP53.
a. Intact protein mass spectrometry analysis of GFP-diUb (K63) cleavage, corresponding to Fig. 3a (500 nM USP53, 1 µM substrate). Diubiquitin removed en bloc by USP53 was identified unequivocally. Calculated (c.) and found (f.) protein masses are given in Dalton. b. Cleavage analysis of GFP-triUb (K63) by USP53, shown as in panel a. Triubiquitin removed en bloc during the earlier time points was cleaved further to diubiquitin. En bloc removal was observed in the earliest time point. Cleavage within the chain leading to accumulation of a minor amount of monoubiquitinated substrate was observed at later time points, consistent with Fig. 3a. c. Cleavage assay for ubiquitinated model substrates modified through isopeptide bonds with K48-linked di- or triubiquitin or with K63-linked triubiquitin. GFP-Ubn substrates (1 µM) were incubated with USP5320-383 (500 nM) or USP5421-369 (300 nM). Cleavage activity was analyzed by as shown in Fig. 3a-b. Uncropped versions of all gels and blots are provided as Source Data. d. Schematic of the identified cleavage activities of USP53 and USP54 on GFP modified with K63- or K48-linked triubiquitin chains. e. Overview of ubiquitin pull-down reagents used in this study. Data are derived from the original publications52,54,55. f. Deconvoluted intact protein mass spectra of biotinylated reagents. Calculated and observed molecular weights are given in Dalton. The species in the pan-polyUb TUBE denoted with an asterisk corresponds to a non-covalent acetonitrile adduct. g-h. MARVELD2 ubiquitination analysis through OtUBD pull-downs. Parental CaCo-2 cells (g) or CaCo-2 shRNA1 (USP53) cells (h) were analyzed as described in Fig. 3g. i. Input protein levels for samples used in Fig. 3g and panel j and analyzed as described for Fig. 3i. j. MARVELD2 polyubiquitination analysis with pan-polyubiquitin (4x UBAUBQLN1) TUBE pull-downs. k. UbiCRest assay on OtUBD eluates as in Fig. 3l, but with 2 h incubation.
Extended Data Fig. 5 Assembly and crystallization of a USP54-diUb(K63)-PA complex.
a. Fluorescence raw data of the protein stability assessment. Corresponding melting temperatures are shown in Fig. 4c. b. Assembly and purification strategy to obtain a pure USP54~diUb(K63)-PA complex for crystallization. Of note, cleavage of the native isopeptide bond in the probe was suppressed by protein labeling at low temperature and with an excess of probe over enzyme. c. Deconvoluted intact protein mass spectra of the K63-linked diUb-PA probe, USP54, and the USP54~diUb(K63)-PA complex. Calculated and observed molecular weights are given in Dalton. d. Asymmetric unit (a.s.u.) of the crystal structure of USP54~diUb-PA. All four copies of USP54~diUb-PA are shown in cartoon representation. e. Asymmetric unit (a.s.u.) of the crystal structure of USP54~diUb-PA as in panel d, all four copies are shown in stick representation overlayed with 2FO-FC electron density contoured at 1 σ. f. Chains ABCDEF and chains GHIJKL, each consisting of 2x USP54~diUb-PA, are shown individually and as an overlay in cartoon representation.
Extended Data Fig. 6 Analysis of the USP fold in the USP54~diUb-PA structure.
a. Typical fold of USP DUBs in the structure of USP54~diUb-PA. The palm subdomain is colored red, the thumb subdomain blue and the fingers subdomain green. Additional structural elements including the blocking loops, the switching loop, the Cys loop, the zinc ions, and the catalytic triad are annotated. Close-up views for the coordination of all zinc ions are shown. b. Chains ABC, DEF, GHI and JKL of the crystal structure of USP54~diUb-PA, corresponding to the conformation in solution, are shown individually and as an overlay in cartoon representation. c. Gel-based cleavage assay assessing the catalytic cysteines. K63-linked tetraubiquitin chains (2 µM) were incubated with USP5320-383 (4 µM, upper) or USP5421-369 (300 nM, lower) and the respective catalytic cysteine mutants. Cleavage activity was analyzed by SDS-PAGE and Coomassie staining. d. Gel-based cleavage assay assessing the catalytic histidines. K63-linked triubiquitin chains (3 µM) were incubated with USP5320-383 (2 µM, upper) or USP5421-369 (300 nM, lower) and the respective catalytic histidine mutants for indicated time points. e. Gel-based cleavage assay as in d assessing potential oxy-anion hole residues. Uncropped versions of all gels are provided as Source Data.
Extended Data Fig. 7 Analysis of the weakened S1 ubiquitin recognition in USP54.
a. Cartoon representation of USP54~diUb-PA as well as of representative USP DUB-ubiquitin aldehyde complexes (USP7, PDB: 1NBF; USP14, PDB: 2AYO). A superposition is shown, illustrating the distinct relative geometry of ubiquitin in the S1 site of USP54. b. Close-up view on the interaction of the Phe4 patch in the S1 ubiquitin with USP54 (left) and USP2 (PDB: 2IBI, center). A superposition (right) (aligned on ubiquitin) illustrates the different environment of Phe4 in USP54. c. Close-up view on the interactions of BL1 in USP54 (left) and in USP14 (PDB: 2AYO, center) with the S1 ubiquitin. A superposition (right) based on the S1 ubiquitin is shown. d. Close-up view on the S1 ubiquitin C-terminal tail interactions with USP54 (left) and USP7 (PDB: 1NBF, center). Hydrogen bonds are illustrated by dotted lines in pink for USP54 and black for USP7. Arrows highlight the missing hydrogen bonds in USP54 in comparison to USP7, also visible in the superposition (right).
Extended Data Fig. 8 Structural analysis of cholestasis-linked mutations in USP53 and conservation of S2 site on back of USP fingers in USP53 and USP54 orthologues.
a. Homology model of the USP53 catalytic ___domain, based on the USP54 crystal structure. Residues of disease-associated single amino acid mutations are shown as red sticks (see Fig. 2a). b. Close-up view on Arg99 in USP53, highlighting bridging interactions to the switching loop and the Cys loop. c. Close-up view on the corresponding residue Arg100 of USP54 in the USP54~diUb(K63)-PA structure. d-e. Close-up view on Gly31 and Cys303 in the USP53 model, highlighting how substitution of these closely surrounded residues for amino acids with larger side chains in the patient mutations G31S and C303Y likely lead to structural perturbations very close to catalytic residues. f. Close-up views on the S2 site, highlighting how a loop in the USP54 thumb ___domain above the S2 site forms engages ubiquitin. g. Sequence alignment of representative USP53 and USP54 orthologues showing the first three USP fingers beta sheets. Secondary structures and numbers are indicated according to human USP54. Species include human (Homo sapiens), Macaque monkey (Macaca mulatta), mouse (Mus musculus), rat (Rattus norvegensis), cat (Felix catus), cow (Bos taurus), zebra finch (Taeniopygia guttata), golden eagle (Aquila chrysaetos chrysaetos), Indian cobra (Naja naja), painted turtle (Chrysemys picta bellii), zebrafish (Danio rerio) and atlantic salmon (Salmo salar). Residues conserved in all sequences are shown in black, the S2 aromatic residues are shown in grey. Cystein residues coordinating the zinc ion at the tip of the fingers are highlighted in red. Residues on the beta sheets with side chains pointing towards the backside / S2 ubiquitin binding site are highlighted with a black triangle. h. Coordination of the Ile44 patch in the S2 ubiquitin binding site. Cartoon representation of the USP54~diUb-PA structure. Ubiquitin residues of the Ile44 patch and residues highlighted with a black triangle in panel g are labeled as sticks.
Extended Data Fig. 9 A cryptic S2 site mediates catalytic activity on longer ubiquitin chains.
a. Purified samples of USP54 with mutated S2 site. USP5421-369 F161K, USP5421-369 F161K~Ub-PA and USP5421-369 F161K~diUb(K63)-PA were analyzed by SDS-PAGE and Coomassie staining. An uncropped version is provided as Source Data. b. Protein stability assessment of S2 site-mutated USP54. Stability of samples from panel a was analyzed by thermal shift analysis. Fluorescence raw data are shown. Melting temperatures (Tm) are plotted from technical replicates. c. Schematic of catalytic USP54 F161K DUB domains after reaction with Ub-PA or diUb-PA probes, illustrating ubiquitin engagement in samples used in panels a and b. S1’, S1 and S2 Ub binding sites are labelled. The mutated S2 site is depicted as a red star. d. Fluorescence polarization cleavage assays for USP5320-383 Y160K and indicated reagents. Fluorescence anisotropy was recorded over time and observed rate constants (kobs, shown as mean ± standard error) were plotted over enzyme concentrations. Values for WT protein were taken from Fig. 2g. e. Fluorescence polarization cleavage assays for USP5421-369 F161K and indicated reagents as described in panel d. f. Cartoon representation of USP54~diUb-PA, CYLD in complex with K63-linked diUb (PDB: 3WXG) and CYLD in complex with M1-linked diUb (PDB: 3WXF). A superposition illustrates features enabling K63-linkage recognition in both enzymes by distinct mechanisms. The catalytic ___domain of CYLD achieves dual K63 and M1 polyubiquitin cleavage specificity through a unique β-hairpin inserted into its catalytic ___domain. This insertion forms an S1’ ubiquitin binding site to recognize the Phe4 patch of the proximal ubiquitin (in both K63- and M1-linked chains) and positions the substrate for efficient catalysis. USP54 lacks an equivalent hairpin insertion.
Extended Data Fig. 10
Mechanisms for polyubiquitin length- and linkage-specificity in DUBs.
Supplementary information
Supplementary Information
Supplementary Tables 1 and 2 and References.
Source data
Source Data Fig. 1
Numerical source data.
Source Data Fig. 1
Uncropped gels and blots.
Source Data Fig. 2
Numerical source data.
Source Data Fig. 2
Uncropped gels and blots.
Source Data Fig. 3
Numerical source data.
Source Data Fig. 3
Uncropped gels and blots.
Source Data Fig. 4
Numerical source data.
Source Data Fig. 4
Uncropped gels and blots.
Source Data Fig. 5
Uncropped gels and blots.
Source Data Fig. 6
Numerical source data.
Source Data Fig. 6
Uncropped gels and blots.
Source Data Extended Data Fig. 1
Numerical source data.
Source Data Extended Data Fig. 1
Uncropped gels and blots.
Source Data Extended Data Fig. 2
Numerical source data.
Source Data Extended Data Fig. 2
Uncropped gels and blots.
Source Data Extended Data Fig. 3
Numerical source data.
Source Data Extended Data Fig. 3
Uncropped gels and blots.
Source Data Extended Data Fig. 4
Numerical source data.
Source Data Extended Data Fig. 4
Uncropped gels and blots.
Source Data Extended Data Fig. 5
Numerical source data.
Source Data Extended Data Fig. 6
Uncropped gels and blots.
Source Data Extended Data Fig. 9
Numerical source data.
Source Data Extended Data Fig. 9
Uncropped gels and blots.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wendrich, K., Gallant, K., Recknagel, S. et al. Discovery and mechanism of K63-linkage-directed deubiquitinase activity in USP53. Nat Chem Biol 21, 746–757 (2025). https://doi.org/10.1038/s41589-024-01777-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41589-024-01777-0
This article is cited by
-
N-glycosylated LTβR increases the Th17/Treg cell ratio in liver cancer by blocking RORC ubiquitination and FOXP3 transcription
Cell Death & Disease (2025)
-
Chimeric deubiquitinase engineering reveals structural basis for specific inhibition of the mitophagy regulator USP30
Nature Structural & Molecular Biology (2025)
