
Abstract
The extraordinary preservation of Cueva de Los Murciélagos (Albuñol, Spain) provides a unique opportunity to identify the materials and the techniques involved in archery during the Early Neolithic period. Arrows with preserved feathers, tied fibres, adhesive substance, and two probable bowstrings have been studied trough an unprecedented multi-proxy investigation, including microscopy and biomolecular methods, to unravel archery techniques. The study has identified the oldest known sinew bowstrings, the first evidence for the use of olive tree (Olea europaea) and reed (Phragmites sp.) to produce arrow shafts in prehistoric European archery, and the identification of birch bark tar as a coating on the shafts. The results of this study provide insights into ancient craft, technological solutions, and adaptations to local resources in the production of these reed-shafted hardwood tipped arrows and bowstrings. Their deposition in a burial cave sheds new light on the role of these artefacts in a Neolithic farming community.
Similar content being viewed by others

Introduction
The study of Neolithic archery has been primarily focused on two main aspects: projectiles and bows. When examining bows, a remarkable consistency in raw materials and production techniques is observed across different regions during the Neolithic1,2,3. Despite variations in size, the prevalent use of yew (Taxus baccata) wood suggests a shared technological tradition throughout Europe during this period. This uniformity implies a sophisticated understanding and transmission of bow-making techniques among these Neolithic communities.
Conversely, research on projectiles has unveiled a rich diversity in both raw materials and shapes. While lithic and bone projectiles dominate the archaeological record due to their frequent discovery4,5,6,7, there are also noteworthy instances of wooden projectiles, as evidenced in waterlogged sites8. This diversity underscores the adaptive strategies and resourcefulness of Neolithic societies in utilising a variety of materials for their weaponry.
Beyond the technical aspects, scholars have extensively debated the social and ideological significance of archery in the Neolithic context8,9,10. Bows and arrows have traditionally been categorised as tools primarily utilised for hunting purposes. Hunting, however, may have played a relatively minor role in Neolithic subsistence strategies, the persistence of archery elements and the diversity of projectiles suggest that archery held significance beyond mere sustenance, possibly linked to rituals, warfare, or other socio-cultural practices. For example, compelling osteological evidence has been found for their involvement in interpersonal violence during the Neolithic period in the Iberian Peninsula11,12,13,14, including cannibalism15, as well as across Europe16.
Other facets of Neolithic archery, such as the production of arrow shafts, remain relatively underexplored in the archaeological record. Factors such as the methods for affixing arrowheads to shafts, the materials and properties of the shafts themselves, and the characteristics of bowstrings have received scant attention, primarily due to the unlikelihood of such delicate remains preserving. Consequently, our understanding of these aspects largely relies on insights drawn from ethnographic studies17,18,19, and experimental archaeology20,21. Notable advances have been made through the study of materials recovered from unique environments such as the ice fields of the Alps3 and Scandinavia22, as well as waterlogged sites23. These discoveries have provided valuable insights into the diversity of raw materials used in arrow production and the associated manufacturing techniques.
The unique archaeological context of Cueva de los Murciélagos (Albuñol, Spain) offers new perspectives on the use of organic materials in prehistoric archery. A wealth of archery artefacts has been recovered from this cave, used as burial cave during, at least, the Neolithic (supplementary material fig. S1). Preserved through desiccation due to the dry conditions within the cave, the material includes shafts and arrows along with their associated organic components, including adhesive, feathers and fibres (Fig. 1), as well as two potential bowstrings (Fig. 2). Such preservation provides a rare and exceptional opportunity to study elements ordinarily absent in archaeology owing to their perishable nature. With the exception of a fragment of cord (CM-P287) found during the recent resumption of excavations at the site since 2022 (supplementary material, fig. S2.1), the artefacts were discovered by miners in the 19th century and no information is known about their surface provenance. Furthermore, all of these objects were found deposited in the cave, unstratified, which poses a challenge for the reconstruction of possible different funerary events that may have occurred at this site (see supplementary material for more details on the site).

Plant-based arrowheads and shafts from Cueva de los Murciélagos. (A) CM-1138/532 (Ua-78245), reed and wooden arrow shaft, (a.1.) Detail of white adhesive and tied fibres at the junction between the reed and the wood, (a.2.) Detail of tied fibres at the end of the reed shaft, (B) CM-1139/532 (Beta-692876), reed arrow shaft, (b.1–4.) Arrow notch, with feathers, tied fibres and black adhesive, (C) CM-1140 (Beta-692878), arrow shaft or point, (c.1.) Detail of the adhesive, (c.2.) Point of the arrow with adhesive, (D) CM-1141 (Ua-78246), arrow shaft or point, (d.1.) Spiral marks. © F. Martínez-Sevilla.

Cordage fragments from Cueva de los Murciélagos identified as possible bowstrings. (A) CM-P287 (Beta-684048), (a.1.) Knot or repair, (a.2.) Detail of twisted fibres; (B) CM-591b (Beta-692877), (b.1.) Broken/fringed cord end, (b.2.) Detail of twisted fibres with organic material between grooves. © F. Martínez-Sevilla.
A multidisciplinary approach combining biomolecular and archaeobotanical methods has been applied to elucidate the diversity of the organic raw materials, the production techniques and characteristics of these ancient projectiles. Traditional identification methods and techno-morphological studies provide crucial information on the identification of woody material and the morpho-technological characteristics of these artefacts, while biomolecular methods enable a more detailed understanding of the nature and properties of the amorphous substances and fibrous elements associated with the archery elements.
Results
Radiocarbon dating
Radiocarbon dating (Table 1) and Bayesian modelling (Fig. 3 and supplementary table S1) revealed that five of the artefacts - a composite object made of wood and reed (CM-1138/532), a reed stem (CM-1139/532), a wooden point (CM-1140), and the two cords (CM-591b, CM-P287) - produced consistent dates for the late 6th millennium and early 5th millennium cal BCE. These new radiocarbon dates, and those previously obtained24, place this archery equipment in the Early Neolithic occupation phase of the cave - a period when different objects were deposited in the cave as grave goods. A single object, CM-1141, can be linked to the Bronze Age, with a date between 1960 and 1745 cal BCE (95% of probability), which corresponds to the last documented phase of prehistoric use of the cave.

Probability distribution of radiocarbon dates from Cueva de Los Murciélagos, including those from this study (in blue) and previously published dates24. Multiphase Bayesian chronological ranges for the estimated start and end of each phase and the modelled ranges of each radiocarbon date. Objects studied in this paper are in blue. OxCal v4.4.4 Bronk Ramsey; r:5 Atmospheric data from Reimer et al. ©J. Santana.
Morpho-technological descriptions and raw material identification
The corpus of objects studied in this research led us to classify them into two groups of wooden objects. The first group includes composite arrow shafts with wooden points, cane shafts, and feathers, with wound fibres fixing the different elements.
The branches were debarked and polished as not cut marks or other traces were observed. This group is dated to the Early Neolithic.
-
CM-1138/532, fragmented at both ends, has an overall length of 228 mm, and is a combination of a wooden arrowhead made of a twig of Salix sp. (willow) and a Phragmites sp. cane (reed) for the shaft (shaft = 82.1 mm) (Table 2; Fig. 4a, b and c). The shaft has a diameter of 9.7 mm, narrowing to 8.4 mm near the wood attachment, and tapering slightly to 8.2 mm at the end. The reed and wood are secured by wound fibres, fixed with an adhesive that was also found on the wooden part.
-
CM-1139/532 is a shaft made of a Phragmithes cane (Table 2), measuring 180 mm in length and 8 mm in diameter. Preserved at one end of the reed shaft, near the notch, are the remains of two feathers tied with fibres, while the other end is also bound with fibres. This shaft is the oldest fletched arrow in European prehistory The shaft is coated with a black substance, possibly indicating treatment for preservation or added functionality.
-
CM-1140 is a wooden point made with a twig of Olea europaea (olive tree) (Table 2; Fig. 4d, e and f), measuring 145 mm in length, the proximal part has a diameter ranging from 6.7 mm to 7.2 mm. The segment that may have been inserted into the shaft measures 52 mm in length, the tip itself extends for 93 mm. The tip is covered with a black substance for possible reinforcement or preservation.
Morphologically the fourth object is quite different, and is dated to the Bronze Age.
-
CM-1141 has been interpreted as a shaft made of a wooden twig of Acer sp. (maple) (Table 2; Fig. 4g, h and i) that appears to be decorated with a dark spiral pattern (Fig. 1d.1.). With a length of 270 mm, its diameter varies at 10 cm intervals: 6.6–7.1 mm − 6.1 mm. One end retains the original shape of the branch, while the other is tapered and appears somewhat oval in cross-section, with longitudinal marks visible. The surface of the tapered part of the branch is polished.

SEM microphotographs of wood artefacts CM-1138/532-1 (Salix/Populus, a: cross section, b: tangential section, c: radial section), CM-1140 (Olea europaea, d and e: cross sections, f: radial section) and CM-1141 (Acer sp., g: tangential section, h and i: radial sections). © M. Martín-Seijo and Raquel Antón.
The two cord fragments are similar in morphology. They are in good condition, although they have a few breaks at the ends.
-
CM-591b is approximately 34 cm long by 3 mm in diameter. This two-ply cord has a Szz twist with a closed twist angle (53º − 60º) and 2 twists per centimetre.
-
CM-P287 is a two-ply cord 59 cm long by 3 mm in diameter. It also has a Szz twist with 2 twists per centimetre, with a twist angle of 55º-60º. The addition of fibres to lengthen the cord can be seen in different spots. A knot is visible, which might be a repair.
Identification of fibres
The observations made with the optical microscope of the two cords (CM-P287 and CM-591b) and from the ties of the shafts CM-1138/532 and CM-1139/532 (Supp. material fig. S4) led to the conclusion that the fibres were not of plant origin, as none of the observed characteristics corresponded to those of a plant organism. The cords were then analysed with SEM, however, the anatomy of the ties was more difficult to observe because they were held together by an adhesive substance which could not be completely separated from the fibres before microscopic examination. Parallel fibres in the longitudinal length were better observed on the two cord samples, revealing the hierarchical structure of undulated fibres (50–300 μm) composed of smaller fibres of 1–20 μm, which is similar to the structure of collagenous animal tissue (Fig. 5).

SEM microphotographs of the cords (a) CM-591b: structure of fibres with an undulated aspect, (b) CM-P287: detail of the collagen fibres, (c and d) modern sinew ©I. Bertin and M. Herrero-Otal.
The animal origin of the cords was confirmed via elemental analysis, with a dominance of elements that are found in connective tissues like ligaments or tendons (supplementary material fig. S5 and table S2). Fourier Transform Infrared (FTIR) spectroscopy (supplementary material fig. S6 and table S3) on the cords and tied fibres also showed absorption bands consistent with collagen25,26, the main component in animal sinew. All spectra (supplementary fig. S7, S8 and table S4) showed distinct peaks at around 1630 cm-1 and 1540 cm-1, corresponding to the amide I and amide II bands, respectively. These bands indicate the presence of collagen-rich tissues. Importantly, the absence of characteristic peaks associated with other major tissues, such as prominent phosphate bands for bone or lipid bands for muscle, and the comparison with modern sinew confirmed the specificity of the observed signals for sinew.
In order to identify the taxa, Zooarchaeology by Mass Spectrometry (ZooMS) was performed using a small sample of each cord. CM-591b was identified as deriving from Capra spp., while CM-P287 exhibited peptide markers for a mixture of taxa, including Sus spp. and likely Caprinae and/or Cervidae (Supplementary Table S4 and Supplementary Fig. S7), indicating that the cord was made from the sinew of multiple species. To test this further, an additional sample, CM_P287-B, was taken from the opposite end of the cordage and analyzed via ZooMS, producing a single identification of roe deer (Capreolus capreolus), confirming the mixing of taxa. An additional marker of high intensity at m/z 2115 was observed in all three samples, and also at m/z 1299 CM-591b (Supplementary Fig. 7). While m/z 2115 is a known collagen type I peptide marker for certain taxa (for example, humans), the other markers present do not coincide with these taxa, suggesting this marker actually represents a different, unknown peptide, potentially from a different type of collagen (such as collagen type III, present in lower quantities in tendon and ligament tissues) or other proteins associated with connective tissues.
Adhesive analyses
Organic residue analyses were performed on 5 adhesive samples from three of the projectiles (CM-1138/532 with two samples (b and c), CM-1139/532 with one sample (b), and CM-1140 with one sample taken near the tip of the arrow (c) and one at the junction between arrowhead and shaft (d)). Two of the samples taken from the composite arrow CM-1138/532 yielded insufficient material to obtain an identification (absence of chromatogram peak). The absence of solvent soluble organic compounds may be attributed to a high yield of carbonisation or to the presence an undocumented modern restoration presumed to be a polymerised insoluble material. The three other samples revealed compounds characteristic for birch bark tar (Fig. 6).

Chromatograms of the adhesive residue on (a) CM-1139/532, (b) CM-1140c and (c) CM-1140d with peaks consistent for birch bark tar. The asterisk in CM-1139/532 corresponds to trace amounts of the following fatty acids, from left to right: Tetradecanoic acid (C14:0); Pentadecanoic acid (C15:0); Heptadecanoic acid (C17:0); (9Z)-Octadec-9-enoic acid (C18:1); Nonadecanoic acid (C19:0); Heneicosanoic acid (C21:0); Tetracosanoic acid (C24:0). ©I. Bertin.
Three main classes of biomolecular markers were detected and identified in the three samples: triterpenoid compounds, long-chain even-numbered fatty acids, and saturated short-chain α,ω-dicarboxylic acids (diacids). The samples from the arrowhead CM-1140 and the shaft CM-1139/532 are largely dominated by the triterpenoid constituents, while fatty acids and diacids are more diverse in the adhesive of CM-1139/532 than in that of CM-1140. For CM-1139/532, the triterpenes constitute 81.4% of the tar and the fatty acids and diacids 18.6%. For CM-1140, the sample of tar from the tip of the arrow (sample c) is composed of 77.5% triterpenes and 22.5% fatty acids, mainly represented by palmitic and stearic acids, while the sample taken near the junction of the arrowhead and shaft (sample d) has a low proportion of fatty acids (3.6%), with 96.4% triterpenes (supplementary table S5 and S6). All the triterpenes identified, such as betulin, lupeol and their derivatives, are from birch bark tar (details in the Supplementary Appendix). Most of the fatty acids probably originate from the degradation of suberin under the influence of heat (Ekman, 1983; Rageot et al., 2019). The high concentration of palmitic and stearic acids, particularly important in CM 1139/532b, could indicate the addition of a fatty substance to the adhesive, but could also result from the manipulation or use of the objects (past or recent).
Discussion
These archery remains demonstrate the technical skills and expertise of the early farming societies in southern Iberia. These communities applied their knowledge to the acquisition and transformation of various raw materials, requiring a deep understanding of organic material properties and the techniques necessary to convert them into useful products. Both animal and plant raw materials were used to produce the archery elements, with different materials selected for specific uses. In prehistoric Europe, the plant raw materials used for bows and arrows identified to date are: Pinus sp., Viburnum lantana, Buxus sempervirens, Cornus sp., Carpinus, Betula sp., and Corylus avellana8,27,28,29,30,31(supp. Material, table S7); thus making the identifications herein of both Salix and Olea unique evidence for arrow making.
According to pollen records, the plants used for making the arrow shafts and points, and the adhesive grew at the vicinity of the site and could have been sourced locally32,33,34,35. The vegetation was characterised during the 6th millenium cal BCE by the passage from pinelands, with Pinus tp. sylvestris-nigra, to a Mediterranean mixed oak-pine forest, along with other mesophilous angiosperms32,33,34,35. The use of olive (Olea europaea) wood as raw material for crafting several artefacts at Cueva de los Murciélagos, such as a mallet in addition to the wooden arrow point, reflects the relevance of this species in the prehistoric communities that inhabited in the Mediterranean area, which until now had only been attested as fuel36,37,38,39, or in the carpological record with the presence of wild olive stones at Cueva de los Murciélagos de Zuheros40,41. Willow wood (Salix sp.) is also only documented as fuelwood during Early Neolithic times in the southern Iberian Peninsula. Reed grasses, such as Phragmites, are found in temperate wetlands and grow nowadays in the ravines and streams close to the cave, and have been documented in the chalcolithic funerary cave of Murcia in the form of a 20 cm long fragment42.
The lack of other evidence for olive wood in arrow production is likely a reflection of its natural geographic distribution not coinciding with the geographic locations of most archaeological sites where archery evidence has been found. Yet, olive wood is very dense and is known for its strength and durability, while still maintaining a good amount of flexibility43. This balance between strength and flexibility (young’s modulus) is crucial for arrow shafts, as they need to be able to bend without breaking when released from a bow. Moreover, the high density of olive wood contributes to the overall weight of the arrow, which can enhance penetration and stability during flight. Heavier arrows made from dense woods like olive tend to face less resistance to wind and maintain their trajectory better44, especially over longer distances. Finally, olive wood can be polished to a very smooth finish, and a smooth arrow shaft reduces friction as it leaves the bow, resulting in a more consistent and accurate flight. While willow wood is not as strong or dense as olive wood, its advantages in terms of flexibility, weight, and workability make it a good choice for certain types of arrows, particularly those designed for speed and ease of crafting. The use of willow as raw material for making arrow shafts has been attested too at La Draga, an Early Neolithic site in the north-eastern Iberian Peninsula45. Willow is a relatively light wood which can be advantageous for creating fast arrows. Lighter arrows can travel at higher speeds, making them suitable for certain types of archery, particularly where speed and distance are prioritised46. Moreover, willow is relatively easy to work with due to its softness compared to harder woods, like olive. Although the properties of willow may vary from species to species47, the use of willow can be linked to its small straight lightweight shoots with high elasticity and impact resistance. Similarly, Phragmites are particularly suitable for archery due to their straightness and good resistance, provided that a few criteria are taken into account when selecting the pieces20. This material has been documented in several ethnographic19 and archaeological contexts, especially in Egypt48,49,50, however not in prehistoric Europe. The presence, however, of this material in many regions and its advantages as an arrow make it a favoured material, and it is conceivable that past populations also used it despite its lack of preservation in the archaeological record.
Regarding the manufacture of arrow shafts, the crafters likely took advantage of the original shape and diameter of perfectly straight, knot-free stems/branches with a certain weight/length ratio. Willow and olive branches tend to grow relatively straight, as does reed, which is a desirable characteristic for arrow making as it requires less correction during the manufacturing process. The selection of these specific natural morphological properties of fresh wood, or wood that has been freshly cut and then dried to improve stiffness, even independent of the species itself, could explain the variety of materials with different properties used. Indeed, the principle of wood selection based on non-objective criteria or not determined by the species concept is well described by Levi Strauss51.
Reed and wooden arrows have some undeniable qualities when it comes to evaluating their hunting efficiency. Recent experimental research on reed stems and reed arrows showed a nice set of ballistic properties20. This type of material confers stiffness, good impact resistance, stable flight with moderate rotation, high degree of target accuracy, and important perforation capacity to an arrow. The velocity of such arrows can reach over 50 m/s when shot with replicas of a Neolithic self-bow or a slight recurve bow of Neolithic Bolkóv type20. Similar velocities were measured on replicas of Apache arrows made from reed for the shaft and dogwood (Cornus sp.) for the fore shaft, and shot with a traditional Apache mulberry bow. The range of these composite arrows was less than 20 m, making them suitable for short-distance hunting and fighting19. Clark also mentions the good penetration and cutting of Egyptian reed and hardwood arrows with a stone point (his type A), emphasising the superior penetration of arrows with hardwood points (type C)49. Further experiments could be carried out to confirm the flight, speed and cutting ability of the arrows from Cueva de los Murciélagos, which are shorter than those in the above-mentioned experiments, and to verify whether they could have been functional for hunting or killing at close range, or whether they could have been non-lethal arrows (e.g. for the recovery of feathers not stained with blood), which would explain the use of Salix sp. Despite the exceptional preservation of arrows and other organic and wooden objects in the cave, no information is known about the type of bow used, as no bows have been identified. This could be due to the absence of bows with the burials, whether they were reused or deliberately excluded. It is worth noting, however, that several artefacts might have been lost or destroyed at the time of discovery. As described by Gongora y Martínez in 186852, the miners who found most of the objects were not meticulous nor attentive to the material deposited in the cave, if bows were present, they may have not survived.
The identification of the twisted cords from Cueva de los Murciélagos as animal sinew from at least three different taxa (Capra spp., Sus spp., and Capreolus capreolus) is coherent with the faunal record from the site. The material found throughout the recent campaigns show a dominance in terms of biomass of domestic caprines. Wild boar and deer were also present in the cave, along with rabbit and hare, and certain birds such as the partridge, documenting hunting activities. In the absence of stratigraphy at the site, the precise age of the elements included in the assemblage is unknown and the chronology is heterogeneous, but similar findings were obtained at the contemporaneous Neolithic Zacatín rockshelter (Gualchos-Castell de Ferro)53. Bowstrings are rarely identified within prehistoric archaeological records due to their rapidly degrading organic nature. For the Early Neolithic in Western Europe, a bowstring has been found at La Draga, made of herbaceous stems (nettle), which are known to be resistant and can endure the stretching required of a bowstring23. Evidence of sinew or gut bowstrings have been documented for the Late Neolithic of Europe, in two snow-patches from Italy and Switzerland3. In the Tisenjoch ice-patch, two bowstring fragments were associated with the remains of Ötzi, dated around 3300 cal BCE. An unprocessed frozen bundle was identified as animal leg sinew, while a finished twisted cord was made of animal fibres3. At Schnidejoch, a cord dated to 2800 cal BCE was identified as animal sinew fibre by microscopy, but the species of the animal could not be identified54. For arid contexts, in more recent periods, several fragments of gut bowstring still attached to bows were described in Egyptian graves, such as the tomb of Tuntankhamon55. Although they were not found attached to a bow, the interpretation of the Murciélagos cords as bowstrings is based on their morphology, which is consistent with other Neolithic bowstrings found at Tisenjoch and Schnidejoch. All these bowstrings are S-twisted cords made of three separate strands of fibre with a thickness of 3–4 cm3, which gives the bowstring sufficient stretch and resistance. One problem that arises in the production of bowstrings is that the material is not always long enough to achieve the desired length. It is therefore necessary either to choose a long tendon/muscle, as the Apaches of North America did by choosing the back of a deer’s leg to make their bowstrings19, or to combine different sinew fragments. At Cueva de los Murciélagos, the latter seems to have been the method of choice, at least for CM_P287, as evidenced by the visible addition of different sinew fragments, especially at the knot, and by the ZooMS identification of at least two different animal taxa. The production processes of bowstrings have been detailed in several ethnographic works18,19. The Western Apaches and Chiricuas of North America were soaking and splitting deer leg sinew into strips, before doubling and twisting them together. Sinew could also be applied on a weakened bow to consolidate it19, and was attached with glue (boiled hooves and horns, and sometimes pinion pitch). The bowstrings at Cueva de los Murciélagos could have undergone similar processes to prepare the S-twisted cords.
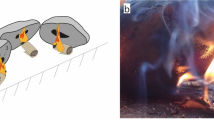
The presence of birch bark tar on the arrow point and the shaft is the first attestation of the use of birch bark and its adhesive properties in the Early Neolithic of Andalusia. Birch was present in the Early Neolithic, in the Mediterranean mixed oak-pine forest along with the mesophilous angiosperms32,33,34,35 or on the lower flanks of the Baetic Mountains, with xerothermophilic scrub and pine forests56. Thus collecting birch bark could be linked to local supplies in different environments and vegetation layers, along with the other taxons used for wood manufacture. The absence of artefacts made of birch wood at Cueva de los Murciélagos, and its absence in charcoal records from contemporaneous settlements, could illustrate a specific management of this taxon, focused on its bark for tar production, which suggests a specific knowledge about its properties and the selection of the appropriate bark quality. The thin regular layer of birch bark tar covering the preserved part of the arrow shaft CM-1139/532 and the arrow 1140 suggests that the object may have been dipped directly in the liquid tar, or that the tar may have been brushed all over the object. The first bark exudates obtained experimentally during per descendum experiments allows a low viscosity tar with a slow solidification time after removal from the heat source57,58. At Cueva de los Murciélagos, the relative absence of strong heating markers correlated with the high proportion of biomarkers in the tar is coherent with the fabrication process of a first exudation of tar. While the tar from CM-1140 has fewer fatty acids, thus more similar to experimental tars produced via condensation, the high proportion of biomarkers and low heating markers suggest it was likely produced per descendum. These variables, between the two objects but also the two samples of 1140, could be a result of anthropic alteration and post-depositional degradation. Caution must be used when comparing archaeological data with experimental data. Thus, we can only conclude that the production of the birch bark tar used at Cueva de los Murciélagos involved a short heating time at temperatures below 350 °C.
There are several possible functions for the birch bark tar coatings on the arrow CM-1140 and the shaft CM-1139/532. A similar coating was noted for other composite objects at Krzyż, an Early Mesolithic site in northern Poland59,60. In this case, it was suggested that in addition to hafting properties, the coating could also have been used for waterproofing properties or to consolidate the wooden point59. At Cueva de los Murciélagos, the addition of birch bark tar to an object made from olive wood, as is the case for CM-1140, for waterproofing purposes seems unlikely as olive is known to be compact, resistant, and naturally impermeable. It is plausible that the adhesive properties of the tar were used to hold together the various elements of this composite object, such as the sinew fibres and feathers, during projection. The tar does not seem to have played a role in ensuring cohesion between the arrowhead and the shaft, however, as no residue was observed on the part of the arrowhead that was presumably inserted into a reed shaft. Moreover, other adhesive substances were present on CM-1138/532, used to maintain the sinew ties. Thus a different substance may have been used for joining the two pieces. In Bronze Age contexts in Egypt, such as Naga-ed-Der, similar reed-shafted hardwood tipped arrows are not (or only rarely) entirely coated, and the adhesive is applied only between the shaft and hardwood tip, maintaining fine fibres or threads that secure the junction49. Another hypothesis is that the tar at Cueva de los Murciélagos was used as decoration. The decoration of bows and arrows has been documented ethnographically19 and archaeologically49, with the presence of black, red or blue paint. The decorative function of birch tar has been documented during the Neolithic on diverse materials, such as jewellery58. The shaft or spear dated to the Bronze Age, CM-1141, also displayed decorative black marks, however its composition could not be identified. Raman spectroscopy could be used in the future to try to characterise this substance, provided that the fluorescence phenomenon is not too strong.
The symbolic significance of the archery equipment is particularly notable within its funerary context. The archery equipment is part of an exceptional set of grave goods found in this cave, including remains of clothing (whole or fragmented sandals, fragments of cloth), mats, wooden or fibre-based containers, remains of rope and a torch. In addition, artefacts directly linked to agricultural activities, such as a sickle and a wooden stick, were documented in publications by Góngora y Martínez52. Unfortunately, the position in the cave and the association of these objects with each other and with the human remains is poorly known. The deposit of at least two hunting or warfare weapons in a funerary context related to a farming society poses the question of the role of hunting and hunters, but also of violence in this society. Evidence of intra-personal violence and cannibalism is well documented from other similar Neolithic contexts such as El Toro15, Carigüela, Las Majólicas, Mármoles, and Malamuerzo61,62. It remains unclear, however, whether the instances of cannibalism reflect acts of violence or ritualistic practices. These findings suggest a complex social dynamic, where the manipulation of human remains played a role. Spanish Levantine rock art provides further evidence, depicting numerous scenes of group violence involving archers63. In the Upper Segura region, these scenes portray fights, battles, or ambushes. Although the origins of Levantine rock art remain unclear, the recurring depictions of archers emphasize their importance throughout prehistory. These representations, however, are difficult to interpret in terms of the actual intensity or nature of these violent encounters due to dating limitations of the paintings and the lack of direct physical evidence of violence, such as arrow wounds, which do not appear in Iberian funerary contexts before the Middle Neolithic64.
At Cueva de Murciélagos, the burial of human remains alongside archery equipment suggests a special significance of this objects for the early Neolithic societies. Moreover, this equipment could signify the continued importance of hunting in early Neolithic Andalusian societies. While domesticated animals constituted the majority of the species used, wild animals remained a significant source of food and other resources. Although it can be difficult to pinpoint the social role of hunting in archaeological societies, some authors have suggested that in early agricultural communities hunting was linked to social power, gender roles, and perceptions of space and time66. Levantine rock art depicting hunting scenes also emphasizes the importance of big-game hunting as an activity tied to social recognition, which persisted and even intensified throughout the Neolithic67. Hunting in these societies was a complex phenomenon, influencing social structures and individual identities, and often serving as a means of negotiating social and political roles66. Consequently, the association of these hunting objects with human burials could point to hunting as a cultural marker of social status.
Methods
Description of artefacts
Based on their shape and size, the wood and reed artefacts (CM-1138/532, CM-1139/532, CM-1140, CM-1141) were proposed to be the remains of arrowheads and/or shafts. The two cord fragments (CM-591b, CM-287) were found to have different morphological and technical features compared to other cordage from the site, and thus were proposed to be bowstrings.
Radiocarbon dating and bayesian modelling
Six archery artefacts were directly radiocarbon dated using AMS at Beta Analytic Laboratory (Miami, USA) and Tandem Laboratory, Uppsala University (Uppsala, Sweden). These measurements are based on terrestrial short-lived plant remains. This ensures a close relationship between the isotopic event measured by AMS and the depositional event targeted for radiocarbon dating. These radiocarbon dates were then subjected to Bayesian modelling to enhance accuracy and reliability in estimating the start, end, and duration of the artefact deposition in Cueva de los Murciélagos. We also incorporated 19 previously published radiocarbon dates from Early and Middle Holocene occupations of the cave, pertaining to the Mesolithic and Neolithic periods67. Analyses were performed using OxCal online software version 4.4 (Bronk Ramsey 2021). We applied the two-sigma probability interval (95.4%) for discussing the14C measurements and included the one-sigma probability interval (68.2%) in the tables and figures, as recommended by Millard68. All radiocarbon measurements were calibrated with the internationally recognized IntCal20 atmospheric calibration curve69.
The uncalibrated radiocarbon dates were modelled using a Multi-Phase Model70. This model integrates the radiocarbon dates into a uniform distribution based on the hypothesis that all dated events have an equal likelihood of occurring at any time within the phase. Additionally, our study utilised the Charcoal Outlier Model71,72, which typically yields younger age estimates because the correct age of the modelled events is younger than the non-modelled calibrated dates. The Charcoal Outlier Model assigns charcoal samples a 100% prior probability of being outliers and adjusts each sample using an exponential distribution to improve dating accuracy. The model was developed using OxCal tools (OxCal’s Sequence, Phase, Boundary, and Interval). Outlier radiocarbon dates were excluded from the Phase modelling. The modelled dates are rounded to the nearest half-decade, as the results can vary slightly with each run (Supplementary material Table S1).
Morpho-technological analysis
The analysis of arrowheads and shafts included their description at a macroscopic level, drawings and photographs. In respect to the morphology, the elements were measured (diameter and length) and their shapes described. The materials and morphometry of the different elements of the composite objects were also recorded. The cord fragments were measured (diameter and length) and their technical features described, including direction of twist, number of strands, type and angle of torsion, etc73. To study the technical aspects behind their production, the information was organised according to the different stages of their chaîne-opératoire74, from raw material procurement to the final object and its deposit as grave good75.
Taxonomic identification of plant-based materials
Wood and reed analysis
The three wood (CM-1140, CM-1141, CM-1138/532) and two reed (CM-138/532, CM-139/532) objects were studied at the Museo Arqueológico Nacional (MAN) and the Museo Arqueológico y Etnológico de Granada (MAEG). They were photographed with a Nikon D5000 camera with a NIKKOR AF-S DX 18–55 mm VR lens, and more detailed images were captured using a portable digital microscope (Dino-Lite Edge Digital Microscope AM7915MZT) with a magnification of x20-220. Along with their morphological and technological study, small fragments (1–2 mm) were sampled from breaks taking care to preserve the original morphology of the objects (CM-1140, CM-1141, CM-1138/532). For the wood objects, the three anatomical sections of wood (cross, tangential and radial) were observed under Scanning Electron Microscope (ZEISS EVO LS 15, RIAIDT, Universidade de Santiago de Compostela). The diagnostic features were compared with atlases for the identification of European woods76,77,78. In the case of Salix/Populus (willow/poplar), the main anatomical features that led to this identification were the diffuse to semi-ring porous wood, with solitary pores or short radial files of 2 to 3 pores (Fig. 4a), uniseriate rays (Fig. 4b), large, simple ray-vessel pits (Fig. 4c) and simple perforation plates72. The distinction between Salix and Populus is not possible when dealing with small wood fragments, because the difference between both genera is based on the number and morphology of ray cells72. Olea europaea (olive) wood is diffuse-porous, with pores generally in short radial multiples of 2 to 4 pores (Fig. 4d and e), rays bi- to tri-seriate, heterogenous with one to 3 (6) square and upright marginal cells (Fig. 4f)72,73. Finally, the criteria for identifying Acer sp. (maple) were its diffuse-porous wood with small, evenly spaced vessels, multiseriate rays (Fig. 4g), perforation plates simple and spiral thickenings in all the vessels (Fig. 4h) and presence of prismatic crystals (Fig. 4i)72. Attending to the ray width and to the presence of crystals it might be classified in the Acer campestre group74.
In parallel with anatomical identification, dendrological attributes were recorded to obtain complementary information about the raw material, such as plant part used and tree-ring curvature, amongst others79, as were taphonomic features, such as evidence of wood biodeterioration80.
Phragmites australis (reed) can be directly identified by the macroscopic characteristics of its hollow stem. Additionally, following the same procedures as the wood elements, cross section and epidermis were observed under an Olympus BX51 microscope equipped with an Olympus DP26 camera and connected to Olympus cellSens software, at the UAB Archaeobotany Lab. The morphology of the stem (surface and cross section) was compared with plant atlases for taxonomic identification and with modern plants as reference material76,77. Vascular bundles are large and arranged in a distinct ring close to the outer edge of the stem. The vascular bundle is composed of two large xylem vessels and phloem tissues, with thick-walled sclerenchyma fibres surrounding them.
Fibre identification
Small fragments of a few millimetres of fibres were sampled from the tied fibres and around 1 cm of material were recovered at the broken edges of the two cords. Identification procedures of fibres collected from the artefacts at the MAN (CM-1138/532, CM-1139/532, CM-591b) and from the cord recovered during the excavation campaign in 2022 (CM-P-287), were carried out under magnifications ranging from ×50 to ×500, using the aforementioned bright-dark field Olympus BX51 microscope at the UAB Archaeobotany Lab. Additionally, they were studied using Scanning Electron Microscopy (Zeiss Merlin, Servei de Microscòpia i Difracció de Raigs X, Universitat Autònoma de Barcelona). The samples were not carbon coated. These archaeological fibres were compared with modern sinew samples sourced from Ángel de la Llave, a traditional archer, who informed us that the sinew was obtained from horses (Equus sp), deer (Cervus sp.), or water buffalo (Bubalus bubalis). He typically purchases sinew from a traditional archery shop but also acquires materials from other sources and he could not specify the exact provenance in this case. The modern sinews were processed using traditional methods of cleaning, drying, and stretching.
Elemental analyses, spectroscopy and proteomic analyses
Elemental analysis was carried out on the two cords (CM-591b, CM-P287) and the modern sinew using an EDS Oxford LINCA-X-Max detector (Servei de Microscòpia i Difracció de Raigs X, Universitat Autònoma de Barcelona) at a resolution of 15 kV. The fibres have not been coated with carbon for possible re-use of the archaeological samples. Fibres from the cords and the wound fibres were also analysed by Attenuated total reflectance Fourier transform infrared (ATR-FTIR) spectroscopy (Perkin Elmer Frontier, at CEPAM, Université Côte d’Azur), a non-invasive and non-destructive method. The fibres were pressed and scanned in one position for 64 scans in the range 4000 and 400 cm−1 with 2 cm−1 resolution. The spectra were processed with software Spectragryph v1.2.16.1 and compared with published data25,26,81.
As the aforementioned analyses suggested both cords were of animal origin, Zooarchaeology by Mass Spectrometry (ZooMS) was performed to attempt taxonomic identification following a modified method as described in Buckley et al.82. ZooMS is a method of peptide mass fingerprinting which uses amino acid variations in collagen type I to taxonomically identify faunal materials, such as bone, antler, ivory, leather, and, as in the case of the cords, sinew. A few fibres 3–4 mm in length were selected from each cord and submerged in 100 µl of 50 mM ammonium bicarbonate (AmBic, (NH4)HCO3, pH 8) then heated at 65 °C for 1 h to gelatinize the collagen. 50 µl of the supernatant was transferred to a new 1.5 ml tube and 0.4 µg of trypsin was added, then incubated overnight at 37 °C. The digestion activity of the trypsin was stopped by acidifying the samples to 1% trifluoroacetic acid (TFA), and the samples were desalted using Pierce™ C18 zip tips (100 µl, ThermoScientific) and the peptides eluted in 50 µl of 50% acetonitrile, 0.1% TFA. 1 µl of sample along with 1 µl of matrix (α-cyano-4-hydroxycinnamic acid) were spotted in triplicate on to a Bruker ground steel MALDI plate and analyzed on a Bruker ultrafleXtreme MALDI-ToF-MS (CoEMS, University of York). The spectra were averaged and analysed using mMass83, and compared with a database of published collagen peptide markers82,84,85.
Organic residue analysis of black adhesives
A few milligrams of each residue were sampled from CM-1138/532, CM-1139/532 and CM-1140. For CM-1138/532, adhesive from the wooden part (b) and a black residue (c) from the edge of the artefact were sampled. A black coating was sampled from the reed shaft of CM-1139/532, as were two spots on CM-1140, one near the tip (c), and the other near the possible shaft attachment where the residue appeared slightly white (d) (supplementary S3). Although lipid analysis is a destructive technique, careful sample management was employed to maximize the analytical yield from the available material.
Lipids were extracted following a published solvent extraction method adapted to adhesives (Rageot et al. 2019 ; Rageot et al. 2021 ; Koch et al. 2024). The samples were sonicated twice for 15 min in a ratio of 2–3 mg/mL of dichloromethane (DCM, CH2Cl2). A volume of 100 µL of the resulting supernatant was recovered and evaporated under a flow of nitrogen at 40 °C. 50 µL of BSTFA (N, O-bis(trimethylsilyl)trifluoroacetamide) was added to the dried residues and heated to 70 °C for 1 h to derivatize the hydroxyl groups. The lipid extract was then solubilized in 100 µL of DCM with an internal standard. 1 µL of sample was injected with a blank control into a Shimadzu GC2010Plus mass spectrometer (CEPAM, Université Côte d’Azur, CNRS, France) equipped with a splitless injector (purge flow of 2.0 mL/min and split ratio of 3.0). A 30 m length DB-5MS (30 m × 0.25 mm i.d.; 0.25 μm film thickness) non-polar capillary column was used, with helium as the carrier gas and programmed at 3mL/min. The ion source temperature was 200 °C and the temperature of the transfer line between the GC and the MS was set at 280 °C. The temperature rise programme was as follows: from 50 °C, held for 2 min, to 150 °C at 10 °C/min, and then increased to 320 °C at 4 °C/min, and held for 15 min. Mass spectra were acquired using electron ionisation at 70 eV. The mass range was scanned, covering masses from 50 to 950 m/z.
The spectra acquired were analysed using GCMS PostRun Analysis software, and matched against previously published data86,87,88,89, the NIST library 2008 edition, and the MaxBirch3 library of birch bark tar standards by Maxime Rageot at CEPAM laboratory. The peak areas of fatty acids, biomarkers (BM), hydrocarbon pentacyclic triterpenes (TPH), doubly degraded pentacyclic triterpenes (TPD2) and other pentacyclic triterpenes derived by BSTFA (TPtms) were integrated, and their proportions in relation to the most abundant peak, reduced to 100%, were calculated. The ratios of biomarkers to total triterpenes, and the ratios of the different types of triterpenes were calculated and compared with other experimental and archaeological results57,60,90,91,92. A degradation index (DI) was calculated to help understand the natural and/or anthropogenic origins of the adhesive degradation24.
Data availability
MALDI-ToF-MS and GC-MS data analysed during the current survey will be available from the ZENODO repository (https://zenodo.org), under DOI: 10.5281/zenodo.13624814. Additional data related to this paper may be requested from the authors on request. Correspondence and requests for materials should be addressed to I.B.
References
Junkmanns, J. Arc et Flèche. Fabrication et Utilisation Au Néolithique (Éditions du musée Schwab, 2006).
Junkmanns, J. Pfeil Und Bogen: Von Der Altsteinzeit Bis Zum Mittelalter (Verlag Angelika Hörnig, 2013).
Junkmanns, J., Klügl, J., Schoch, W. H., Di Pietro, G. & Hafner, A. Neolithic and bronze age archery equipment from alpine ice-patches: a review on components, construction techniques and functionality. Jnanabha 283–314 (2019).
Shea, J. J. The origins of lithic projectile point technology: Evidence from Africa, the Levant, and Europe. J. Archaeol. Sci. 33, 823–846 (2006).
Borrell, F. & Štefanisko, D. Reconstructing projectile technology during the Pre-pottery Neolithic B in the Levant: An integrated approach to large tanged points from Halula. J. Archaeol. Sci. 69, 130–142 (2016).
Choyke, A. & Bartosiewicz, L. Osseous projectile points from the Swiss neolithic: Taphonomy, typology and function. In Violent Interactions in the Mesolithic. Evidence and Meaning (ed Roksandic, M.), Vol. 1237 (British Archaeological Reports, 2004).
Cortell-Nicolau, A., García-Puchol, O., Barrera-Cruz, M. & García-Rivero, D. The spread of agriculture in Iberia through approximate bayesian computation and neolithic projectile tools. PLoS One 16, e0261813 (2021).
Piqué, R. et al. Characterizing prehistoric archery: Technical and functional analyses of the neolithic bows from la Draga (NE Iberian Peninsula). J. Archaeol. Sci. 55, 166–173 (2015).
Palomo, A. et al. La caza en El yacimiento lacustre de la Draga (Banyoles-Girona). In III Congreso Del Neolítico en la Península Ibérica (Universidad de Cantabria, Santander, 2005).
Dias-Meirinho, M. H. Des Armes et des Hommes. L’archerie à la transition fin du Néolithique/Age du Bronze en Europe occidentale (Université Toulouse le Mirail-Toulouse II, 2011).
Mercadal, O. et al. La Costa De Can Martorell (Dosrius, El Maresme, Barcelona). Muerte Y violencia en una comunidad del litoral catalán durante El tercer milenio cal BC. In III Congreso Del Neolítico en la Península Ibérica (ed Arias Cabal, P., Ontañon Peredo, R. & García Moncó Piñeiro, C.) 671–680 (Universidad de Cantabria, Cantabria, 2005).
Marquez, M., Gibaja, J. F., Gonzalez, J. E., Ibañez, J. J. & Palomo, A. Projectile points as signs of violence in collective burials during the 4th and the 3rd millennia cal. BC in the North-East of the Iberian Peninsula. Prehistoric Technol. 40, 321–326 (2008).
Fernández-Crespo, T. An arrowhead injury in a late Neolithic/Early chalcolithic human cuneiform from the rockshelter of La Peña De Marañón (, Spain). Int. J. Osteoarchaeology 26, 1024–1033 (2016).
Fernández-Crespo, T. et al. Large-scale violence in late neolithic Western Europe based on expanded skeletal evidence from San Juan ante Portam Latinam. Sci. Rep. 13, 17103 (2023).
Santana, J., Rodríguez-Santos, F. J., Camalich-Massieu, M. D., Martín-Socas, D. & Fregel, R. Aggressive or funerary cannibalism? Skull-cup and human bone manipulation in Cueva De El Toro (early neolithic, southern Iberia). Am. J. Phys. Anthropol. 169, 31–54 (2019).
Iversen, R. Arrowheads as indicators of interpersonal violence and group identity among the Neolithic Pitted Ware hunters of southwestern Scandinavia. J. Anthropol. Archaeol. 44, 69–86 (2016).
Mason, O. T. North American Bows, Arrows, and Quivers (JM Carroll., 1893).
Prieto Iglesias, A. Arquería de Tierra Del Fuego. Cuarto Propio. (2011).
Šmit, Ž. Archery by the apaches–implications of using the bow and arrow in hunter-gatherer communities. Doc. Praehistorica 43, 515–525 (2016).
Cetwińska, A. et al. Investigating the potential of using reed (Phragmites) arrowshafts in prehistoric Europe through mechanical and experimental research. Archaeometry. https://doi.org/10.1111/arcm.12967 (2024).
Salem, P. E. & Churchill, S. E. Penetration, tissue damage, and lethality of wood-versus lithic-tipped projectiles. In Multidisciplinary Approaches Study Stone Age Weapon. 203–212 (2016).
Callanan, M. Melting snow patches reveal neolithic archery. Antiquity 87, 728–745 (2013).
Els Objectes De Fusta Del Poblat Neolític De La Draga. Excavacions De 1995–2005 (CASC-Museu d’Arqueologia de Catalunya, Girona, 2006).
Martínez-Sevilla, F. et al. The earliest basketry in southern Europe: Hunter-gatherer and farmer plant-based technology in Cueva De Los Murciélagos (Albuñol). Sci. Adv. 9, eadi3055 (2023).
Jung, C. Insight into protein structure and protein-ligand recognition by Fourier transform infrared spectroscopy. J. Mol. Recognit. 13, 325–351 (2000).
Barth, A. Infrared spectroscopy of proteins. Biochim. Biophys. Acta 1767, 1073–1101 (2007).
Callanan, M. Bronze age arrows from Norwegian alpine snow patches. J. Glacial Archaeol. 1, 25–49 (2014).
Leroy, G. et al. Neolithic wood use at the Scheldt river banks in Bouchain (France). Quat Int. 659, 11–23 (2023).
Hafner, A. Archaeological discoveries on Schnidejoch and at other Ice sites in the European Alps. Arctic 65, 189–202 (2012).
Oeggl, K. The significance of the Tyrolean Iceman for the archaeobotany of Central Europe. Veg. Hist. Archaeobot 18, 1–11 (2009).
Lepers, C. & Rots, V. The important role of bow choice and arrow fletching in projectile experimentation. A ballistic approach. J. Archaeol. Science: Rep. 34, 102613 (2020).
Carrión, J. S. Patterns and processes of late quaternary environmental change in a montane region of southwestern Europe. Quat Sci. Rev. 21, 2047–2066 (2002).
Carrión, J. S. et al. Holocene environmental change in a montane region of southern Europe with a long history of human settlement. Quat Sci. Rev. 26, 1455–1475 (2007).
Anderson, R. S., Jiménez-Moreno, G., Carrión, J. & Pérez-Martinez, C. Postglacial history of alpine vegetation, fire, and climate from Laguna De Río Seco, Sierra Nevada, southern Spain. Quat Sci. Rev. 30, 1615–1629 (2011).
Manzano, S. et al. A palaeoecological approach to understanding the past and present of Sierra Nevada, a Southwestern European biodiversity hotspot. Glob Planet. Change 175, 238–250 (2019).
Oflaz, A., Dörfler, W. & Weinelt, M. An overview of olive trees in the eastern Mediterranean during the mid- to late Holocene: Selective exploitation or established arboriculture? In HOW’S LIFE? Living Conditions in the 2nd and 1st Millennia BCE (eds. Dal Corso, M. et al.) 131–165 (2019).
Carrión Marco, Y., Ntinou, M. & Badal, E. Neolithic woodland in the north Mediterranean basin: A review on Olea europaea L. In Proceedings of the Fourth International Meeting of Anthracology (ed. Damblon, F.) 31–40 (Oxford, Brussels, 2013).
Rodriguez-Ariza, M. O. & Montes, E. M. Origen y domesticación del olivo en Andalucía (España) a partir de los hallazgos arqueológicos de Olea europea L. In I Congreso de la Cultura del Olivo (Diputación Provincial de Jaén, Instituto de Estudios Giennenses, Jaén, 2007) (2005).
Buxó i Capdevila, R. Presence of Olea Europea and Vitis vinífera in archaeological sites from the Iberian Peninsula. Lagascalia 19, 271–282 (1997).
Rodríguez Ariza, M. O. Análisis antracológicos de yacimientos neolíticos de Andalucía. In Actes del I Congreso Del Neolítico en la Península Ibèrica 73–85 (Rubricatum, Barcelona, Gavà, 1996).
Urquijo, J. G. et al., Córdoba) estudio arqueobotánico y de la función del utillaje. 11, 171–190 (2000).
Ayala, M. M. Estudio preliminar del ritual funera- rio calcolítico en la comarca De Lorca. Murcia Zephyrus 37, 77–86 (1990).
Engler, N. Wood and Woodworking Materials (Rodale, 1995).
Hughes, S. S. Getting to the point: Evolutionary change in prehistoric weaponry. J. Archaeol. Method Theory 5, (1998).
Piqué, R. et al. Woody and non-woody forest raw material at the early neolithic site of La Draga (Banyoles, Spain). In The Missing Woodland Resources: Archaeobotanical Studies of the use of Plant raw Materials (eds Berihuete-Azorín, M., Piqué, R. & López-Bultó, O. & Martín Seijo, M.) 37–58 (Barkhuis, Eelde, 2022).
Alix, C., Hare, G., Andrews, T. D. & MacKey, G. A Thousand Years of Lost Hunting arrows: Wood analysis of ice patch remains in Northwestern Canada. Arctic 65, 95–117 (2012).
Vargas, C. A., Nascimento, M. B., de Muñiz, G. I. B. & Nisgoski, S. Physical and mechanical properties of four salix species. Revista Ciência Da Madeira (Brazilian J. Wood Science) 3, https://doi.org/10.12953/2177-6830.v03n02a02 (2012).
Cartwright, C. & Taylor, J. Wooden Egyptian archery bows in the collections of the British Museum. Tech. Res. Bull. 77–83 (2008).
Desmond Clark, J., Phillips, J. L. & Staley, P. S. Interpretations of prehistoric technology from ancient Egyptian and other sources: Part 1: Ancient Egyptian bows and arrows and their relevance for African prehistory. Paléorient 2, 323–388 (1974).
McLeod, W. E. Egyptian composite bows in New York. Am. J. Archaeol. 66, 13–19 (1962).
Lévi-Strauss, C. La Pensée Sauvage (Plon, 1962).
Góngora, M. de. Antigüedades Prehistóricas de Andalucía (Madrid, 1868).
Martínez Sánchez, R. M. et al. The Zacatín rock shelter (Granada, Spain): New data on the mesolithic-neolithic transition in the south-eastern Iberian coast. Quat Int. 680, 36–54 (2024).
Junkmanns, J., Francuz, J., Mischler, K. & Räss, U. Schnidejoch Und Loetschenpass: Bogen, Pfeile und andere Teile Von Bogenausrüstungen. in Schnidejoch Und Lötschenpass. Archäologische Forschungen in den Berner Alpen (eds Hafner, A. et al.) vol. 1 280–319 (Archäologischer Dienst des Kantons Bern, Bern, (2015).
Mcleod, W. Self Bows and Other Archery Tackle from the Tomb of Tut’Ankhamun (Tut’Ankhamun’s Tomb Series IV, 1982).
Pantaleón-Cano, J., Yll, R. & Roure, J. M. Evolución Del paisaje vegetal en El sudeste de la Península Ibérica Durante El Holoceno a partir del análisis polímico. SAGVNTVM Extra 2, 17–23 (1999).
Rageot, M. et al. Birch bark tar production: Experimental and biomolecular approaches to the study of a common and widely used prehistoric adhesive. J. Archaeol. Method Theory 26, 276–312 (2019).
Rageot, M. et al. Management systems of adhesive materials throughout the neolithic in the North-West Mediterranean. J. Archaeol. Sci. 126, (2021).
Kabaciński, J. et al. Expedient and efficient: An early mesolithic composite implement from Krzyż Wielkopolski. Antiquity 97, 295–313 (2023).
Koch, T. J. et al. Chemical analyses reveal dual functionality of early mesolithic birch tar at Krzyż Wielkopolski (Poland). J. Archaeol. Science: Rep. 57, 104591 (2024).
Jiménez-Brobeil, S. A., Botella López, M. C. & Alemán, I. Cráneos trepanados neolíticos de Andalucía Oriental. In Salud, enfermedad y muerte en el pasado. Actas del III Congreso Nacional de Paleopatología (eds. Pérez, A. & Pérez, A.) 271–276 (Fundación Uriach, Barcelona, 1996).
Botella, M. Restos humanos eneolíticos con incisiones de la provincia de Granada. Anales Del. Desarrollo 17, 401–423 (1973).
López-Montalvo, E. Violence in neolithic Iberia: New readings of levantine rock art. Antiquity 89, 309–327 (2015).
Campillo, D., Mercadal, O. & Blanch, R. M. A mortal wound caused by a flint arrowhead in individual MF-18 of the neolithic period exhumed at Sant Quirze Del Valles. Int. J. Osteoarchaeol 3, 145–150 (1993).
Gibaja, J. F. Los Enterramientos neolíticos Del Noreste De La Península Ibérica. Anthropologica et Prœhistorica 116, 165–178 (2005).
Hamilakis, Y. The sacred geography of hunting: Wild animals, social power and gender in early farming societies. Br. School Athens Stud. 9, 239–247 (2003).
López-Montalvo, E. Hunting scenes in Spanish Levantine rock art: An unequivocal chrono-cultural marker of Epipalaeolithic and Mesolithic Iberian societies? Quaternary International (2018).
Millard, A. R. Conventions for reporting radiocarbon determinations. Radiocarbon 56, 555–559 (2014).
Reimer, P. J. et al. The IntCal20 northern hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62, 725–757 (2020).
Bronk Ramsey, C., Higham, T. F. G., Brock, F., Baker, D. & Ditchfield, P. Radiocarbon dates from the oxford AMS system: Archaeometry datelist 33. Archaeometry 51, 323–349 (2009).
Bronk Ramsey, D. Dealing with outliers and offsets in radiocarbon dating. Radiocarbon 51, 1023–1045 (2009).
Dee, M. W. & Bronk Ramsey, C. High-precision Bayesian modeling of samples susceptible to inbuilt age. Radiocarbon 56, 83–94 (2014).
Carr, C. & Maslowski, R. F. Cordage and fabrics. In Style, Society, and Person: Archaeological and Ethnological Perspectives (eds Carr, C. & Neitzel, J. E.) 297–343 (Springer US, Boston, MA, 1995).
Pelegrin, J., Karlin, C. & Bodu, P. Chaînes opératoires: Un outil pour les préhistorien. in Technologie préhistorique 55–62 (Editions du CNRS, Paris, (1988).
Sands, R. Life beyond life: Repair, reuse, and recycle—the many lives of wooden objects and the mutability of trees. Archaeometry 64, 168–186 (2022).
Schweingruber, F. H. European Wood Anatomy (Bern, 1990).
Gale, R. & Cutler, D. Plants in Archaeology. Identification Manual of Vegetative Plant Materials Used in Europe and the Southern Mediterranean to c. 1500 (Westbury and Royal Botanic Gardens., 2000).
Hather, J. G. The Identification of the Northern European Woods. A Guide for Archaeologists and Conservators (Archetype., 2000).
Marguerie, D. & Hunot, J. Y. Charcoal analysis and dendrology: Data from archaeological sites in north-western France. J. Archaeol. Sci. 34, 1417–1433 (2007).
Martín-Seijo, M. The presence of decayed wood in iron age contexts of northwest Iberia: wood-borer galleries and fungal hyphae. Environ. Archaeol. 29, 34–50 (2024).
Monnier, G., Frahm, E., Luo, B. & Missal, K. Developing FTIR Microspectroscopy for the analysis of animal-tissue residues on Stone Tools. J. Archaeol. Method Theory 25, (2018).
Buckley, M., Collins, M., Thomas-Oates, J. & Wilson, J. C. Species identification by analysis of bone collagen using matrix-assisted laser desorption/ionisation time-of-flight mass spectrometry. Rapid Commun. Mass. Spectrom. 23, 3843–3854 (2009).
Strohalm, M., Hassman, M., Kosata, B. & Kodícek, M. mMass data miner: An open source alternative for mass spectrometric data analysis. Rapid Commun. Mass. Spectrom. 22, 905–908 (2008).
Kirby, D. P., Buckley, M., Promise, E., Trauger, S. A. & Holdcraft, T. R. Identification of collagen-based materials in cultural heritage. Analyst 138, 4849–4858 (2013).
Welker, F. et al. Palaeoproteomic evidence identifies archaic hominins associated with the Châtelperronian at the Grotte Du Renne. Proc. Natl. Acad. Sci. 113, 11162–11167 (2016).
Hayek, E. W. H. et al. Identification of archaeological and recent wood tar pitches using gas chromatography/mass spectrometry and pattern recognition. Anal. Chem. 62, 2038–2043 (1990).
Aveling, E. M. & Heron, C. Identification of birch bark tar at the mesolithic site of Star Carr. Anc. Biomol. 2, 69–80 (1998).
Regert, M., Delacotte, J. M., Menu, M., Pétrequin, P. & Rolando, C. Identification of neolithic hafting adhesives from two lake dwellings at Chalain (Jura, France). Anc. Biomol. 2, 81–96 (1998).
Regert, M., Bland, H. A., Dudd, S. N., van Bergen, P. F. & Evershed, R. P. Free and bound fatty acid oxidation products in archaeological ceramic vessels. Proceedings of the Royal Society B-Biological Sciences 265, 2027–2032 (1998).
Schmidt, P. et al. Production method of the Königsaue birch tar documents cumulative culture in neanderthals. Archaeol. Anthropol. Sci. 15, 84 (2023).
Kozowyk, P. R. B., Baron, L. I. & Langejans, G. H. J. Identifying palaeolithic birch tar production techniques: Challenges from an experimental biomolecular approach. Sci. Rep. 13, 1–11 (2023).
Chasan, R., Baron, L. I., Kozowyk, P. R. B. & Langejans, G. H. J. Complicating the debate: Evaluating the potential of gas-chromatography-mass spectrometry for differentiating prehistoric aceramic tar production techniques. J. Archaeol. Sci. 164, 105960 (2024).
Acknowledgements
We are grateful with Eduardo Galán Domingo and Ruth Maicas Ramos curators of the Museo Arqueológico Nacional (Madrid) and the Director of the Museo Arqueológico y Etnográfico (Granada) Manuel Ramos Lizana for permission to study the material and undertake sampling. We also thank the fieldwork team: Antonio Peralta, Blas Ramos Rodríguez, Luis Miguel Zahonero Gómez, Nacho Acosta Gómez, Sofía Elipe Garrido, Cristina Martínez Carrillo, Olmo de Celis Marcelo, Andrea Salamanca Casuso and Francisca Fernández Castaño. Additionally, the authors would like to thank Arnaud Mazuy from CEPAM, as well as Cristina Navarro Senent and Martí de Cabo Jaume from UAB’s Servei de Microscòpia and Servei de Difracció de Raigs-X for their invaluable technical support.This research has been carried out within the framework of different research projects: ‘De los museos al territorio: actualizando el estudio de la Cueva de los Murciélagos de Albuñol (Granada)’ (MUTERMUR) (Referencia CM/JIN/2021-009) financed by the program of Young Researchers of Comunidad de Madrid (directed by F.M.S. as the principal investigator); ‘The organic chemistry and molecular biology of archaeological artefacts’ (ChemArch) funded by the European Union’s Horizon 2020 research and innovation programme under ChemArch grant agreement No 956351 (I.B. as beneficiary); Grant CNS2023-144343 funded by MICIU/AEI /10.13039/501100011033 and European Union NextGenerationEU/PRTR (M.M.S. as principal investigator). J.S. is funded by Ramon y Cajal fellowship (RYC2019-028346) and Consolidación Investigadora (CNS2022-136039) by the Spanish Ministry of Science and Innovation (MCIN). R.P. is ICREA academia researcher and member of the research group TEDAS 2021-SGR 00190 founded by the AGAUR. This work contributes to the EarlyFoods research group (SGR-Cat 2021, 00527).The fieldworks at the site during the years 2022-2023 has had permission from Delegación Territorial de la Junta de Andalucía de Cultura y Patrimonio Histórico de Granada (EXP: BC.03.143/22 14100 and EXP: BC.03.136/23 15490).
Author information
Authors and Affiliations
Contributions
Conceptualization: I.B., M.R., I.T., R.P., F.M.-S- and M-M.-S. Selection of archaeological samples: I.B., M.M.-S, F.M.-S., M.H.-O. and R.P. Methodology: I.B., M.M.-S., K.M., J.S., M.H.-O, M.R., R.P. Funding acquisition : I.B., R.P. and F.M.-S. Writing original draft : I.B., M-M-S., F.M.-S, K.M., J.S., R.P. and I.T. Visualisation : I.B., M.M-S., F.M.-S., J.S., K.M. Writing-review and editing : I.B., M.M-S., F-M.-S., K.M., J.S., M.H.-O., R.M.S., A.P., M.R., I.T., R.P. All authors have reviewed and contributed to the manuscript, and all approve of its submission to this journal.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
41598_2024_77224_MOESM1_ESM.pdf
Supporting information 1: Site description; Datation and Bayesian modelling; Microscopical and SEM photographs of the bowstring; SEM-EDS results; FTIR results; Lipid compounds identified and their relative proportions; ZooMS spectra; Table of neolithic arrowshafts from other archaeological contexts.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Bertin, I., Martín-Seijo, M., Martínez-Sevilla, F. et al. First evidence of early neolithic archery from Cueva de los Murciélagos (Albuñol, Granada) revealed through combined chemical and morphological analysis. Sci Rep 14, 29247 (2024). https://doi.org/10.1038/s41598-024-77224-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-77224-w
