
Abstract
Exploring the components of soil organic carbon (SOC) and aggregate stability across different elevations is crucial to assessing the stability of SOC in subtropical forest ecosystems under climate change. In this study, we investigated the spatial variation of active carbon (C) compositions, aggregate distribution, and stability in Chinese fir (Cunninghamia lanceolata) plantations across an elevation gradient from 750 to 1150 m a.s.l. on the northern foothills of the Dabie Mountains, China. The results showed that macroaggregates accounted for more than 80% of all fractions at different elevations. In the 0–10 cm soil layer, the macroaggregates, mean weight diameter (MWD), geometric mean diameter (GMD), and SOC exhibited a U-shaped distribution trend with increasing elevation. Conversely, in the 10–50 cm soil layer, these indicators showed a consistent increasing trend. Similarly, the contents of easily oxidizable carbon (EOC) and particulate organic carbon (POC) gradually increased with increasing elevation. Microbial biomass carbon (MBC) and silt + clay C exhibited a unimodal distribution pattern along the elevational gradient, peaking at 850 m a.s.l., which is mainly related to soil pH and C/N ratio. Across all elevations, The silt + clay C was significantly higher than that of macro- and micro-aggregate C. Macro- and micro-aggregate C, and dissolved organic carbon (DOC) were significantly positively correlated with MWD. The results demonstrated that elevation and soil layer have significant effects on SOC and aggregate stability. The physical protection of silt + clay fractions and the active carbon pools may be the main mechanisms for organic carbon preservation in the Dabie Mountains. These results contribute to further deepening the impact of elevation on climate change and the C cycling of forest ecosystems.
Similar content being viewed by others
Introduction
Forest soil serves as vast carbon (C) pools in terrestrial ecosystems, depositing about 1500 ± 230 Pg C. It accounted for about 73% of global organic carbon1. Minor fluctuations in forest soil C pools might significantly impact atmospheric CO2 concentration, ultimately affecting global climate dynamics2. Climate warming has become a concern with widespread impacts on global biogeochemical cycles3. Many studies indicate that climate warming accelerates the mineralization and decomposition of organic carbon4,5. However, most field warming experiments have been conducted in high-latitude alpine regions6,7. Little is known about the sensitivity of organic carbon in low-latitude subtropical forests to climate change. Elevation can achieve microclimates and hydrothermal cycling processes by controlling zonal climatic conditions across vertical gradients, making it a suitable natural tool for understanding ecosystem homeostasis under climate change8. Therefore, investigating the stabilization mechanisms of organic carbon in subtropical forests across an elevation gradient is crucial to our understanding of future climate change at global scales.
Soil active C pool comprises various components, each with distinct stability, decomposition rate, and turnover characteristics9. It contains particulate organic carbon (POC), easily oxidizable organic carbon (EOC), dissolved organic carbon (DOC), and microbial biomass carbon (MBC)10. They exhibit diverse responses to climate change and soil fertility11. POC is primarily derived from plant residues and plays a crucial role in aggregate binding and stabilization12. EOC is stemmed from the leaching of plant residue, decomposition of organic matter, and root secretion13. It accounts for a relatively large proportion of SOC, and changes in the soil C pools mainly occur in EOC14. DOC and MBC, as key factors in nutrient turnover, are sensitive to short-term ecosystem changes15. A study revealed that warming increased above- and below-ground biomass, resulting in a 2.3% increase in DOC and a 15.5% rise in light fraction organic carbon (LFOC)16. This underscored the significant impact of environmental factors such as temperature and humidity on the soil C pools15. Elevation is a combined effector, inducing spatial variations in temperature and humidity17. However, the results of studies on the distribution of soil C pools across elevations were inconsistent18. For example, Ma et al.19 showed that SOC and labile organic carbon (LOC) contents of the same vegetation type were significantly higher in high-elevation than in low-elevation, with the contents of MBC, LFOC, and SOC being regulated by temperature, humidity, and elevation. In contrast, in the alpine meadow region of Tibet, China, the concentrations of SOC, DOC, and EOC exhibited a unimodal distribution along the elevation gradient, peaking at 4800 and 4950 m, and subsequently declining. This pattern was attributed to the distribution of aboveground biomass (AGB)20. Dai and Huang21 analyzed the SOC data from different regions of China to detect the relationship between soil organic matter and climate as well as elevation, concluding that there exist regional differences in the main factors influencing soil C pools.
Furthermore, SOC, which serves as the cementing substance for aggregates, enhances their formation and stabilization. The higher its content, the more beneficial it is to soil aggregate stability22. Therefore, the SOC contents in aggregate fractions are often employed as a crucial metric in assessing the sequestration and stability of organic carbon23. Based on aggregate size, it can be categorized into three classes: macroaggregates (> 0.25 mm), microaggregates (0.053–0.25 mm), and silty + clay fractions (< 0.053 mm)24. Their distribution affects the transportation and transformation of SOC25. Notably, the mechanisms of organic carbon stabilization and soil quality varied significantly among different-sized aggregates26. According to Six et al.26, most of the organic carbon was retained in microaggregates and silty + clay fractions, and the physical protection of organic carbon by soil aggregates tended to increase as their size decreased. However, there have also been noted that aggregate C increased when aggregate size increased27. Since soil aggregates physically protect organic carbon from microbial degradation, their stability is a key factor influencing carbon sequestration28. Apart from particle size, climate change can significantly affect aggregate C17. Guan et al.29 performed warming experiments on the Tibetan Plateau. They found that short-term climate warming significantly increased macroaggregate C while decreasing microaggregate C. Zhou et al.30 found that long-term warming-dominated climate change positively affected the DOC, free microaggregate C, and nonaggregated silt + clay C, while negatively affected the EOC, MBC, silt + clay inside microaggregates within macroaggregates C and aggregate stability. Given the diverse sensitivities of organic carbon components to environmental change, it is essential to reveal the response mechanisms of soil aggregate C to climate change in forest ecosystems.
The Dabie Mountains are one of the major mountain ranges in the East China, rich in plant resources. Comprehensive research on the preservation of organic carbon along the elevation gradient in the Dabie Mountains’ ecosystem is relatively scarce. Thus, studying the spatial distribution characteristics of the active carbon pools in the Dabie Mountains ecosystem can aid in understanding the stabilization mechanisms of SOC. As the global temperature rose, significant changes in environmental and spatial patterns such as temperature and precipitation have occurred. These influence plant physiological and ecological adaptations31, thereby altering soil C pools. Chinese fir (Cunninghamia lanceolata), a fine coniferous timber species in southern China, is widely cultivated, with the upper limit of its vertical distribution varying based on topography and climatic conditions. Our study explored the distribution patterns of active C pools and aggregates along an elevation gradient (750–1150 m) in the Chinese fir plantations at the northern foothills of the Dabie Mountain. We aimed to quantify the response of active C pools, aggregate C, and aggregate stability to the elevation gradient. We hypothesized that: (1) due to the decrease of mean annual temperature (MAT), and the increase of mean annual precipitation (MAP) and organic matter inputs, active C pools, macroaggregate, and aggregate stability were higher in high-elevation sites than in low-elevation sites, and (2) physical protection of microaggregates was the main mechanism for organic carbon preservation because of the long-term protective effect of microaggregates on soil organic matter. Overall, this study emphasized the changes in active C pools and aggregate stability with elevation gradient in subtropical forests. It can provide a reference for assessing the impact of global climate change on forest ecosystems.
Materials and methods
Site description
The experimental area is situated in the Mazongling State-owned Forest Farm in Mt. Dabie, Anhui Province, China (31°10′–31°20′ N, 115°31′–115°50′ E), with a total forest area of 3500 ha and an elevation range of 600–1671 m. The area has a variety of landforms such as mountains and hills, as well as valleys and intermountain basins. This area falls under the northern subtropical humid monsoon climate zone, with a mean annual temperature (MAT) of 14.6 ℃ and mean annual precipitation (MAP) of 1510 mm (at the elevation of 680 m a.s.l.), mostly concentrated in May-September. The soils are developed from gneiss-weathered material and are acidic. The soil types are yellow-brown soil at low elevations and mountain yellow soil at high elevations, which are Ferric Alisols and Haplic Alisols according to the FAO soil classification32. The forest types in the area are rich, mainly the north subtropical evergreen and deciduous broad-leaved mixed forest. The dominant tree species are Quercus glauca, Q. myrsinifolia, Castanopsis sclerophylla, and Q. variabilis. There are also a small number of natural forests of Euptelea pleiosperma, Carya dabeishansis, and Cercidiphyllum japonicum. The plantation forests mainly include timber forests such as Chinese fir, Pinus hwangshanensis, and Phyllostachys edulis. In particular, since Chinese fir is an important timber tree species with high economic benefits, a large number of Chinese fir plantations were cultivated from the 1970s to 1980s in the area. In 2012, the area was designated as a national nature reserve, and the Mazongling Forest Farm stopped logging. At present, there are more than 1500 hectares of Chinese fir plantation existing in this area.
Experimental design and soil sampling
In October 2021, according to the growth and distribution of Chinese fir plantations in the area, the plantation stands at four elevations (750 m, 850 m, 1000 m, and 1150 m) that were selected for the study (Table 1). Three standard 20 m × 20 m plots were randomly designed for each elevation. We used a stainless steel soil drill (inner diameter 3 cm) to collect soil samples at 0–10 cm, 10–30 cm, 30–50 cm in each plot. Three subsamples for each plot. A total of 108 soil samples were collected. All soil samples were further divided into 2 parts for analysis. One was used for the determination of active organic carbon components, and another was air-dried and used to determine soil aggregate fractions.
Soil aggregate separation and aggregate stability analysis
Soil aggregate fractions were sieved using the conventional wet-sieving method described33. At first, the soil samples were air-dried at ambient temperature (25 °C), and the other visible materials such as fine roots, plant residues, and gravels, etc. were removed from the soils. It was then sieved using a 5-mm sieve. Finally, 100 g of soil was placed in the 0.053 mm and 0.25 mm sieves in the soil aggregate analyzer for separation. After the separation, each fraction was dried at 60 °C and weighed. The aggregates of > 0.25 mm were classified as macroaggregates, 0.053–0.25 mm aggregates were microaggregates, and < 0.053 mm aggregates were silt + clay fractions.
The mean weight diameter (MWD) and geometric mean diameter (GMD) reflect aggregate stability. The specific calculation formulas are as follows34.
where \({\bar{x}}\)i is the mean diameter of aggregates at level i (mm); wi is the amount of aggregates at level i (%).
Soil properties
Soil pH and EC were measured according to the soil and distilled water ratio (w:v) of 1: 2.5 and 1: 5, respectively. The contents of SOC and TN were determined by an elemental analyzer (EA 3000, Vector, Italy). Particulate organic carbon (POC) was extracted by the sodium hexametaphosphate method33, easily oxidizable organic carbon (EOC) was extracted by KMnO4 oxidation method35, and then POC, EOC, and the SOC contents in the aggregate fractions were determined by using an elemental analyzer. Dissolved organic carbon (DOC) was extracted by K2SO4 leaching and microbial biomass carbon (MBC) was extracted by chloroform fumigation extraction method36 and then determined using a TOC/TN analyzer (Multi N/C 3100, Analytik Jena, Germany).
Statistical analysis
The significance of the differences in the data was tested by the Least Significant Difference (LSD) method (p < 0.05). Two-way ANOVA was used to test the effects of elevation and soil layer on the distribution and stability of aggregates, aggregate C, and active organic carbon components. Before the ANOVA, the homogeneity test of the difference was performed, and if the hypothesis was not met, the data was converted logarithmically. Spearman correlation analysis, principal component analysis (PCA), and regression analysis were used to investigate the relationship between the distribution of aggregates and organic carbon fractions along the elevation gradient. R Studio 4.3.1 and SPSS 22.0 were used to analyze data, Origin 2021 was used for plotting.
Results
Aggregate fractions distribution and stability along an elevation gradient
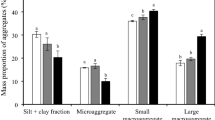
The distributions of soil aggregate fractions at different elevations in Mt. Dabie were significantly different (p < 0.05, Table 2). At the different elevations, macroaggregates (> 0.25 mm) had the highest proportion of 80.33–88.42% in all fractions (Fig. 1).

The distribution of aggregate fractions in Chinese fir plantations at different elevations.
In the 0–10 cm soil layer, the content of macroaggregates showed a “U” distribution with elevation, and the highest proportion was found at 1150 m. However, the microaggregates and silt + clay fractions were in the opposite trend. In the soil layer of 10–50 cm, the macroaggregates showed a gradual increase, while a gradual decrease in the microaggregates and silt + clay fractions with elevation. The results of two-way ANOVA (Table 2) showed that the distribution of soil aggregates was significantly affected by elevation (p < 0.05), and not significantly affected by soil depth and the interaction between elevation and soil depth.
The MWD and GMD of aggregates changed with elevation in a consistent trend, showing a down then up trend in the 0–10 cm soil layer, and a gradual increase in the 10–50 cm soil layer, both of which reached their highest at 1150 m (Fig. 2). Overall, MWD and GMD were significantly higher in high-elevation stands than in low-elevation stands.

The mean weight diameter (MWD) and geometric mean diameter (GMD) of soil aggregate in Chinese fir plantations at different elevations. Different letters on the bars (Means ± S.E., n = 9) indicate significant differences between elevations in the same soil layers (A and B), and between the different soil layers in the same elevations (a and b) (LSD test, p < 0.05).
Distribution of SOC in different aggregate fractions
The SOC contents were significantly affected by elevation and soil depth (p < 0.01, Fig. 3d). SOC content variated from 21.85 to 67.39 g kg−1. In the 0–10 cm soil layer, SOC decreased first and then increased with the increase of elevation. The soil layer of 10–50 cm showed a gradually increasing trend.

The contents of soil organic carbon in Chinese fir plantations at different elevations. Different letters on the bars (Means ± S.E., n = 9) indicate significant differences between elevations in the same soil depth (A and B), and between the different soil depths in the same elevations (a and b) (LSD test, p < 0.05).
The SOC contents in the different aggregate fractions were significantly affected by elevation and soil depth (p < 0.05, Fig. 3a–c). The contents of macroaggregate C ranged from 21.79 to 56.67 g·kg−1, microaggregate C and silt + clay C were 19.76 –51.96 g kg−1 and 29.72–77.19 g kg−1, respectively. The change pattern of C in the macro- and micro-aggregates with elevation was not obvious, and C in the silt + clay showed a unimodal distribution. In general, the SOC contents of aggregate fractions showed a gradual increase with decreasing size, with the highest in silt + clay fractions. In addition, the SOC contents of aggregate fractions decreased gradually (p < 0.05) with soil depth.
Distribution of soil active organic carbon components along an elevation gradient
The elevation gradient significantly affected the soil active organic carbon components (p < 0.05, Figs. 4 and 5). POC was the dominant active organic carbon component at the different elevations with the highest contents varied from 5.46 to 23.42 g kg−1, followed by EOC, MBC, and DOC. The contents of EOC and POC were significantly higher in high-elevation stands than in low-elevation stands (Fig. 8). The contents of MBC showed a unimodal distribution pattern with increasing elevation, while DOC showed a “U”-shaped distribution pattern. In addition, soil depth also significantly affected soil active organic carbon components. EOC, POC, and MBC contents all decreased with deepening soil layers, while DOC appeared no significant change.

The contents of soil active organic carbon components in Chinese fir plantations at different elevations. EOC easily oxidizable organic carbon, POC particulate organic carbon, DOC dissolved organic carbon, MBC microbial biomass carbon. Different letters on the bars (Means ± S.E., n = 9) indicate significant differences between elevations in the same soil layers (A and B), and between the different soil layers in the same elevations (a and b). (LSD test, p < 0.05).

Principal component analysis (PCA) of soil active organic carbon components at different elevations. EOC easily oxidizable organic carbon, POC particulate organic carbon, DOC dissolved organic carbon, MBC microbial biomass carbon.
Relationship between soil aggregates and active organic carbon components and environmental factors
Linear mixed effect model (Fig. 6) showed that MWD was significantly positively correlated with macroaggregates, DOC, macro- and micro-aggregates C, while significantly negatively correlated with microaggregates and silt + clay fractions. In addition, DOC was significantly positively correlated with macro- and micro-aggregates C. MBC was positively correlated with macroaggregate C and silt + clay C (Fig. 7).

Relationships between the MWD and aggregate fractions (a–f) and soil active organic carbon components (g–j). MWD mean weight diameter, LMA macroaggregates, MIA microaggregates, SCA silt + clay fractions, LMA-C macroaggregate C, MIA-C microaggregate C, SCA-C silt + clay C, POC particulate organic carbon, EOC easily oxidizable organic carbon, DOC dissolved organic carbon, MBC microbial biomass carbon. Significant correlations are denoted by *p < 0.05; **p < 0.01; ***p < 0.001.

Relationships between the SOC in different aggregate fractions and soil active organic carbon components (a–l). LMA macroaggregates, MIA microaggregates, SCA silt + clay fractions, LMA-C macroaggregate C, MIA-C microaggregate C, SCA-C silt + clay C, POC particulate organic carbon, EOC easily oxidizable organic carbon, DOC dissolved organic carbon, MBC microbial biomass carbon. Significant correlations are denoted by *p < 0.05; **p < 0.01.
The distribution and stability of aggregates and active C pools along the elevation gradient were closely related to environmental factors (Table 3). The results of Spearman correlation analysis showed that MWD, GMD, macroaggregates, macro- and micro-aggregate C, POC, and EOC were significantly positively correlated with elevation and MAP, and negatively correlated with MAT. However, the opposite was true for the microaggregates and their SOC. Moreover, the SOC in aggregate fractions and active organic C components were significantly positively correlated with SOC and TN. In addition, POC, EOC and MBC were positively correlated with C/N. DOC was significantly negatively correlated with pH. The results indicated environmental factors greatly influenced the aggregate stability.
Discussion
The distribution and stability of soil aggregate are vital indicators for assessing soil structural stability37. It has been shown that the widespread use of indicators such as MWD and GMD in gauging soil aggregate stability, where higher corresponds to a more stable soil aggregate structure38. Our findings reveal a “U”-shaped pattern in the distribution of macroaggregates, MWD, and GMD in the 0–10 cm layer and gradually increased in the 10–50 cm layer as the elevation increased. And all of them reached the peak at 1150 m a.s.l. This suggests that the elevation and soil depth significantly affect aggregate distribution and stability (Figs. 1 and 2). The increase in elevation is often accompanied by changes in environmental factors such as vegetation, temperature, and precipitation. As elevation rises, the MAT gradually decreases, while the MAP increases, which results in low turnover of SOC in the high-elevation. Consequently, aggregate stability was higher in these areas7. Moreover, the canopy cover was higher in high-elevation than in low- and middle-elevation forest stands (Table 1), which favored overall stability. The vegetation cover provided benefits through canopy shading and a surface litter layer, mitigating soil erosion, drastic microclimate changes, and soil water loss. These favorable microclimate conditions and soil moisture supported fine-root growth, turnover, and soil fungal mycelium development. Those changes promoted the development of aggregates29. Some studies have been conducted to show that macroaggregate content at high elevations was associated with higher fungal content and metabolic activity39,40. Our previous study provided evidence that microbial diversity increased with the elevation in the Chinese fir plantations in Mt. Dabie41. The microorganisms regulated the decomposition of organic matter and the production of secondary metabolites, facilitating the transformation of microaggregates into macroaggregates, ultimately enhancing soil aggregate stability19. Bai et al.42 demonstrated that the increased soil moisture significantly promoted the biological process of aggregate formation, strengthening the coupling of aggregates to fungal biomass and exchangeable Mg2+. Furthermore, the input of organic matter is closely related to aggregate stability. In this study, MWD were positively correlated with DOC, macro- and micro-aggregate C (Fig. 6). This underscores the sensitivity of soil aggregate stability to environmental changes and its significant dependence on aggregates C and active C pools. The increased biomass, litterfall, and root exudates along the elevation gradient contribute to increased organic matter inputs, thus promoting aggregate stabilization43.
In forest ecosystems, soils containing a high proportion of macroaggregates are typically regarded as possessing superior soil structure and stability26. Our findings indicate that macroaggregates (> 0.25 mm) accounted for the highest proportion (over 80%) across different elevations (Fig. 1), indicating that the soil aggregates were better structured and stabilized at the study sites. This observation was rationalized with the soil aggregation theory, which posited that macroaggregates are primarily formed by organic matter-derived soil microaggregates and silt + clay fractions, while providing physical protection for microaggregates23. Consequently, the higher the proportion of macroaggregates, the more stable the soil structure becomes. However, the microaggregates and silt + clay fractions decreased with elevation (Fig. 1). Our results showed that MWD were significantly positively correlated with macroaggregates and significantly negatively correlated with microaggregates and silt + clay fractions (Fig. 6). It is consistent with the study conducted by Feyissa et al.44 in the alpine forests and grasslands of the Yulong Snow Mountain in Southwest China. Their study revealed that the proportion of soil macroaggregates was the largest at elevations of 2600 and 3900 m, with 50.54% and 49.11%, respectively, contributing significantly to the total soil C content. Kong et al.45 found that the proportion of 0.25–1.00 mm and 0.053–0.25 mm aggregates decreased significantly with elevation. Notably, the response of macroaggregates is susceptible to external environmental factors46, especially in the topsoil layer.
SOC primarily originated from plant litterfall, roots, animal and microbial residues23. The SOC contents in the different aggregate fractions represented a microscopic characterization of soil organic matter formation and decomposition. Our study found that the SOC content of silt + clay fraction appeared a unimodal pattern along the elevation gradient, peaking at 850 m. In the low- and middle-elevation Chinese fir plantations, the higher soil C/N ratio in the silt + clay fraction compared to the macro- and micro-aggregates indicated an increased accumulation of refractory organic matter within aggregate fractions (Table 1), slower SOC turnover, and differences in microbial community habitats7. It may also be due to differences in microbial biomass and microbial residue accumulation (Figs. 4 and 6)47. In addition, we observed that the SOC content was significantly higher in silt + clay fraction than in macro- and micro-aggregates across different elevations (Fig. 3). This finding highlights the crucial role of silt + clay fraction in organic carbon preservation in soil ecosystems. According to previous studies, it is known that macroaggregates are mainly formed by litterfall residues, microaggregates are formed inside the macroaggregates, and the silt + clay fractions are associated with mineral-associated organic carbon (MAOC)48,49. The input of plant C leads to the decomposition of organic matter. Roots, hyphae, and polysaccharides then preferently bind to mineral particles to form silt-clay-organic composites50. This suggests that physical protection in the silt + clay fractions was the main mechanism for organic carbon preservation26. The contribution of SOC in the silt + clay fractions to organic carbon preservation was relatively greater, which was confirmed by the findings of Virto et al.51. In contrast, many studies showed that the macroaggregate C was significantly higher than that of microaggregates and silt + clay fractions27,52. It may be more in line with the hierarchy theory of aggregates53. However, the macroaggregate C was mainly derived from humus, making it more easily mineralized and less stable than microaggregates24. This implies a high risk of organic carbon loss from soil macroaggregates under a warming climate, especially in subtropical forested areas54. Therefore, understanding the dynamics and stability of different soil aggregate fractions is crucial for effective carbon sequestration strategies in vulnerable ecosystems.
Soil active organic C is not only related to land use practices and vegetation types, but also to the combined effects of climatic factors such as temperature and moisture30,46. In general, elevation indirectly modulates temperature and moisture, thereby affecting the rate of OC mineralization to some extent. Some studies demonstrated that the contents of EOC, POC, and DOC respond significantly to increasing elevation, exhibiting an upward trend19,55. Our study revealed that the contents of EOC and POC were significantly higher at high-elevation than at low-elevation stands (Fig. 4). The results support this by showing that the decrease in soil temperature and increase in humidity with the rising elevation, resulted in slow mineralization and decomposition of soil organic matter and an increase of SOC accumulation. Conversely, SOC accumulation was lower at the lower elevations. Previous studies have shown that elevation can affect soil LOC by regulating soil hydrothermal8,30. Fang et al.16 similarly demonstrated that warming treatments led to substantial decreases in the contents of POC and MBC in low-elevation forests, highlighting the effect of temperature change on soil organic carbon in mountain forests. It can be inferred that the organic carbon turnover occurred more rapidly at the low elevation than at the high elevation. On the other hand, Kong et al.45 demonstrated that changes in organic carbon fractions at high-elevation may be closely related to macroaggregate C. These results are similar to our findings. MBC, the C contained in the microbial biomass, depends on the abundance of soil microorganisms15. DOC primarily originates from the soil microbial-driven decomposition of organic matter, and its content depends on the degree of microbial degradation of organic matter56. Our results showed that MBC appeared a unimodal pattern and DOC was a “U” shape distribution (Fig. 8) that could associated with higher pH and C/N ratio at mid-elevations (850 m and 1000 m)57. In this study, pH increased first and then decreased with increasing elevation (Table 1). As elevation rises, temperature decreases, leading to a slowdown in organic carbon mineralization and a gradual increase in pH, which enhances microbial activity. The highest soil C/N ratio at mid-elevations are largely attributed to the slowed decomposition of organic matter, directly influencing the accumulation of DOC and MBC. Furthermore, our study found a decrease in the SOC contents of active carbon components and aggregate fractions as soil depth increased (Figs. 3 and 4). This trend can be explained by the decreasing inputs of litterfall, reduced root exudates, and diminished microbial activities within the soil profile44,58.

Linear regressions of the EOC (a) and POC (b), and quadratic regressions of the DOC (c) and MBC (d) versus Elevations. R values are shown for all regressions; the significance of regressions is reported.
Conclusions
In summary, our study revealed the significant influence of different soil depth and elevation on soil properties across the vertical soil profile. Specifically, in the 0–10 cm soil layer, MWD, GMD, and macroaggregates content all exhibit a U-shaped distribution trend with the increase of elevation. Conversely, in the 10–50 cm soil layer, these indicators demonstrate a consistent trend across the elevation gradient. Furthermore, the contents of EOC and POC steadily increase with the rise in elevation, indicating more pronounced accumulation of soil organic matter in high-elevation sites. Notably, MBC and silt + clay C in 0–50 cm soil layers display a unimodal distribution pattern along the elevation gradient, peaking at 850 m a.s.l. The silt + clay C contents were significantly higher than macroaggregate C and microaggregate C along the elevation gradient. Our finding offers a novel perspective on understanding C sequestration mechanisms under elevational gradients. The contents of EOC, POC, MBC, and aggregate organic carbon all decreased gradually with soil depth. Interestingly, the MWD and GMD were significantly positively correlated with DOC, macroaggregates, and macroaggregate- and microaggregate-C, while negatively correlated with microaggregates and silt + clay fractions. This suggests that SOC content has a strong influence on aggregate stability. The results indicated a close relationship among aggregate, active organic C components, and environmental factors. Physical protection of aggregates and organic carbon accumulation may be the main mechanisms for SOC preservation along an elevation gradient in the Dabie Mountain. It is essential to further study organic carbon sequestration under climate change and thus accurately predict the response of organic carbon stability to climate warming.
Data availability
All data generated or analyzed during this study are included in this published article.
References
Schlesinger, W. H. J. N. Evidence from chronosequence studies for a low carbon-storage potential of soils. Nature 348, 232–234. https://doi.org/10.1038/348232a0 (1990).
Wang, M. et al. Global soil profiles indicate depth-dependent soil carbon losses under a warmer climate. Nat. Commun. 13, 5514. https://doi.org/10.1038/s41467-022-33278-w (2022).
Kay, J. E. Early climate models successfully predicted global warming. Nature 578, 45–46. https://doi.org/10.1038/d41586-020-00243-w (2020).
Xu, W. F., Yuan, W. P., Cui, L. L., Ma, M. N. & Zhang, F. G. Responses of soil organic carbon decomposition to warming depend on the natural warming gradient. Geoderma 343, 10–18. https://doi.org/10.1016/j.geoderma.2019.02.017 (2019).
Qiu, Y. P. et al. Warming and elevated ozone induce tradeoffs between fine roots and mycorrhizal fungi and stimulate organic carbon decomposition. Sci. Adv. 7, eabe9256. https://doi.org/10.1126/sciadv.abe9256 (2021).
Che, M. et al. Effects of elevation and slope aspect on the distribution of the soil organic carbon associated with Al and Fe mineral phases in alpine shrub-meadow soil. Sci. Total Environ. 753, 141933. https://doi.org/10.1016/j.scitotenv.2020.141933 (2021).
Murugan, R. et al. Spatial distribution of microbial biomass and residues across soil aggregate fractions at different elevations in the Central Austrian Alps. Geoderma 339, 1–8. https://doi.org/10.1016/j.geoderma.2018.12.018 (2019).
Puissant, J. et al. Climate change effects on the stability and chemistry of soil organic carbon pools in a subalpine grassland. Biogeochemistry 132, 123–139. https://doi.org/10.1007/s10533-016-0291-8 (2017).
Li, N., Long, J., Han, X., Yuan, Y. & Sheng, M. Molecular characterization of soil organic carbon in water-stable aggregate fractions during the early pedogenesis from parent material of Mollisols. J. Soils Sediments 20, 1869–1880. https://doi.org/10.1007/s11368-020-02563-w (2020).
Plaza-Bonilla, D., Alvaro-Fuentes, J. & Cantero-Martinez, C. Identifying soil organic carbon fractions sensitive to agricultural management practices. Soil Till. Res. 139, 19–22. https://doi.org/10.1016/j.still.2014.01.006 (2014).
Benbi, D. K., Brar, K., Toor, A. S. & Sharma, S. Sensitivity of labile soil organic carbon pools to long-term fertilizer, straw and manure management in rice-wheat system. Pedosphere 25, 534–545. https://doi.org/10.1016/s1002-0160(15)30034-5 (2015).
Bian, Q. et al. Divergent accumulation of microbe- and plant-derived carbon in different soil organic matter fractions in paddy soils under long-term organic amendments. Agric. Ecosyst. Environ. 366, 108934. https://doi.org/10.1016/j.agee.2024.108934 (2024).
Liu, X. J. et al. Aggregate-associated soil organic carbon fractions in subtropical soil undergoing vegetative restoration. Land Degrad. Dev. 34, 4296–4306. https://doi.org/10.1002/ldr.4777 (2023).
Conteh, A., Blair, G., Lefroy, R. & Whitbread, A. Labile organic carbon determined by permanganate oxidation and its relationships to other measurements of soil organic carbon. Humic Subst. Environ. 1, 3–15 (1999).
Yang, Z. Y. et al. Soil carbon storage and accessibility drive microbial carbon use efficiency by regulating microbial diversity and key taxa in intercropping ecosystems. Biol. Fertil. Soils. 60, 437–453. https://doi.org/10.1007/s00374-024-01804-1 (2024).
Fang, X. et al. Translocating subtropical forest soils to a warmer region alters microbial communities and increases the decomposition of mineral-associated organic carbon. Soil Biol. Biochem. 142, 107707. https://doi.org/10.1016/j.soilbio.2020.107707 (2020).
Siles, J. A., Cajthaml, T., Filipová, A., Minerbi, S. & Margesin, R. Altitudinal, seasonal and interannual shifts in microbial communities and chemical composition of soil organic matter in Alpine forest soils. Soil Biol. Biochem. 112, 1–13. https://doi.org/10.1016/j.soilbio.2017.04.014 (2017).
Liu, W. et al. Spatial distribution and stability mechanisms of soil organic carbon in a tropical montane rainforest. Ecol. Indic. 129, 107965. https://doi.org/10.1016/j.ecolind.2021.107965 (2021).
Ma, H., Yang, X., Guo, Q., Zhang, X. & Zhou, C.-N. Soil organic carbon pool along different altitudinal level in the Sygera Mountains, Tibetan Plateau. J. Mt. Sci. 13, 476–483. https://doi.org/10.1007/s11629-014-3421-6 (2016).
Li, C. et al. Elevational gradient affect functional fractions of soil organic carbon and aggregates stability in a tibetan alpine meadow. Catena 156, 139–148. https://doi.org/10.1016/j.catena.2017.04.007 (2017).
Dai, W. H. & Huang, Y. Relation of soil organic matter concentration to climate and altitude in zonal soils of China. Catena 65, 87–94. https://doi.org/10.1016/j.catena.2005.10.006 (2006).
Piazza, G., Pellegrino, E., Moscatelli, M. C. & Ercoli, L. Long-term conservation tillage and nitrogen fertilization effects on soil aggregate distribution, nutrient stocks and enzymatic activities in bulk soil and occluded microaggregates. Soil Till. Res. 196, 104482. https://doi.org/10.1016/j.still.2019.104482 (2020).
Bronick, C. J. & Lal, R. Soil structure and management: a review. Geoderma 124, 3–22. https://doi.org/10.1016/j.geoderma.2004.03.005 (2005).
Six, J., Elliott, E. T. & Paustian, K. Soil macroaggregate turnover and microaggregate formation: a mechanism for C sequestration under no-tillage agriculture. Soil Biol. Biochem. 32, 2099–2103. https://doi.org/10.1016/s0038-0717(00)00179-6 (2000).
Liu, D. et al. Associated soil aggregate nutrients and controlling factors on aggregate stability in semiarid grassland under different grazing prohibition timeframes. Sci. Total Environ. 777, 146104. https://doi.org/10.1016/j.scitotenv.2021.146104 (2021).
Six, J., Conant, R. T., Paul, E. A. & Paustian, K. Stabilization mechanisms of soil organic matter: implications for C-saturation of soils. Plant Soil 241, 155–176. https://doi.org/10.1023/a:1016125726789 (2002).
Zhang, Y. et al. Effects of farmland conversion on the stoichiometry of carbon, nitrogen, and phosphorus in soil aggregates on the Loess Plateau of China. Geoderma 351, 188–196. https://doi.org/10.1016/j.geoderma.2019.05.037 (2019).
Smith, A. P., Marin-Spiotta, E., de Graaff, M. A. & Balser, T. C. Microbial community structure varies across soil organic matter aggregate pools during tropical land cover change. Soil Biol. Biochem. 77, 292–303. https://doi.org/10.1016/j.soilbio.2014.05.030 (2014).
Guan, S. et al. Climate warming impacts on soil organic carbon fractions and aggregate stability in a tibetan alpine meadow. Soil Biol. Biochem. 116, 224–236. https://doi.org/10.1016/j.soilbio.2017.10.011 (2018).
Zhou, M. et al. Warming-dominated climate change impacts on soil organic carbon fractions and aggregate stability in Mollisols. Geoderma 438, 116618. https://doi.org/10.1016/j.geoderma.2023.116618 (2023).
Paudel, I., Rose, K. M. E., Landhausser, S. M. & Jacobs, D. F. Co-limitation of resources reveals adaptations of a tropical tree to heterogeneous environments along an elevational cline. Front. Glob. Change 6, 1225778. https://doi.org/10.3389/ffgc.2023.1225778 (2023).
FAO. Guidelines for soil Description 4th edn (FAO/ISRIC, 2006).
Cambardella, C. & Elliott, E. Methods for physical separation and characterization of soil organic matter fractions. In Soil Structure/Soil Biota Interrelationships 449–457. https://doi.org/10.1016/B978-0-444-81490-6.50036-4 (1993).
Kemper, W. & Rosenau, R. J. M. Aggregate stability and size distribution. 5, 425–442. https://doi.org/10.2136/sssabookser5.1.2ed.c17 (1986).
Blair, G. J., Lefroy, R. D. B. & Lisle, L. J. A. Soil carbon fractions based on their degree of oxidation, and the development of a carbon management index for agricultural systems. Aust. J. Agric. Res. 46, 1459–1466. https://doi.org/10.1071/AR9951459 (1995).
Brookes, P., Landman, A., Pruden, G. & Jenkinson, D. S. Chloroform fumigation and the release of soil nitrogen: a rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 17, 837–842. https://doi.org/10.1016/0038-0717(85)90144-0 (1985).
Regelink, I. C. et al. Linkages between aggregate formation, porosity and soil chemical properties. Geoderma 247–248. https://doi.org/10.1016/j.geoderma.2015.01.022 (2015).
Chen, Y., Day, S. D., Wick, A. F. & McGuire, K. J. Influence of urban land development and subsequent soil rehabilitation on soil aggregates, carbon, and hydraulic conductivity. Sci. Total Environ. 494, 329–336. https://doi.org/10.1016/j.scitotenv.2014.06.099 (2014).
Hernandez-Caceres, D. et al. Vegetation creates microenvironments that influence soil microbial activity and functional diversity along an elevation gradient. Soil Biol. Biochem. 165, 108485. https://doi.org/10.1016/j.soilbio.2021.108485 (2022).
Wu, M. et al. Aggregate-associated carbon contributes to soil organic carbon accumulation along the elevation gradient of Helan Mountains. Soil Biol. Biochem. 178, 108926. https://doi.org/10.1016/j.soilbio.2022.108926 (2023).
Fan, W. Soil microbial community characteristics and their driving mechanisms of the first- and second-generation Chinese fir plantations across an altitude gradient at Mazongling in Anhui. https://doi.org/10.26919/d.cnki.gannu (2023).
Bai, T. et al. Interactive global change factors mitigate soil aggregation and carbon change in a semi-arid grassland. Glob. Change Biol. 26, 5320–5332. https://doi.org/10.1111/gcb.15220 (2020).
Sun, S. et al. Soil warming and nitrogen deposition alter soil respiration, microbial community structure and organic carbon composition in a coniferous forest on eastern tibetan Plateau. Geoderma 353, 283–292. https://doi.org/10.1016/j.geoderma.2019.07.023 (2019).
Feyissa, A., Raza, S. T. & Cheng, X. L. Soil carbon stabilization and potential stabilizing mechanisms along elevational gradients in alpine forest and grassland ecosystems of Southwest China. Catena 229, 107210. https://doi.org/10.1016/j.catena.2023.107210 (2023).
Kong, J. et al. Elevational gradients and distributions of aggregate associated organic carbon and nitrogen and stability in alpine forest ecosystems. Soil Sci. Soc. Am. J. 84, 1971–1982. https://doi.org/10.1002/saj2.20121 (2020).
Zhang, J. et al. Impacts of long-term fertilization on the molecular structure of humic acid and organic carbon content in soil aggregates in black soil. Sci. Rep. 9, 11908. https://doi.org/10.1038/s41598-019-48406-8 (2019).
Six, J., Frey, S. D., Thiet, R. K. & Batten, K. Bacterial and fungal contributions to carbon sequestration in agroecosystems. Soil Sci. Soc. Am. J. 70, 555–569 (2006).
Angst, G. et al. Unlocking complex soil systems as carbon sinks: multi-pool management as the key. Nat. Commun. 14, 2967. https://doi.org/10.1038/s41467-023-38700-5 (2023).
Peng, X. Y., Huang, Y., Duan, X. W., Yang, H. & Liu, J. X. Particulate and mineral-associated organic carbon fractions reveal the roles of soil aggregates under different land-use types in a karst faulted basin of China. Catena 220, 106721. https://doi.org/10.1016/j.catena.2022.106721 (2023).
Silveira, M. L., Xu, S. T., Adewopo, J., Franzluebbers, A. J. & Buonadio, G. Grazing land intensification effects on soil C dynamics in aggregate size fractions of a spodosol. Geoderma 230, 185–193. https://doi.org/10.1016/j.geoderma.2014.04.012 (2014).
Virto, I., Barre, P. & Chenu, C. Microaggregation and organic matter storage at the silt-size scale. Geoderma 146, 326–335. https://doi.org/10.1016/j.geoderma.2008.05.021 (2008).
Okolo, C. C., Gebresamuel, G., Zenebe, A., Haile, M. & Eze, P. N. Accumulation of organic carbon in various soil aggregate sizes under different land use systems in a semi-arid environment. Agric. Ecosyst. Environ. 297, 106924. https://doi.org/10.1016/j.agee.2020.106924 (2020).
Six, J., Bossuyt, H., Degryze, S. & Denef, K. A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil Till. Res. 79, 7–31. https://doi.org/10.1016/j.still.2004.03.008 (2004).
Yang, H., Wang, J. & Zhang, F. Soil aggregation and aggregate-associated carbon under four typical halophyte communities in an arid area. Environ. Sci. Pollut. Res. 23, 23920–23929. https://doi.org/10.1007/s11356-016-7583-3 (2016).
Zhao, M. et al. Variations in soil microbial communities in the sedge-dominated peatlands along an altitude gradient on the northern slope of Changbai Mountain, China. Ecol. Indic. 129, 107964. https://doi.org/10.1016/j.ecolind.2021.107964 (2021).
Dou, W., Xiao, B., Revillini, D. & Delgado-Baquerizo, M. Biocrusts enhance soil organic carbon stability and regulate the fate of new-input carbon in semiarid desert ecosystems. Sci. Total Environ. 918, 170794. https://doi.org/10.1016/j.scitotenv.2024.170794 (2024).
Dai, Z. et al. Metagenomic insights into soil microbial communities involved in carbon cycling along an elevation climosequences. Environ. Microbiol. 23, 4631–4645 (2021).
Chen, L. et al. Soil carbon persistence governed by plant input and mineral protection at regional and global scales. Ecol. Lett. 24, 1018–1028. https://doi.org/10.1111/ele.13723 (2021).
Acknowledgements
We thank Pengfei Deng, Ruoyong Yin, Zitao Du, and Jiaqi Wang for their valuable efforts in establishing the research plots and helping in soil sampling. Funding for this experiment was provided by the National Key Research and Development Program of China (Grant No. 2016YFD0600304-03).
Author information
Authors and Affiliations
Contributions
X.C.: data curation, formal analysis, methodology, software, visualization, writing-original draft. Y.X.: data curation, investigation, methodology. F.W.: investigation. Z.Z.: investigation. X.X.: conceptualization, funding acquisition, methodology, project administration, supervision, writing-review & editing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Cao, X., Xu, Y., Wang, F. et al. Changes of soil organic carbon and aggregate stability along elevation gradient in Cunninghamia lanceolata plantations. Sci Rep 14, 31778 (2024). https://doi.org/10.1038/s41598-024-81847-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-81847-4
