
Abstract
The cochlear nuclear complex (CN), the starting point for all central auditory processing, encompasses a suite of neuronal cell types highly specialized for neural coding of acoustic signals. However, the molecular logic governing these specializations remains unknown. By combining single-nucleus RNA sequencing and Patch-seq analysis, we reveal a set of transcriptionally distinct cell populations encompassing all previously observed types and discover multiple hitherto unknown subtypes with anatomical and physiological identity. The resulting comprehensive cell-type taxonomy reconciles anatomical position, morphological, physiological, and molecular criteria, enabling the determination of the molecular basis of the specialized cellular phenotypes in the CN. In particular, CN cell-type identity is encoded in a transcriptional architecture that orchestrates functionally congruent expression across a small set of gene families to customize projection patterns, input-output synaptic communication, and biophysical features required for encoding distinct aspects of acoustic signals. This high-resolution account of cellular heterogeneity from the molecular to the circuit level reveals the molecular logic driving cellular specializations, thus enabling the genetic dissection of auditory processing and hearing disorders with a high specificity.
Similar content being viewed by others

Introduction
Information processing along parallel pathways is an important feature of most biological sensory systems. In the mammalian auditory system, auditory signals are encoded by cochlear hair cells and transmitted to the first processing station in the brain, the cochlear nuclear complex (CN), where all higher-level parallel pathways are initiated1. Neurons in the CN are organized tonotopically into precise parallel sensory maps and each is highly specialized for processing different aspects of acoustic information including sound ___location, intensity, frequency content, and spectrotemporal modulations2,3. Classical studies, using histology, in vivo and in vitro recording, and pathway tracing, have classified CN projection neurons into at least four types: the bushy, T-stellate, fusiform, and octopus cells, which each carry distinct ascending signals needed for high-level sound processing4,5,6. In addition, multiple types of interneurons within CN determine the pattern and magnitude of signals that ascend through the neuraxis6. These cell types in CN have been extensively characterized and clearly defined based on their unique morpho-physiological properties, synaptic connectivity, projection targets, and functions4,5,6. We term these well-defined projection neurons and interneurons as established cell types (Fig. 1a). Despite their remarkable phenotypic differences, molecular distinctions and mechanisms that specify and distinguish these cell types are still largely unknown. Since most of these cell types remain molecularly inaccessible, the exact contribution of any given cell type in CN to sound representations in higher brain regions is still unknown. In addition, numerous lines of evidence suggest that the established cell types do not capture the true range of cellular heterogeneity and their functions. For example, in vivo recordings document diverse patterns of acoustic response within a given type of neuron, suggesting that existing definitions of cell types do not have sufficient resolution to account for the variety of downstream anatomical projections and physiological responses7,8,9. In addition, recent in vitro physiological studies using Cre mouse lines and optogenetics have suggested the existence of previously undescribed cell types in CN10,11. Therefore, a full spectrum of cell types and their functions in CN have yet to be established, preventing a full understanding of central auditory processing.

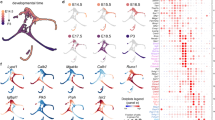
a Anatomy of mouse cochlear nucleus (CN; Light blue: VCN. Dark blue: DCN) and depiction of neuronal cell types (cartoon drawings) across the CN (sagittal view), colored by cell type identity. D dorsal, P posterior, M medial. The 3D brain model was generated with Brainrender (Ver. 2.1.10). b UMAP visualization of 31,639 nuclei from CN neurons clustered by over-dispersed genes, colored by cluster identity. Each cluster labeled by key marker genes. c Right: dendrogram indicating hierarchical relationships between clusters. Left: dot plot of scaled expression of selected marker genes for each cluster shown in (b); see Supplementary Data 1. Circle size depicts percentage of cells in the cluster in which the marker was detected (≥1 UMI), and color depicts the normalized average transcript count in expressing cells. See Figs. S1–3 for more technical details and annotations.
Single-cell transcriptomics offers a powerful, unbiased approach to profile cellular and molecular heterogeneity and define cell types through gene expression profiling of individual cells12,13,14. To capture the full spectrum of cellular heterogeneity and their functions in CN, we employed single-nucleus RNA sequencing (snRNA-seq) to profile the transcriptome of the whole CN at the single-cell level and generate a comprehensive taxonomy of transcriptomic cell types. To relate each transcriptionally distinct cell population to the established CN cell types with functional relevance, we performed parallel Patch-seq experiments that link each cell’s transcriptome to their morphological and physiological features15, followed by the validation using fluorescent in situ hybridization (FISH) and transgenic mouse lines. These approaches allow us to establish a clear correspondence between transcriptionally distinct cell types and all previously established CN cell types. Importantly, we also discover multiple subtypes of major projection neurons based on transcriptomic profiling that reflect hitherto unresolved distinctions in anatomy, morphology, and physiology of cell types16,17, suggesting the existence of previously undescribed auditory processing pathways in the mammalian brain. Our analyses of single-cell transcriptomes identified specific molecular markers for CN cell types and illustrated potential molecular and gene regulatory mechanisms that specify cell type identity, connectivity and projection patterns, and remarkable synaptic and biophysical features, providing molecular delineations of parallel auditory pathways at their initiation.
Results
Identification of transcriptionally distinct cell populations in mouse CN
To investigate transcriptional and cellular heterogeneity within the CN, we performed RNA sequencing on individual nuclei isolated from CN of male and female C57BL/6 mice (postnatal day (P) 22–28) (Fig. S1a). Our final dataset contained 61,211 CN nuclei (‘‘Methods’’ and Figs. S2, 3), and analysis of sequence reads revealed ~4000 uniquely detected transcripts from ~1900 genes for each nucleus. Following data integration, unsupervised dimensionality reduction techniques identified 14 major transcriptionally distinct cell populations (Fig. S1b). By inspection of known markers, we identified all non-neuronal cell types in CN (Fig. S1c and Supplementary Data 1). Using the pan-neuronal marker Snap25, we identified 31,639 neuronal nuclei expressing either the gene for vesicular glutamate transporters Slc17a7 or Slc17a6 or the gene for the glycine transporter Slc6a5, thereby separating glutamatergic and glycinergic populations (Fig. S1b, c). Clustering analysis of neuronal nuclei resulted in 13 molecularly distinct populations (Fig. 1b) including seven glutamatergic/excitatory clusters (Slc17a7+ and/or Slc17a6+) and six glycinergic/inhibitory clusters (Slc6a5+, Fig. 1c).
Based on known markers, we assigned three clusters to known CN cell types. Two glutamatergic clusters that expressed only Slc17a7 (not Slc17a6) were either enriched with Gabra6, a marker gene for granule cells (GCs)18,19, or with Tbr2/Eomes and Samd3, gene markers for unipolar brush cells (UBCs)20,21, thus corresponding to GCs and UBCs in CN, respectively (Fig. 1b, c, Fig. S1d). The UBC cluster was further validated with RNA FISH staining for Samd3 (Fig. S1d) and characterized by graded and inversely correlated expression of glutamate excitatory signaling pathways and inhibitory pathways, reminiscent of UBCs in cerebellum (Fig. S1e)21,22,23. One glycinergic cluster was enriched with Grm2 (Fig. 1c), a marker gene for Golgi cells in CN (in addition to UBCs)24,25, thus corresponding to auditory Golgi cells (Fig. 1b, c). Differentially expressed gene (DEG) analysis identified Cacng3 and Tfap2b as more specific marker genes for auditory Golgi cells (Fig. 1b, c), which were validated with FISH (Fig. S1f). Other major clusters are unknown in terms of their potential correspondence to well-studied CN cell types (Fig. 1a) but could be labeled by one or two candidate gene markers identified via DEG analysis (Fig. 1b, c and Supplementary Data 1).
Molecular profiles of CN excitatory neurons
To relate the unknown clusters to known CN cell types, we performed Patch-seq, starting with four major excitatory projection neurons, bushy, T-stellate, octopus, and fusiform cells4, targeting each by their known anatomical ___location aided by transgenic mouse lines (see ‘‘Methods’’). Membrane potential responses to current injections were recorded, cytosol and nucleus mRNA were extracted and reverse transcribed, and the resulting cDNA was sequenced (Fig. S3g–k, ‘‘Methods’’). The morphology of each neuron was post hoc recovered and reconstructed. Based on their stereotypical morphological and electrophysiological properties, each neuron could be readily identified and assigned to a type, a process we term “expert classification” (“Methods”; Figs. 1a, 2a, b, Supplementary Data 2)26. To validate expert classification and support cell-type assignment of each Patch-seq neuron, we used a random forest classifier (“Methods”; Figs. 2c, d and S4a) or clustering-based k-means classifier (“Methods”; Fig. S4b) based on either electrophysiology or morphology to distinguish between any two types of CN excitatory neurons as labeled manually, resulting in successful separation of almost all cell type pairs, thereby supporting our expert classification. UMAP projection of these Patch-seq cells resulted in five transcriptomic clusters, and cells labeled as the same type clustered together and were rarely confused with another cluster (Fig. 2e), except for bushy cells, which fell into two prominent clusters. Using the gene expression profiles, we mapped these Patch-seq neurons to the transcriptomic clusters identified via snRNA-seq (Fig. 1b)27,28, demonstrating close correspondence in the two datasets and identifying transcriptomic clusters for the established excitatory cell types (“Methods”, Fig. 2f, g). Importantly, those Patch-seq bushy cells clustering together as Cluster 1 in Patch-seq UMAP space (Fig. 2e) were all mapped to the Hhip+ cluster (Fig. 2f), while almost all bushy cells in Cluster 2 (Fig. 2e) were mapped to the Atoh7+ cluster (Fig. 2f). Thus, our analysis not only reveals a strong congruence across three modalities in excitatory neuron classification, but also molecular heterogeneity among bushy cells (see below).

a Examples of reconstructed excitatory neurons positioned at their site of origin in representative mouse CN (sagittal view), colored by cell type identity. b Example responses of CN excitatory cell types to current steps. Bottom traces show the injected currents. Scalebar: 100 mV for potential, 2 nA for injected currents for all cells except octopus cells (10 nA). c Left: UMAP visualization of physiological features of 404 CN excitatory neurons (E-cluster), colored by expert cell type. n = 243 for bushy, n = 67 for T-stellate, n = 58 for octopus, n = 36 for fusiform. Right: Confusion matrix shows performance (an average of 99.2% accuracy) of the cell-type classifier trained with physiological features. See Fig. S4 for more details. d Left: UMAP visualization of morphological features of 157 CN excitatory neurons (M-cluster), colored by expert cell type. n = 105 for bushy, n = 16 for T-stellate, n = 10 for octopus, n = 26 for fusiform. Right: Confusion matrix shows performance (an average of 95.5% accuracy) of the classifier trained with morphological features. See Fig. S4 for more details. e Left: UMAP visualization of 293 Patch-seq excitatory neurons clustered by over-dispersed genes, colored by transcriptomic cluster (T-cluster). Right: The matrix shows proportion of Patch-seq cells assigned to a specific cell type (the expert classification) among each T-cluster. All cells in T-cluster 1 (n = 67/67) and almost all cells in T-cluster 2 (n = 111/113) are bushy cells; all cells in T-cluster 3 are fusiform cells (n = 22/22); all cells in T-cluster 4 are octopus cells (n = 43/43); almost all cells in T-cluster 5 are T-stellate cells (n = 46/48). f Left: Patch-seq neurons (n = 293) mapped to snRNA-seq UMAP space as shown in Fig. 1b. Individual dots represent each cell, colored by T-cluster. Right: Matrix shows proportion of Patch-seq cells assigned to each T-cluster in (e) mapped to one of 13 clusters in UMAP space. g UMAP visualization and annotation of molecular cell types with established morpho-electrophysiological types in CN. Source data are provided in the Source Data file.
DEG analysis was performed to find marker genes for each cell type (or cluster, Supplementary Data 1), while FISH and immunohistochemistry validated their expression and cell type-specificity by leveraging the known anatomical ___location of each cell type (‘‘Methods’’). With these approaches, a suite of discriminatory markers was identified for octopus cells (Figs. S6a, b, S7a, S8a, and Supplementary Data 1), a major projection neuron type for coincidence detection29,30. Antibodies to one of these markers, PHGDH, labeled neurons (i.e., cells with a large soma) restricted to the octopus cell area (OCA) (Fig. S6c). Indeed, when patch-clamp recordings from the OCA to label octopus cells with biocytin and post hoc immunostaining were performed for PHGDH, most labeled cells (12/13) were immunopositive for PHGDH (Fig. S6d). Similarly, Necab2 and Pbfibp1, among other genes, were confirmed as major markers for fusiform cells of the DCN (Figs. S6e, f, S7b, S8b and Supplementary Data 1). Immunostaining combined with patch-clamp analysis provided translation-level evidence for Necab2 as a highly selective marker for the entire population of fusiform cells (Fig. S6g, h).
Bushy cells consist of two major molecular subtypes
In anatomical and physiological studies, bushy cells have been divided into two populations, spherical and globular bushy cells (SBCs and GBCs, respectively), based largely on differences in their axonal projections31. Generally, SBCs are located anteriorly in VCN while GBCs are more posterior6. Some investigators have proposed two subtypes of SBCs (small and large), based on their axonal targets in lateral and medial superior olive6, although it is not clear how distinct these populations are. Our initial analysis identified two transcriptomically distinct bushy cell populations marked by high levels of Atoh7 or Hhip (Atoh7+ and Hhip+, Fig. 3a, b), which may correspond to SBCs and GBCs. We therefore performed FISH co-staining for Atoh7 and Hhip to determine the spatial ___location of these cell groups. Given that Sst is specifically expressed in the Atoh7 subtype rather than the Hhip subtype (Fig. 3b, c) and is expressed preferentially in SBCs rather than GBCs in VCN32, we also performed FISH co-labeling for Sst and Atoh7, and for Sst and Hhip. Both Atoh7 and Hhip signals were almost exclusively restricted to VCN (Fig. 3c), in line with our snRNA-seq data (Figs. 3b and S5a, b, d) and Allen ISH data (Fig. S8c). Furthermore, Atoh7+ cells were predominantly localized in the rostral anterior VCN (AVCN), while Hhip+ cells densely populated the posterior VCN (PVCN) and caudal AVCN, and less so rostrally (Fig. 3c), similar to SBCs and GBCs in terms of their typical locations31,33,34. In addition, Sst signals mostly overlapped with Atoh7 and much less so with Hhip, and double-labeled cells (Sst+/Atoh7+and Sst+/Hhip+) were restricted to rostral AVCN (Fig. 3c), further validating two molecularly distinct bushy cell subtypes marked by Atoh7 or Hhip. Based on the preferential ___location of each molecular subtype with differential Sst expression, the Atoh7 subtype may correspond to SBCs, while the Hhip subtype may correspond to GBCs.

a UMAP visualization of Hhip+ and Atoh7+ clusters and distribution of Patch-seq bushy cells in UMAP space, colored by cluster. Individual dots represent each Patch-seq cell. b UMAP visualization of two bushy cell clusters and normalized expression of Hhip, Atoh7, and Sst. Top: snRNA-seq, Bottom: Patch-seq. c FISH co-staining for Atoh7 and Hhip (left), Atoh7 and Sst (middle), or Hhip and Sst (right) in CN sagittal sections. Lines along the images indicate density of single-labeled or double-labeled cells along two axes of CN. Insets show the total counts of single-labeled and double-labeled cells across eight consecutive sagittal sections for each FISH experiment. d Left: A representative morphology of Hhip+ or Atoh7+ bushy cell. Axon (shown in red) is truncated. Right: Sholl analysis of Hhip+ or Atoh7+ bushy cell dendrites. Data are presented as mean ± SEM. n = 20 for Hhip+ cells from 15 mice, n = 51 for Atoh7+ cells from 20 mice, Two-way mixed model ANOVA. e Heat map displaying normalized expression of the top 20 DEGs for two subtypes. Scale bar: Expression level. Cell number in each subcluster is indicated below the figure. f Top: Example responses of Atoh7+ (left) or Hhip+ (right) bushy cell to a hyperpolarized current step and a near-threshold depolarizing step. Bottom-left: Individual APs from the Hhip+ and Atoh7+ cells shown above, aligned with onset of the depolarizing current. Bottom-right: Two most-discriminating features for Hhip+ and Atoh7+ cells, spike delay and spike duration (half-width). n = 67 for Hhip+ cells from 25 mice, n = 110 for Atoh7+ cells from 29 mice, Two-sided t-test. Data are presented as mean ± SEM. The dots above the bars represent the original values. g Volcano plot showing the log2 fold change (log2FC) and −log10(p-value) of detected genes comparing the two subtypes. The p-values were adjusted by Benjamini–Hochberg correction. Among DEGs are 15 genes encoding voltage-gated ion channels. See Fig. S9 for more analysis. Source data are provided in the Source Data file.
To determine if molecular subtypes of bushy cells are associated with unique physiological and morphological features, we examined the properties of each Patch-seq cell that was assigned to the Hhip or Atoh7 subtype. Two subtypes exhibited similar soma size and shape, but there were differences in their dendritic arborization (Supplementary Data 3). While the majority of Hhip bushy cells have only one primary dendrite (85%, 17 out of 20 cells), more than half of Atoh7 bushy cells have two or even three primary dendrites (~53%, 27 out of 51 cells), each with its own secondary and filamentous branches (Supplementary Data 3). In addition, Hhip bushy cells had short dendritic trunks capped with relatively compact dendritic tufts, while Atoh7 bushy cells had longer dendritic trunks that bifurcated and sent off diffuse, thin dendritic processes (Fig. 3d and Supplementary Data 3). Dendrites of either Atoh7 bushy cells or Hhip bushy cells were not oriented in any particular direction within the VCN.
Molecularly, two bushy cell subtypes were best discriminated by cell-adhesion molecule (CAM) genes (e.g., Plxdc2, Sema5a, Epha6, Kitl, Slit3, Cntnap5a, Sema3e), suggestive of their distinct connectivity and projection patterns (Fig. 3e). Bushy cells are biophysically specialized for encoding precise timing by expressing a prominent low-threshold K+ current and a hyperpolarization-activated current, which together bestow single-spike firing, and low input resistance and short time constants enabling brief and sharply timed synaptic responses35,36,37. Both bushy cell subtypes exhibited single-spike firing property, but they were electrophysiologically distinguishable with significant differences in numerous features, with spike delay and half-width being the most prominent (Figs. 3f and S9a, Supplementary Data 3). Hhip bushy cells had lower input resistances and shorter membrane time constant (thus shorter delays to firing onset), deeper and quicker voltage sags in response to hyperpolarizing currents, and fewer spikes in response to suprathreshold depolarizing currents than Atoh7 bushy cells (Fig. 3f, Supplementary Data 3), indicative of larger near-rest outward current with faster kinetics. These properties match a higher expression of ion channels active in that voltage range (Kcna1, Hcn1) in Hhip bushy cells (Figs. 3g and 7b)35,38. In addition, Hhip bushy cells had faster depolarization and repolarizations (larger dV/dt) resulting in narrower spikes (Fig. 3f, Supplementary Data 3), matching their enrichment of Kv3 (Kcnc3) (Fig. 3g), a potassium channel enabling brief APs for auditory neurons to follow high-frequency input with temporal precision39,40,41. In contrast, Atoh7+ cells had a higher expression of Kv2 (Kcnb2) in addition to a lower expression of Kv1 (Figs. 3g and 7b), which may enable them to fire more spikes than Hhip+ cells42.
Patch-seq profiles (with more detected genes than snRNA-seq) mapped to two main subtypes of bushy cells form distinct subclusters in snRNA-seq UMAP space (Fig. 3a), suggesting further bushy cell-type molecular heterogeneity. Subsequent subclustering analysis identified three subpopulations of Atoh7+ bushy cells (Atoh7/Dchs2, Atoh7/Tox, and Atoh7/Sorcs3) (Fig. S9b) and two subpopulations of Hhip+ bushy cells (Hhip/Calb1 and Hhip/Galnt18) (Fig. S9f). To further assess these subpopulations, we examined the properties of Patch-seq cells that were assigned to each subcluster, including their anatomical locations. Within Atoh7+ bushy cells, Atoh7/Dchs2 cells appear to be restricted to the nerve root area and PVCN, while Atoh7/Sorcs3 cells and Atoh7/Tox cells appear to have less spatial preferences (Fig. S9c). Electrophysiologically, Atoh7/Dchs2 cells displayed distinct firing properties compared to Atoh7/Sorcs3 and Atoh7/Tox cells, which shared similar firing patterns (Fig. S9d, Supplementary Data 3). Morphologically, Atoh7/Dchs2 cells generally had the longest dendritic trunks (stem) among the three subclusters (Fig. S9e, Supplementary Data 3). Within Hhip+ bushy cells, Hhip/Calb1 cells were preferentially localized in the posteroventral VCN, while Hhip/Galnt18 cells were preferentially localized in the dorsoanterior VCN (Fig. S9g). Hhip/Calb1 cells exhibited distinct firing properties from Hhip/Galnt18 (Fig. S9h, Supplementary Data 3). Additionally, the Hhip/Calb1 cells generally had shorter dendritic projections, but more complex dendritic tufts compared to Hhip/Galnt18 cells (Fig. S9i, Supplementary Data 3). These analyses support functionally and topographically distinct subpopulations within each bushy cell subtype.
T-stellate cells consist of two subtypes
Patch-seq mapping indicated that the Fam129a+ cluster corresponds to T-stellate cells (Fig. 2f, g). This cluster comprised two visibly separate lobes on the UMAP, suggesting two sub-populations of T-stellate cells (Figs. 1c and 4a). With more than 1100 DEGs identified between two putative sub-populations, we focused on Dchs2 and Fn1, two top DEGs expressed almost exclusively in one of the two sub-populations with high coverage (Figs. 4a, b and S10a, b). To validate their expression and determine how well these two genes separate T-stellate cell subpopulations, we performed FISH co-labeling for Fam129a and Dchs2, and for Fam129a and Fn1. Fam129a signals were restricted to VCN (Fig. 4c, d) and almost exclusively expressed in excitatory neurons (Fig. S7c), thus validating Fam129a as a general marker for T-stellate cells (Figs. 2f, S5a, b, S5d and S8d). Only a subset of Fam129a+ neurons co-expressed Fn1 (Fam129a+/Fn1+), and these double-labeled neurons were exclusively localized in the AVCN (Fig. 4c). Similarly, only a subset of Fam129a+ neurons co-expressed Dchs2, and these Fam129a+/Dchs2+ neurons were restricted to the VCN nerve root area and to PVCN (Fig. 4d). These data suggest two spatially separated subpopulations of T-stellate cells. To further substantiate this conclusion, we examined the ___location of Patch-seq cells assigned as T-stellate cells. These cells were mapped to either the Dchs2+ subcluster or the Fn1+ subcluster almost equally (Fig. 4a), allowing for an unbiased comparison of their spatial locations. Fn1+ T-stellate cells were restricted to the AVCN, while Dchs2+ T-stellate cells were predominantly localized in the PVCN and the nerve root area of the VCN. Importantly, these two subpopulations were spatially non-overlapping in the VCN (Fig. 4e), indicating that T-stellate cells could be classified into two subtypes by their anatomical locations. The existence of two T-stellate cell subtypes was further supported by their distinct morphological and electrophysiological properties. While both Fn1+ or Dchs2+ T-stellate cells exhibited typical morphology of T-stellate cells (Fig. 4f)43, there were subtle differences in their dendritic arborization (Supplementary Data 4). The dendrites of Fn1+ cells in general branched earlier and more profusely around the soma compared to that of Dchs2+ cells, while the dendrites of Dchs2+ cells arborized more extensively away from the soma with more branched, complex endings (Fig. 4f, g, Supplementary Data 4). In addition, the major dendrites and the terminal arbors of Fn1+ and Dchs2+ cells had distinct preferential angles with respect to their soma (Fig. 4h). Given that the major dendrites and the terminal arbors of T-stellate cells in general run parallel to tonotopically arranged ANFs (Fig. 4f) whose direction in each subregion is well-established44,45, such differences between two subtypes reflect their distinct anatomical locations in the VCN, further substantiating the conclusion that the subtypes of T-stellate cells are defined by their anatomical locations.

a Left: UMAP visualization of two subclusters of Fam129a+ neurons, colored by subcluster. Right: All Patch-seq cells mapped onto Fam129a+ cluster on the left. Individual dots represent each patch-seq cell. b UMAP visualization of normalized expression of Fam129a, Fn1, and Dchs2 in T-stellate cell cluster. c FISH co-staining for Fam129a and Fn1 in a CN sagittal section. Lines along images depict density of single-labeled or double-labeled neurons along the two axes of CN. Inset pie chart: proportion of double-labeled cells among all Fam129a+ cells counted across eight consecutive sagittal sections. Dashed contours indicate the CN region. Two images on the right are zoomed-in views of boxed regions on the left. Arrows point to double-labeled cells. d FISH co-staining for Fam129a and Dchs2 in a CN sagittal section. Inset pie chart shows the proportion of double-labeled cells among all Fam129a+ cells counted across six consecutive sections. Two images on the right are zoomed-in views. e Left: 2D spatial projection of Patch-seq T-stellate cells onto a sagittal view of CN, colored by subtypes. Right: Comparison of the distance to CN posterior edge between T-Fn1 and T-Dchs2. n = 12 for T-Fn1 cells from 10 mice, and n = 16 for T-Dchs2 cells from 10 mice; Data are presented as mean ± SEM; Two-sided t-test. f Left: Representative morphology of two Fn1+ and two Dchs2+ cells in a CN sagittal view. Right: zoomed-in of T-stellate cells shown on the left. Axons in red. g Sholl analysis of T-stellate cell dendrites. Data are presented as mean ± SEM. n = 6 for T-Fn1 cells from 6 mice, and n = 20 for T-Dchs2 cells from 12 mice; Two-way mixed model ANOVA. h Polar histograms showing the distribution of dendritic branch termination points with respect to soma of T-Fn1 and T-Dchs2. p = 3.0e-19 for the difference between the distributions using the Kolmogorov-Smirnov statistic. i Example responses of two T-stellate subtypes to current steps. j Comparison of electrophysiological features between two T-stellate subtypes. n = 16 for T-Fn1 cells from 13 mice, and n = 29 for T-Dchs2 cells from 16 animals, Mean ± SEM; Two-sided t-test. Source data are provided in the Source Data file.
T-stellate cells fire continuously in response to suprathreshold inputs, a biophysical property required for encoding sound intensity42,46. Both Fn1+ and Dchs2+ T-stellate cells fired tonically, as reported previously, but were physiologically distinguishable showing differences in numerous features (Figs. 4i, j, S10c and Supplementary Data 4). Among these, spike delay (latency to first spike at rheobase), time constant, and input resistance alone separated almost all Dchs2+ cells from Fn1+ cells (Fig. S10d). Such prominent physiological distinctions well aligned with their top DEGs being dominated by numerous genes encoding potassium channels (Fig. S10e). Notably, Fn1+ cells were enriched for expression of multiple leak potassium channels including K2P9.1 (Kcnk9), K2P10.1 (Kcnk10), and K2P12.1 (Kcnk12) and subthreshold-operating Kv channels including Kv7.4 (Kcnq4), Kv10.2 (Kcnh5) and Kv12.1 (Kcnh8), matching their lower input resistance and smaller time constant compared to Dchs2+ cells47,48 (Figs. 4g and S10d). In contrast, Dchs2+ cells were enriched for expression of Kv2 and Kv3 (Kcnb2, Kcnc2), two potassium channels required for auditory neurons to maintain high-frequency repetitive firing39,42,49, matching their more “fast-spiking’’ phenotype (e.g., no adaptation) compared to Fn1+ cells.
Molecular profiles of inhibitory neurons in CN
We also sought to determine the correspondence of glycinergic clusters to established inhibitory cell types in the CN. Among the six glycinergic/inhibitory clusters (including Golgi cells), four expressed both Slc6a5 and genes for GABA synthetic enzymes (Gad1/2, Fig. 1c), consistent with dual transmitter phenotypes described in CN25,46,50. The other two clusters, Slc6a5+/Sst+ and Penk+ cluster, expressed only Slc6a5 and no Gad1/2, indicative of being pure glycinergic populations (Fig. 1c). Notably, the Slc6a5+/Sst+ cluster likely corresponds to D-stellate cells in VCN, a known pure glycinergic population with specific Sst expression10,46. A definitive correspondence of glycinergic clusters to established inhibitory cell types (Figs. 1a and 5a) was then determined by targeting glycinergic cell types for Patch-seq (see “Methods’’). These included vertical, cartwheel, and superficial stellate cells (SSC) in DCN, and L- and D-stellate cells in VCN (Figs. 1a and 5a), each of which resides in distinct anatomical locations (except for L- and D-stellate cells) and exhibits unique morpho-electrophysiological features (Fig. 5a, b). As with excitatory neurons, we validated the expert classification and cell-type assignment of each Patch-seq cell. Both the random forest classifier and clustering-based k-means classifier demonstrated high accuracy in distinguishing each cell type pair using morphological features (see “Methods’’, Figs. 5c and S4c, d). The performance was also robust using electrophysiological features, especially when anatomical ___location information was incorporated (see “Methods’’, Figs. 5d and S4e–h). This robust performance across multiple modalities and methods supports cell-type assignment for each Patch-seq cell. UMAP projection of these Patch-seq cells resulted in five clusters which were then mapped onto the snRNA-seq dataset, successfully identifying transcriptomic clusters for established inhibitory cell types (Fig. 5e–g). As with excitatory neurons, we validated the molecular correspondences to inhibitory neurons using FISH. We validated the Stac+ cluster as cartwheel cells by FISH staining for Stac or Kctd12, two major putative marker genes for cartwheel cells (Figs. 5h, i,S5c–e, and S8e Supplementary Data 1). FISH staining for Penk in wild-type mice and Cre-positive cells in an existing Penk-Cre mouse line validated the Penk+ cluster as vertical cells (Figs. 5i, j and S11a), consistent with these inhibitory neurons being pure glycinergic51,52. FISH staining also validated Penk-Cre mice as a specific Cre driver for vertical cells, and thus electrophysiology and morphology of Cre-positive cells in DCN all matched with vertical cells (‘‘Methods’’; Fig. 5k, l). Interestingly, in addition to labeling vertical cells, Penk expression was also enriched in glycinergic neurons in the ‘small-cell cap’ (Figs. 5i–k and S8f), a narrow CN region between the granule-cell and magnocellular domains of the VCN53. This topographically unique inhibitory subgroup was devoid of Gad1 (i.e., purely glycinergic, Fig. S11b), and thus genetically more similar to vertical cells than to other small glycinergic neurons in VCN (i.e., Gad1+ L-stellate cells, Figs. 1c and S11c, also see below). The Penk+ cluster thus includes both vertical cells and glycinergic cells in the small-cell cap.

a Examples of reconstructed inhibitory neurons in a sagittal section, colored by cell type. b Example responses of inhibitory cells to current steps. c Left: UMAP visualization of morphological features of 116 inhibitory neurons (M-cluster), colored by expert cell type. Vertical: n = 38; SSC: n = 13; Cartwheel: n = 15; D-stellate: n = 15, L-stellate: n = 15. Right: Performance (Ave: 96.9%) of the classifier trained with morphological features. d Left: UMAP visualization of electrophysiological features along with anatomic locations (DCN:1; VCN:0) of 172 inhibitory neurons (E-cluster). Vertical: n = 50, SSC: n = 21; cartwheel: n = 21; D-stellate: n = 26, L-stellate: n = 53. Right: Performance (Ave: 97.9%) of the classifier trained with electrophysiological features and anatomic locations. See Fig. S4e, f for more analysis. e Left: UMAP visualization of 94 Patch-seq inhibitory neurons, colored by T-clusters. Right: Proportion of Patch-seq cells in each T-cluster assigned to a specific cell type. T-cluster 1: 12/12 are cartwheel cells; T-cluster 2: 20/21 are D-stellate cells; T-cluster 3: 35/35 are L-stellate cells; T-cluster 4: 10/12 are SSCs; T-cluster 5: 12/14 are vertical cells. f Top: 94 Patch-seq inhibitory neurons mapped onto snRNA-seq UMAP space, colored by T-cluster. Bottom: Proportion of Patch-seq cells assigned to each T-cluster in (e) mapped onto one of 13 distinct clusters in UMAP space. g UMAP visualization of all CN neuronal clusters, along with annotation with morpho-electrophysiological types. h UMAP visualization and annotation of all glycinergic CN cell types, and normalized expression of their respective marker genes. i FISH co-staining for Slc6a5/Stac, or for Slc6a5/Penk in sagittal sections. Lines indicate density of double-labeled neurons along two CN axes. Insets show proportion of double-labeled cells among all Stac+ or Penk+ cells counted across eight or six sections. j FISH co-staining for Slc6a5/tdTomato (left), or for Slc17a6/tdTomato (right) in sagittal sections in Penk-Cre:Ai9 mice. Insets show proportion of double-labeled cells among all tdTomato+ cells counted across six sections. k Micrographs showing labeled cells for recording (left), example responses to current steps (middle), and morphologies of labeled neurons (right) in Penk-Cre:Ai9 mice. l Distribution of labeled cells (red) in electrophysiological or morphological UMAP space of all inhibitory neurons. Source data are provided in the Source Data file.
FISH staining further validated the Sst+/Slc6a5+ cluster as D-stellate cells that could be discriminated with a combination of two genes or even single genes10 (Figs. S5e, S11d, and Supplementary Data 1). To validate Kit+/Cdh22+cluster as L-stellate cells, we performed co-labeling for Kit and Cdh22, and for Kit and Esrrb, two combinations predicted to distinguish L-stellate cells (Figs. 5f–h and S5e). In line with snRNA-seq data (Figs. 1c, 5h, and Fig. S5e), Kit+ cells were almost exclusively glycinergic with a smaller soma size compared to Kit− glycinergic neurons (Fig. S11e), and Kit+ neurons labeled by Cdh22 or Essrb transcripts (Kit+/Cdh22+ or Kit+/Essrb+) were mainly detected in the VCN (Fig. S11f), consistent with the ___location of L-stellate cells. Interestingly, double-labeled glycinergic neurons were sparsely detected in DCN as well (Fig. S11f), suggesting that the Kit+/Cdh22+ cluster may also include glycinergic neurons in the main body of DCN that are neither cartwheel cells nor vertical cells (Fig. S11g, h), a hitherto undescribed cell type. Finally, to validate Kit+/Ptprk+ clusters as SSCs, we performed co-staining for Kit and Ptprk, and for Kit and Hunk, two potential combinations to discriminate SSCs (Figs. 1c, 5f–h and S5e). In line with our analysis, double-labeled glycinergic neurons (Kit+/Ptprk+ or Kit+/Hunk+) were mostly detected in the molecular layer of the DCN (Fig. S11i). Double-labeled cells were also sparsely detected at the VCN, likely reflecting the expression of Hunk or Ptprk in a small subset of L-stellate cells (Figs. 5h and S11i).
Molecular basis of CN cell type identity and wiring specificity
Our analysis indicates that all cell types in CN with anatomical and physiological identity (each with unique morphophysiological features, connectivity, projection patterns, and function)4,5,6 can be defined by their transcription profiles, suggesting that cellular specializations are rooted in differential gene expressions (Fig. 5g). Gene profiles conferring neuronal phenotypes are often dictated by differential expression of a small set of gene families54,55,56, and determined by the concerted action of transcription factors (TFs)57,58,59,60. We thus performed supervised computational screen using MetaNeighbor algorithm54 to identify gene families whose differential expression could reliably predict CN neuron types (‘‘Methods’’). The top-performing gene families were dominated by three functional categories including TF (‘‘Methods’’, Fig. 6a, b, Supplementary Data 5), in line with the notion that TFs are key regulators of cell type identity57,58,59,60. We therefore set out to characterize cell types by their TF activity and ask if CN cell types are defined by specific yet overlapping TF network features. Using a more comprehensive TF database with precisely annotated TF families61, we found differentially expressed TFs among CN cell types were predominantly from the C2H2 zinc finger family (‘‘Methods’’, Supplementary Data 5); approximately one-third of the most differentially expressed TFs belong to C2H2-Zinc finger TFs, a pattern of TF expression that appears unique to this brain region54,62,63. Notably, no single TF exhibited expression strictly correlated with a specific cell type, highlighting the importance of combinatorial TF codes in specifying cell type identity (Fig. 6c, d; Supplementary Data 6). We identified a set of TFs that are selectively co-expressed in specific cell types and hence may define identities of cell types (Fig. 6c, d, Supplementary Data 6). Cell type identity is often determined by a small set of core TFs that exhibit synergistic transcriptional regulation58,64,65,66,67,68,69,70,71. To investigate the core TFs for each CN cell type, we leveraged a computational method that capitalizes on key principles of core TF expression: relatively high and cell-type-specific expression and transcriptional synergy64,72. This analysis predicted a distinct combination of core TFs for each cell type (synergistic identity TF cores, Supplementary Data 7). Supporting predictions, many core TFs identified for projection neurons (Pax6, Ebf2, Rorb, Tshz3, Bnc2, Tead1, Meis1, Bcl11a, Prox1, Zfp536, Zfhx3, Ikzf2, Zfp322a) are known for their role in maintaining cell identity, and deletion of these TFs in postnatal age or adult results in the loss of markers and physiological properties characteristic to their identity73,74,75,76,77,78,79,80. In addition, these predicted core TFs, along with many other cell type-enriched TFs (Fig. 6c, highlighted in red) are also expressed during early CN development, including in progenitor cells (Supplementary Data 7, 8)81,82,83,84,85, suggesting developmental continuity of TF expression from embryonic precursors to mature neurons. For example, while TFs like Atoh7, Ebf2, Pax6, Lhx9, Zfp322a, Sox11, Tshz2, Klf3, and Camta1 are each enriched in a specific mature projection neuron type (Fig. 6c), they exhibit early and broad expression within the developing Atoh1 lineage that gives rise to these neurons (Supplementary Data 8)81,83,84. Similarly, many predicted core TFs (e.g., Tfap2b, Myt1l, Npas3, Pax2, Sox5, Etv6, Id2, Nfia, Nfix, Nfib, Maf, Pparg, Rora, Arid5b, Mef2c) and other TFs enriched in each CN inhibitory cell type (Fig. 6d, highlighted in red) are also expressed during early neural development and are known to be essential for the differentiation and specification of GABAergic/glycinergic neurons (Supplementary Data 7, 8). These observations suggest that many developmental transcription programs, initiated early in post-mitotic differentiation, persist in mature neurons that may act in concert to specify and maintain cell phenotypes and identity54.

a Distribution of AUROC values of 5,735 GO terms across all 13 cell types in the snRNA-seq dataset. Red, AUROC ≥ 0.8. Right: GO term probability density by keywords: “synaptic”, “cell adhesion” and “ion channel” are skewed with AUROC ≥ 0.8. b AUROC value distribution of 1424 HGNC gene families across all 13 cell types. 104 families with AUROC ≥ 0.8 (red bars) are classified into seven categories (pie chart). See Supplementary Data 5 for more details. c Heatmap displays the z-scored expression levels of the top 10 specific TFs for each excitatory projection neuron type. TFs highlighted in red show expression in the developing cochlear nucleus, including progenitors and distinct developmental stages of projection neurons (see Supplementary Data 8 for reference support). d Heatmap showing the z-scored expression levels of the top 10 specific TFs for each inhibitory cell type. TFs in red have been shown to be involved in the differentiation and specification of glycinergic/GABAergic interneurons in cochlear nucleus or other brain regions (see Supplementary Data 8 for reference support). e Top-performing cell-adhesion molecule (CAM) families, their CAMAUROC value, and their roles in synaptic connectivity. “+” denotes the degree of involvement in the listed function. f Heatmap showing the differential expression of 9 CAM and 2 carbohydrate-modifying enzyme families across excitatory projection neuron types. g Heatmap showing the differential expression of 9 CAM and 2 carbohydrate-modifying enzyme families across inhibitory cell types.
CN neurons exhibit precise wiring specificity, each receiving inputs from and extending output to specific pre- and post-synaptic neurons, respectively4,5,6, while CAMs are key determinants of neuronal identity and wiring specificity54,86. In line with these, gene ontology (GO) terms containing the keyword “synaptic” and CAM gene families gave the highest score in distinguishing CN cell types (Fig. 6a, b, Supplementary Data 5). We selected 342 genes encoding all major neuronal CAMs implicated in neural development and organized them into 10 CAM groups according to sequence homology and receptor-ligand relationships54,87,88, and nearly all groups are among the most distinguishing families (Fig. 6d, Supplementary Data 5). Among these CAM genes, 135 show highly distinct cell type profiles (Supplementary Data 6). Notably, multiple CAM families each manifest differential expression among CN cell types (Fig. 6f, g). For example, Slit2/3 and their receptors (Robo1/2); netrin family (Ntn4, Ntng1/2) and their receptors (Dcc, Neo1); contactins and their binding partners; semaphorins and their receptors (plexins), exhibited highly differential expression, often with binary on/off patterns among five major projection neuron types (Fig. 6f). These CAM families also exhibited highly differential expression among inhibitory neuronal cell types, often with distinct family members being employed from those in projection neurons (Fig. 6g). These receptor-ligand pairs mediate axonal guidance and cell-cell recognition (either attractive or competitive/repulsive) during neuronal development, and thus might be key molecules for the establishment and maintenance of specific projection pattern of each CN cell type54,89. Each of the major synaptic CAM families including neurexin, neuroligin, protein tyrosine phosphatases (PTPR), leucine-rich repeat proteins (LRR), cadherin, neural IgCAM was also differentially expressed among CN cell types (Fig. 6f, g). Cell-specific expression of these families, particularly LRRs, might contribute to post- and trans-synaptic specializations that customize the property of synapse types defined by each neuron and their specific target neuron54,89,90,91. In addition, two families of carbohydrate modifying enzymes (sulfotransferases and sialyltransferases) also exhibited highly distinct cell type profiles (Fig. 6f, g, Supplementary Data 5), which might increase the molecular diversity of glycosylated CAMs and proteoglycans on the cell membrane and in extracellular matrix92,93,94. Together, cell- and synaptic CAM families likely constitute a multi-faceted cell-surface molecule code throughout the neuronal membrane of each CN neuron to determine their specific wiring.
Molecular underpinnings of functional and biophysical specializations of CN neurons
We observed that the families of genes encoding ion channels are also highly predictive of CN cell types (Figs. 6a, b, 7a, S13a and Supplementary Data 5). Two top families in this category encode potassium channels, while the gene families encoding sodium channels were not predictive of cell type identity (Figs. 6a, b, 7a and Supplementary Data 5). This suggests that potassium channels, rather than sodium channels, are the major determinants for biophysical features and firing properties of specific CN cell types (Figs. 2 and 5). To further explore this contention, and identify potential molecular determinants, we took advantage of the bimodal gene expression and physiological data in the Patch-seq dataset and performed sparse reduced-rank regression to identify gene transcripts that were predictive of firing properties of CN neurons95,96. To enhance interpretability, our analysis was restricted to voltage-sensitive ion channels within excitatory projection neurons (“Methods”, interneurons were excluded from the analysis due to insufficient data). These neurons exhibit either phasic firing (single-spike: bushy cells and octopus cells) or tonic firing (T-stellate and fusiform cells), which are differentiated by a set of highly correlated physiological features (Figs. 2b, 7b, S12d–f and Supplementary Data 2). Our sparse reduced-rank regression model selected 25 ion channel transcripts that achieved near-maximal predictive accuracy (Figs. 7b and S12b, c). With additional statistical evaluations and the literature review, we identified two sets of transcripts among these 25 that are strongly predictive of the physiological hallmarks of projection neurons (“Methods”, Figs. 7b and S12c–f). One set of transcripts, including Kcna1 (Kv1.1), Hcn1 (HCN1), and Kcnq4 (Kv7.4), were predictive of single-spike firing and associated features including low input resistance, short time constant, a high rheobase and threshold for spike generation, large voltage sag, and small AP amplitude (Figs. 7b and S12c, d). This analysis suggested Kv1.1 and HCN1 as candidate molecular determinants of single-spike firing property, in line with prior evidence35,36,37,97,98,99, but also suggested another subthreshold-operating K+ conductance conferring single-spike firing property of CN neurons, Kv7.4-mediated conductance100,101,102,103. In contrast, a distinct set of transcripts including Kcnb2 (Kv2.2) and Kcnc2 (Kv3.2) were predictive of high-frequency tonic firing and associated features including large AHP, large AP amplitude, quick AP repolarization, and narrow APs (Figs. 7b and S12c, e), matching the role of these channels enabling repetitive high-frequency firing (by expediting AP repolarization and speeding the recovery of Na+ channels from inactivation)39,42,49,104,105,106. Among other relevant transcripts selected, two encoding potassium channels for transient A-type current, Kcna4 (Kv1.4) and Kcnd3 (Kv4.3), correlated with AP delay (Figs. 7b and S12c, f), matching the transient nature of A-type current that mediates the delay to AP onset107,108,109. Thus, Kv1.4 and Kv4.3 could mediate prominent A-currents in fusiform cells, determining their three distinct in vivo response patterns (“pauser”, “buildup”, and “chopper”)108,110.

a Dot plots showing scaled expression of the genes encoding ion channels across CN cell type (from snRNA-seq dataset). Sodium and potassium channel subtypes or subunits with low expression levels (<20% fraction of the cell in any cell type) were excluded in the figure. Circle size depicts the percentage of cells in the cluster in which the marker was detected (≥1 UMI), and color depicts the normalized average transcript count in expressing cells. b Sparse reduced-rank regression (RRR) model to predict electrophysiological features by the expression patterns of 119 ion channel genes (middle, cross-validated R2 = 0.35). The models selected 25 genes. Cross-validated correlations between the first three pairs of projections were 0.83, 0.65, and 0.61. Both transcriptome (left) and electrophysiology (Right) were embedded in the latent space (bibiplots). In each biplot, lines represent correlations between a feature (gene expression or electrophysiology) and two latent components; the circle corresponds to the maximum attainable correlation (r = 1). Only features with a correlation above 0.4 are shown. Source data are provided in the Source Data file. c Dot plots showing scaled expression of genes encoding ionotropic glutamate receptors in each CN cell type. AMPARs AMPA receptors, NMDARs NMDA receptors, KARs Kainate receptors, GluDRs delta glutamate receptors. See Fig. S12 for additional information.
In addition to biophysical specializations, CN neurons rely on specialized structural and synaptic features to fulfill their tasks111,112. Bushy cells, for example, require large presynaptic terminals (endbulbs of Held) with postsynaptic fast-gating AMPA receptors to enable precise spike timing with AN inputs112. These AMPA receptors are formed by GluR3 (Gria3) and GluR4 (Gria4), and are devoid of GluR2 (Gria2), thus conferring calcium permeability, large conductance, and exceptionally rapid gating kinetics113,114,115. We therefore examined expression patterns of ionotropic receptors across cell types (Fig. 7c), and found that such exceptional AMPARs were also expressed in other CN cell types targeted by ANFs (i.e., T-stellate cells, octopus cells, vertical cells), but not in cell types targeted by non-ANF fibers, matching the observation that the very fastest synaptic currents are found only in neurons of the auditory system111. Thus, cartwheel cells, targeted by parallel fibers only, mainly expressed GluR1 (Gria1) and GluR2 (Gria2)116, an AMPAR composition with slow gating kinetics and calcium impermeability, while AMPARs in UBC, targeted by the mossy fibers, formed almost exclusively by GluR2 (Gria2, Fig. 7c), an uncommon form of native AMPAR117. Furthermore, fusiform cells, targeted by both ANF and parallel fibers, have mixed slow (GluR2) and fast AMPAR (GluR3/4) composition (Fig. 7c)118,119. This observation thus supports that postsynaptic AMPAR is specialized according to the source of synaptic input, likely illustrating a general rule governing synaptic specialization across the brain116,118,119. As for the other two types of ionotropic glutamate receptors (iGluRs; kainate and NMDA receptors), the expression of these relatively slow kinetic receptors was in general low in time-coding neurons (Fig. 7c), a necessary feature for fast high-fidelity transmission120,121.
CN neurons are subject to cell type-specific synaptic inhibition and neuromodulation to achieve or refine their functional specialization122,123,124. Our analysis indicates a wide variety of receptors for glycine, GABA, and neuromodulators in CN are cell-type specific (Fig. S13b, c, Supplementary Data 5), in line with prior cellular and systems-level physiological observations. Particularly, a set of protein families in second messenger pathways (downstream of cell-surface receptors) including protein kinases, regulators of G-protein signaling, small guanosine triphosphatases (GTPase), and GTPase regulatory proteins (e.g., Rho-GEFs) are among the most differentially expressed gene families (Fig. 6b, Supplementary Data 5). Differential expression of signaling proteins likely shapes the specificity and spatiotemporal dynamics of broadly acting second messengers, which might provide the mechanism and capacity to maintain the diversity of CN neuron morphology, synaptic connection, and neurite motility125. Finally, approximately half of the known deafness genes showed expression in single types or small sets of cell types, suggestive of their essential roles for central auditory processing (Fig. S13d). Our dataset thus could provide a starting point to illustrate central components of hereditary deafness, and to differentiate poorly recognized central causes of deafness from peripheral causes (Fig. S13d)126,127.
Discussion
One of the biggest challenges in defining a cell-type taxonomy of the brain is to meaningfully reconcile definitions of cell types across the many modalities of measurement typically used to characterize brain cells128. Here in the CN, the first relay station in the auditory system where all high-order parallel processing pathways are initiated, we succeeded by first defining neuronal populations using unbiased single-nucleus RNA sequencing, and then relating these populations to anatomical, physiological, and morphological cell identities using Patch-seq combined with histology and FISH. We reveal a clear correspondence between molecularly defined cell populations and all previously defined cell types with known behavioral significance, as well as discover multiple hitherto unknown subtypes of the major projection neurons in the CN. We thus built a comprehensive and thoroughly validated atlas of cell types in the CN that harmonizes the cell type definitions across all modalities.
Neurons in the CN are highly specialized for processing different aspects of acoustic information and initiate distinct parallel pathways that encode sound ___location, intensity, frequency content, and spectrotemporal modulations2,3. Our analysis indicates that cellular and functional specializations for parallel auditory processing are bestowed by differential transcriptional programs dominated by a small set of gene families, and that these programs can be leveraged to define, discriminate, target, and manipulate the distinct processing pathways. Given the organizational similarities observed across the stations and different sensory systems129, we anticipate that this molecular design applies to downstream auditory stations and may extend beyond the auditory modality. Molecular definitions of cell types that initiate parallel pathways in sensory systems in general, and the auditory system in particular, open up new avenues for functional dissections of parallel processing. To date, no molecular tools are available to manipulate cell types in the CN to determine the impact on downstream sound processing and behavior. The molecular entry points and molecular basis of each cell type provided here ensure the rational design of a molecular toolbox (including Cre lines and in vivo cell type-specific phototagging130) to link the activity of brainstem microcircuits to cortex population response patterns, and to perceptual learning or performance. We have validated an existing Penk-Cre line as a specific driver line for vertical cells131. The high cell-type expression specificity of Atoh7, Hhip, Necab2, Stac, and Phgdh makes them excellent candidates for the development of Cre lines to target and manipulate SBC, GBC, fusiform cells, cartwheel cells, and octopus cells, respectively. Leveraging the extensive list of marker genes, transcription factors, CAMs, and ion channels with relatively specific expression patterns across cell types (Supplementary Data 1, Figs. 6 and 7), intersectional approaches utilizing two or more of these genes could offer an alternative strategy for targeting and manipulating specific cell types. In addition, the cellular and transcriptomic atlas of the cochlear nucleus (CN) we generated offers an extensive resource and reference to investigate and identify hitherto unknown central components of hearing disorders132 and to illustrate central consequences of hearing loss and auditory trauma at a molecular level, potentially revealing the molecular underpinnings of neurological conditions such as tinnitus133,134. The atlas may also facilitate the rational design of auditory brainstem implants135.
Identification of additional cell types in mouse CN
Our combined snRNA-seq/Patch-seq approach molecularly defined all previously described excitatory cell types, except for the sparse giant cells, which likely have a similar molecular identity to fusiform cells136,137. This approach also revealed molecular subtypes within the classically defined cell types. By using transcriptomic clustering to parse electrophysiological and morphological datasets into different groups, we were able to identify subtypes of bushy cells and T-stellate cells with distinct properties. Two molecular subtypes of bushy cells have distinct anatomical preferences and response kinetics, reminiscent of GBC and SBC, a conventional category of bushy cells with distinct projection targets and functions138,139,140. We also found that within the groupings of Atoh+ and Hhip+ bushy cells, distinct variation exists in transcriptomic, topographic, and electrophysiological features, supporting the concept that SBCs and GBCs are broad cell types that each may contain distinct subtypes. More generally, using transcriptomic clusters to parse cells of similar electrophysiological features proved to be a powerful approach for extracting important biological meaning from the variance in a set of physiological measurements.
T-stellate cells play remarkably diverse roles in auditory function, having axonal projections within the CN and but also extending to multiple downstream nuclei as far as the auditory midbrain6. Such broad projections suggest diverse physiological functions, yet there has been little prior information to support subtypes of the T-stellate cell class. Our finding that T-stellate cells comprise two groups with prominently distinct gene expression, intrinsic physiological properties, and topography within the CN provides clear evidence for subtypes of T-stellate cells. In vivo recordings refer to T-stellate cells as “choppers”, based on a frequency-independent, regular pattern of spikes observed in spike histograms. Interestingly, several classes of choppers have been distinguished in a wide variety of species including mice, with sustained and transient response patterns being the most common141. It is possible that the two T-stellate subtypes in the present study correspond to these different classes of chopper. Beyond morphological and electrophysiological properties, studies of these subtypes of bushy and T-stellate cells in terms of their synaptic inputs and axonal projections may uncover additional parallel auditory pathways and principles of auditory coding in the mammalian brain.
Until recently, the greatest diversity of inhibitory interneurons of CN was thought to be in the DCN142. The recent discovery of L-stellate cells in VCN indicated that VCN may also harbor multiple interneuron types besides D-stellate cells10. Here, we show that L-stellate cells, defined simply as small glycinergic neurons in the VCN, are molecularly diverse. Those glycinergic neurons in the small cell cap resemble vertical cells in DCN (Penk+, purely glycinergic), differ from other L-stellate cells (dual transmitters, Penk−), and thus may comprise a hitherto unknown type of inhibitory neurons in the VCN. Within the DCN Penk and Stac were selective markers for vertical and cartwheel cells, respectively. As these two cell types play distinct roles in gating auditory and multisensory signals in DCN, the identification and development of specific genetic tools based on their specific markers should facilitate future studies of their role in auditory processing. Another important interneuron marker is Kit, a gene specifically labeling those small glycinergic neurons across CN (Fig. 5 and Fig. S11) (i.e., excluding vertical cells, cartwheel cells, and D-stellate cells), and its mutations cause deafness suggestive of an important functional role in CN interneurons (Fig. S13d)143,144. Guided by Kit expression, we observed small glycinergic neurons in the DCN that are neither vertical cells nor cartwheel cells, suggesting a hitherto unrecognized inhibitory cell type in the DCN. These glycinergic neurons in the DCN appear to be genetically similar to L-stellate cells in the VCN with dual transmitter phenotypes (Fig. 1c, Fig. 5, and Fig. S11).
Molecular underpinnings of cellular and functional specializations for auditory processing
Cell types with unique phenotypes are deeply rooted in the differential expression of many proteins. What types of genes distinguish cell types and also readily inform their phenotypes? TFs are widely thought to establish and maintain cell-type identity, while CAMs determine wiring specificity. Consistent with this contention, we found that TF and CAM gene families are among the most differentially expressed across CN cell types. The maintenance of cell identity involves the coordinated action of many TFs (“combinatorial codes”). In addition, the expression of only a handful of key TFs (core TFs) is often sufficient to maintain the identity of a cell subpopulation64,65,66,67,68,69,70,71,145; depletion of these regulators causes significant alteration of cell identity, while forced expression of these regulators can effectively reprogram cells to a different cell type146. These TFs have been called terminal selectors, which activate or repress the cohort of effector genes that determine morphological, physiological, and molecular features specific to each cell type57,59. In the CN, while developmental programs governing early CN cellular differentiation are well defined132,147, the key TFs governing post-mitotic specification and maintenance of CN cell types remain largely unknown. Our analysis with bioinformatic tools predicts the potential candidate TFs for each CN cell type, offering targets for experimental validation. Furthermore, our dataset could serve as a valuable source to employ various approaches for identifying and exploring additional candidate TFs essential for CN cell type identity. Notably, most of these TFs have been shown to be expressed in different developmental stages of CN neurons81,85,148,149, suggesting that neuronal identities in adulthood are maintained by sustained expression of the same set of TFs54,60. Further investigation across different developmental stages may help reveal definitive TFs and regulatory programs that define each cell type. Ultimately, identifying these TFs will be critical for designing cell engineering strategies targeting distinct CN cell populations.
Synapses and neural circuits form with great specificity during brain development ensuring the accurate flow of neural information. CN represents a “prototype” sample; each CN neuron is organized tonotopically and receives the specific inputs from spinal ganglion neurons and projects downstream targets with high anatomical and target specificity6. Although this property of synapses and neural circuits in CN has been extensively investigated, molecular mechanisms underlying this property are largely unknown. Many studies suggest that the construction of the circuits, including central auditory circuits is at least partly defined by a combinatorial code of levels of cell-cell adhesion and axon guidance molecules54,89,133,134,150. The different types of CN neurons might assemble various types of CAMs and their receptors in time and space to mediate the selective contributions of diverse attractants and repellents from cells of their surrounding environment134,151,152. Our genome-wide mRNA sequencing data allows a comprehensive depiction of the molecular CAM framework dictating the anatomical/cell/synaptic-specific organization of CN circuits. Our analysis suggests such a molecular framework is encoded by a small set of CAM gene families, and combinatorial and coordinated expression of select members across these families together with their binding specificities contributes to precise neuronal connections and projection patterns. Many of these CAMs have been previously shown to be essential for the precise circuit wiring in the auditory system151,152,153,154,155,156 and in various other brain regions. A few seem specific to the auditory system, including Cntn5157,158, Epha10159, and Carmil1160. Importantly, these CAMs are associated with various neurodevelopmental disorders including hearing loss (e.g., Kirrel3, Ptprd, Dcc, Slit2/3, Robo1, Ntng1/2, Cntn5, Cntn6, Cdh13, Dmd, Utrin, Epha10, Carmil1)159,160,161,162,163,164,165,166,167,168,169, dyslexia (e.g., Cntnap5a, Robo1)170,171, and speech sound language disorders (Robo1)165, autism spectrum disorder (ASD) and intellectual disability (ID) with hearing defect as an essential component, constituting candidate molecules for experimental follow-up.
Our transcriptomic dataset offers a comprehensive resource to unravel molecular underpinnings of morphological, synaptic, and biophysical specializations required for CN neurons to encode auditory signals. Our analysis suggests that the biophysical specializations and diversity of CN neurons is bestowed by a small set of ion channel families, particularly potassium channels. Notably, we predicted three transcripts including Kcna1, Hcn1, and Kcnq4 as potential molecular determinants for single-spike firing property, a property required for time-coding neurons to encode precise AP timing for binaural sound detection172, in line with the observations that mutations of these genes impair binaural hearing173,174,175,176 (Fig. S13d). While Kv7.4 currents in time-coding neurons were previously undescribed despite strong Kcnq4 expression in CN, their role in enabling single-spike firing in concert with Kv1 is well-established in many other cell types100,101,102,103, and KCNQ4 mutations cause profound hearing loss in human DFNA2 that could not be solely explained by hair cell dysfunctions177,178. Another important ion channel for CN projection neurons is high-threshold Kv3, and these neurons appear to employ distinct subunits of this family to fulfill their unique biophysical needs. While Kv3.2 (Kcnc2) seems crucial for the tonic firing of T-stellate and fusiform cells (Fig. 7b), Kv3.3 transcript (Kcnc3) is specifically enriched in time-coding neurons (Fig. 7a) to possibly mediate their fast spike repolarization, a necessary feature for the use of both temporal and rate coding in acoustic processing39,40. This observation may explain why SCA13 patients (with KCNC3 mutations) exhibit impaired interaural timing and intensity discrimination179 (Fig. S13d). Interestingly, our analysis did not support a significant contribution of sodium channel diversity to remarkable biophysical properties of CN projection neurons (Supplementary Data 5)180. These neurons exhibited similar expression patterns of sodium channel α subunits, primarily Scn1a, Scn2a, Scn8a, and unexpectedly, Scn9a (Fig. 7a), a subunit previously thought to be restricted to the peripheral nervous system181. However, these genes were more differentially expressed among CN inhibitory neuron cell types (Fig. 7a), suggesting a greater role for sodium channels in shaping the diverse firing properties of CN interneurons. Further investigation is warranted to fully elucidate the contribution of ion channel diversity, including both potassium and sodium channels, to the biophysical properties of CN interneurons.
Methods
Animals
All experiments were performed according to the guidelines of the Institutional Animal Care and Use Committee (IACUC) of Baylor College of Medicine and Oregon Health and Science University. Wild-type mice or transgenic mice of both sexes on the C57/BL6/J background at the age of postnatal (P) 22-28 were used for single-nucleus RNA sequencing and slice electrophysiology (including Patch-seq). To help distribute our sampling across cell types and to increase the chance of recording certain specific cell types, GlyT2-EGFP (RRID:MGI:3835459) mouse line was used to target glycinergic/inhibitory neuronal populations (labeled neurons), while unlabeled cells in this line were targeted for excitatory neuron recording10. SST-Cre (RRID:IMSR_JAX:013044) cross with Ai9 (RRID:IMSR_JAX:007909) mouse line was crossed with GlyT2-EGFP mice to identify D-stellate cells and L-stellate cells in the CN for patch clamp recordings10. Some recordings from excitatory neurons were performed using wild-type mice as well. In this study, we used 222 mice (Mus musculus) in total (121 males and 101 females), including 132 wild-type mice, 61 GlyT2-EGFP mice, 17 Penk-Cre:Ai9 mice, and 3 Gabra6-Cre (RRID:IMSR_JAX:007909):Ai9 mice. Penk-Cre (RRID:IMSR_JAX:025112) mice are a gift from Yong Xu Lab at Baylor College of Medicine131, while Gabra6-Cre mice are a gift from Susumu Tomita Lab at Yale University. All animals were maintained in the animal facility with a light cycle from 6 am to 6 pm daily, with temperatures ranging from 68 to 72 °F and humidity ranging from 30% to 70%.
Single-nucleus extraction and single-nucleus RNA sequencing (snRNA-seq)
Mice were deeply anesthetized with 3% isoflurane and decapitated immediately, and their brains were immediately removed from the skull and then transferred into an iced oxygenated NMDG solution (93 mM NMDG, 93 mM HCl, 2.5 mM KCl, 1.2 mM NaH2PO4, 30 mM NaHCO3, 20 mM HEPES, 25 mM Glucose, 5 mM Sodium Ascorbate, 2 mM Thiourea, 3 mM Sodium Pyruvate, 10 mM MgSO4 and 0.5 mM CaCl2, pH 7.4) for further dissection. Cochlear nuclei (CNs) were dissected from each brain under a stereoscopic microscope and then transferred into a 1.5 mL Eppendorf tube, and tubes were immediately immersed in liquid nitrogen. Brain tissues were then transferred into a −80 °C freezer for long-term storage. Each tissue sample was pooled from 8 to 10 mice for single-nucleus extraction and single-nucleus RNA sequencing (snRNA-seq).
Single nucleus extraction from brain tissues followed a published protocol182. Briefly, frozen tissues were homogenized with 2 mL homogenization buffer (HB, 250 mM Sucrose, 25 mM KCl, 5 mM MgCl2, 10 mM Tris, 1 mM DTT, 1x Protease Inhibitor, 0.4 units/µL Rnase Inhibitor, 0.1% Triton X-100) in Wheaton Dounce Tissue Grinder. Cell debris and large clumps were removed with a 40 µm cell strainer (Flowmi). The homogenate was centrifuged at 1000 g for 5 min at 4 °C. The pellet was resuspended with 25% iodixanol in the centrifugation buffer (CB: 250 mM sucrose, 25 mM KCl, 5 mM MgCl2, 10 mM Tris) and layered on top of 29% iodixanol. The mixture was centrifuged at 13,500 g for 15–30 min at 4 °C. The supernatant was carefully removed without disrupting the nuclei pellet. The nuclei pellet was resuspended with the resuspension buffer (RB: 1 % UltraPure BSA in Rnase-free PBS).
Nuclei were counted using a hemocytometer or CellCounter (Countess II, Invitrogen) and diluted to ~1000 nuclei/µL for single-nucleus capture on the 10x Genomics Chromium Next GEM Single Cell 3’v3 system following the standard user guide. Droplet-based snRNA-seq libraries were prepared using the 10x Genomics Chromium Single Cell kit according to the manufacturer’s protocol and were sequenced on the Illumina NovaSeq 6000 or Hiseq 4000 platform with a pair-end 150 bp strategy (average depth 40k–50k reads/nucleus). Cell Ranger (version 5.0.1) with the Mus musculus genome (GRCm38) and annotation GTF (version M23) were used to generate the output count matrix.
Slice preparation and electrophysiology
Brain slices were prepared from the mouse CN as previously described with a slight modification183,184,185. Briefly, P22-28 mice were deeply anesthetized with 3% isoflurane and then immediately decapitated. The brain was removed and placed in the iced oxygenated NMDG solution (for the recipe, see above). The brain tissue containing CN was sliced into 200–300 µm thick sections parasagittally in the NMDG solution with a microtome (Leica, VT1200). The slices were transferred into the oxygenated NMDG solution (34 ± 0.5 °C) for 10 min and then incubated in the artificial cerebrospinal fluid (ACSF, 119 mM NaCl, 2.5 mM KCl, 1 mM NaH2PO4, 25 mM NaHCO3, 1 mM MgCl2, 25 mM glucose and 2 mM CaCl2, pH 7.4) at 34 ± 0.5 °C for 40–60 min before recording.
Whole-cell recordings were obtained from CN neurons as previously described185. Briefly, borosilicate glass pipettes (3–5 MΩ) were pulled with micropipette pullers (P-1000, Sutter) and filled with intracellular solution containing 120 mM potassium gluconate, 10 mM HEPES, 4 mM KCl, 4 mM MgATP, 0.3 mM Na3GTP, 10 mM sodium phosphocreatine, and 0.5% biocytin (pH 7.25). Whole-cell recordings were performed at 32 ± 0.5 °C with the EPC 10 amplifiers (HEKA Electronics, Lambrecht, Germany). PatchMaster (HEKA) was used to operate the recording system and record the electrophysiology data. The membrane potential response of each neuron to increasing current steps (600 ms) every 1 s was digitized at 25 kHz. To compare the intrinsic properties and firing pattern across cell types, their membrane responses to a hyperpolarizing current step, to the current step at the rheobase, and to a current step at 2X rheobase were recorded and shown for each cell type (Figs. 2–5). Recorded neurons were then fixed for post hoc morphological recovery.
Patch-seq
Patch-seq was performed on CN neurons following our published protocol186. The RNA was extracted from each neuron upon completion of whole-cell recordings with a modified internal solution (111 mM potassium gluconate, 4 mM KCl, 10 mM HEPES, 0.2 mM EGTA, 4 MgATP, 0.3 Na3GTP, 5 mM sodium phosphocreatine, 0.5% biocytin and 0.48 units/µL RNase inhibitor, pH 7.25). Recorded neurons were then fixed for post hoc morphological recovery. The RNA was reverse transcribed with Superscript II Reverse Transcriptase (Invitrogen). Sequences of primers used for cDNA generation and amplification were shown in Supplementary Data 9. The cDNA samples that passed quality control were used to generate sequencing libraries using the Nextera XT DNA Library Preparation Kit (Illumina, FC-131-1096), following the user guide. Each library was sequenced at a depth of 1–2 million pair-end 150 bp reads on the Illumina NovaSeq 6000 or Hiseq 4000 platform. Raw reads were aligned to the Mus musculus genome (GRCm38) with annotation GTF (version M23) using STAR 2.7.7a187, and an expression matrix was generated with FeatureCounts188.
RNA fluorescence in situ hybridization
Wild type or Penk-Cre:Ai9 mice at P22-28 were deeply anesthetized with 3% isoflurane and decapitated immediately, and their brains were immediately removed from the skull and embedded in optimal cutting temperature (OCT, Sakura) compound on dry ice. The brain blocks were then sagittally sectioned into 25 µm thick slices. RNA fluorescence in situ hybridization (FISH) was performed on sections containing the CN at the RNA In Situ Hybridization Core at Baylor College of Medicine, using a previously described in situ hybridization method with slight modifications189. Briefly, digoxigenin (DIG) and fluorescein isothiocyanate (FITC) labeled mRNA antisense probes were first synthesized from reverse-transcribed mouse cDNA. Following fixation and acetylation, the two probes were then simultaneously hybridized to the brain sections. Following a series of washing and blocking steps, DIG-labeled probes were visualized with tyramide-Cy3 Plus, and FITC-labeled probes were visualized with tyramide-FITC Plus. The brain sections were then mounted with Prolong Diamond (Invitrogen). Images were captured at 20× magnification using LSM 710 or 880 confocal microscope (Zeiss). Typical FISH images were exported for the final layout with ZEN 2.3 (Zeiss). Primer sequences for the probes were derived from Allen Brain Atlas (http://www.brain-map.org). Sequences of primers used for generating the FISH probes were shown in Supplementary Data 9.
Immunofluorescence staining
GlyT2-EGFP mice at P22-28 were euthanized with 3% isoflurane in fume hood and then perfused transcardially with 20 mL PBS followed by 20 mL of 4% PFA in 0.1 M PB. The brain was removed, post-fixed in 4% PFA for 1 day, and then sagittally sectioned into 50 µm thick slices in PBS. The slices were washed three times in PBS before blocking with 10% goat serum and 1% Triton X in PBS for 1 hour. The slices were incubated first with mouse anti-PHGDH (Invitrogen, PA5-54360, 1:400) or rabbit anti-NECAB2 (Sigma-Aldrich, HPA013998, 1:500) as primary antibodies, and then incubated with Alexa-Fluor goat anti-mouse 633 (Invitrogen, A-21052) or goat anti-rabbit Alexa-Fluor 647 (Invitrogen, A-21244) as secondary antibodies for at least 1 h. Slices were then mounted with the mounting medium (Abcam, ab104139). To stain PHGDH or NECAB2 in recorded neurons, brain slices were immediately fixed after whole-cell recording with 4% PFA for 1 day. The slices were first incubated with primary antibodies (see above) for at least 1 h, and then incubated with Alexa Fluor 568 conjugates of streptavidin (Invitrogen, S11226) to visualize biocytin together with those secondary antibodies (see above) to visualize putative signals of PHGDH or NECAB2 in recorded neurons. Images were captured at 10× or 20× magnification using LSM 710 confocal microscope (Zeiss).
Morphological recovery
Morphological recovery of recorded neurons and light microscopic examination of their morphology were carried out following previously described methods185. In brief, upon the completion of whole-cell recordings, the slices were immediately fixed in 2.5% glutaraldehyde/4% paraformaldehyde in 0.1 M PB at 4 °C for 10–14 days and then processed with an avidin-biotin-peroxidase method with an ABC kit (Vector Laboratories, PK-4000) to reveal cell morphology. Recovered neurons were examined and reconstructed using a 100× oil-immersion objective lens and camera lucida system (Neurolucida, MicroBrightField). For morphological feature extraction, only cells with intact dendritic structures were included. Axons were excluded from this assessment as they are typically severed during slice preparation. We took special care to exclude reconstructions of the neurons that showed any signs of dendritic damage (due to the slicing procedure), or poor overall staining due to the aspiration procedure involved in Patch-seq (invisible or unclear soma and dendrite, likely caused by the collapse of the soma during nucleus extraction and subsequent leakage of the biocytin).
Computational analysis of snRNA-seq data
For snRNA-seq data, we used CellRanger (version 5.0.1) to generate a barcode table, gene table, and gene expression matrix. Downstream analysis and visualization were performed using Scvi-tools and SCANPY190. For preprocessing, we followed the standard protocols used in the field191. Specifically, nuclei with >5% UMIs mapped to mitochondrial genes and those with less than 200 genes detected were excluded from the analysis. We removed doublets sample-by-sample using Scrublet192. Doublet score and doublet predictions histogram of each sample were manually inspected, adjusted, and then concatenated. A putative single nucleus with a doublet score exceeding the 95th percentile was removed from the downstream analysis. Those nuclei co-expressing multiple canonical marker genes for different cell types as follows were also considered as doublets and were excluded for further analysis: Mobp/Mog/Mag (ODC), Gabra6 (granule cell), Ptprc/Csf1a/C1qa (microglia), Slc1a2/Aqp4/Gja1 (astrocyte), Dcn (fibroblast), Gad1/Gad2/Slc6a5 (inhibitory neurons), Flt1/Cldn5 (endothelial). Counts of all the nuclei were normalized by the library size multiplied by 10,000 and then logarithmized. The top 6000 highly variable genes, which were identified by dispersion-based methods, were used for principal component analysis. Leiden graph-clustering method and UMAP were used for clustering and visualization.
Due to the proximity of the CN to the cerebellum (CB), paramedian lobule (dorsal to the CN), to the principal sensory nucleus of the trigeminal nerve (PSN; ventral to CN), and to the dentate nucleus, our microdissection inevitably includes more or less of these adjoining tissues. To remove potential CB-derived nuclei from our dataset (Fig. S2), we compared our data to a recently published CB snRNA-seq dataset21. By examining the expression of marker genes for each CB cell type in our snRNA-seq data (Supplementary Data 1), we identified several putative neuronal and non-neuronal CB clusters in our dataset. Firstly, two clusters show clear molecular signatures of Bergmann cells and Purkinje cells respectively (Supplementary Data 1), indicating they are contaminated nuclei from these two CB-specific cell types (Fig. S2a, b). Secondly, two large clusters assigned to granule cells (GCs, i.e., enriched with the canonical marker gene Gabra6, Supplementary Data 1) could be discriminated by the expression levels of Grm4 and Col27a1 (Fig. S2c). Based on the molecular signatures of GCs in CB (Supplementary Data 1) and Allen Brain Atlas ISH data (Fig. S2c), we concluded that Grm4−/Col27a1+ cluster corresponds to GCs from CN, while the Grm4+/Col27a1− cluster corresponds to GCs from CB (CB_Granule, Fig. S2a). After removing these CB nuclei from our dataset, we computationally isolated the neuronal population (except for GCs and UBC) and then performed subclustering analysis to reveal distinct neuronal clusters (Fig. S2d). One neuronal cluster enriched with Htr2c and two clusters enriched with Necab2 were first identified (Fig. S2e, f). As shown on Allen Brain Atlas (Fig. S2e), Htr2c expression is restricted to the dentate nucleus, and thus this Htr2c-enriched cluster in our dataset is from the dentate nucleus. Two Necab2+ clusters are either Tnnt1+ or Tnnt1−, and only the Tnnt1+ nuclei are from the CN while Tnnt1− nuclei are from the PSN, based on Allen ISH data (Fig. S2e, f) and our FISH data (Fig. S7b5).
We then identified two pure GABAergic clusters (Gad1+/Slc6a5−) enriched with Tfap2b (Fig. S2g), reminiscent of MLI1 and MLI2 from the CB (Supplementary Data 1)21. Since pure GABAergic interneurons in the CN are absent or rare (Fig. S2h), we concluded these two clusters are indeed MLI1 and MLI2 from the CB. We also identified two Tfap2b-enriched glycine/GABA combinatorial clusters reminiscent of Golgi cells21, which were either Scrg1+ or Scrg1− (Fig. S2g). We concluded that the Scrg1+ cluster was Golgi cells in the CB (CB_Golgi) based on their marker gene in CB (Supplementary Data 1)21 and our FISH data (Fig. S2i); only the Scrg1− cluster corresponds to auditory Golgi cells (CN_Golgi, Figs. S2i and S5). We also identified a glycine/GABA combinatorial cluster enriched with both Klhl1 and Slc36a2 (Fig. S2g), marker genes of Purkinje layer interneurons (PLIs) from the CB (Supplementary Data 1)21. We conclude that this indeed corresponds to PLIs from the CB based on the expression patterns of Klhl1 and Slc36a2 across CN and CB (Fig. S2j).
The potential inclusion of non-neuronal nuclei from adjacent regions (with the exception of Bergmann glia) in our dataset was not addressed, since non-neuronal populations are not the focus of our analysis. Thus, the non-neuronal nuclei as shown in Fig. S1b may include those from adjoining regions. The UBC population in our dataset may include CB-derived UBCs, but we believed that this contamination, if any, was minimal. Firstly, using the ratio of the UBC population versus GC population in the CB (~1: 300)21, we estimated ~80 UBCs from CB might be included in our dataset (based on 24,118 CB-derived GCs in our dataset). In addition, the region of the CB close to the CN (i.e., CB paramedian lobule) has a smaller population of UBCs compared to other CB regions. We thus concluded that out of 3653 UBCs in our dataset, only a very small number may be UBCs from the CB, which would not significantly affect our analysis.
After the removal of the nuclei from adjoining tissues, the remaining 61,211 nuclei were used for further clustering analysis, and each cluster and cell type were annotated with the commonly used marker genes as listed in Supplementary Data 1 (including the marker genes for CB cell types). Calculation of the local inverse Simpson’s index193 demonstrated that these populations were well mixed and contained cells from each sample, indicating successful integration (Fig. S3). Our initial clustering combined the single-nucleus expression data from male (n = 31,115 nuclei) and female samples (n = 13,140 nuclei); each cluster comprised cells from both sexes, indicating that there were few sex-dependent differences (Fig. S3). The data from both sexes were thus pooled for subsequent analyses.
Patch-seq data analysis
In total, we performed Patch-seq from 438 cells (323 excitatory neurons, 115 inhibitory neurons, from 69 mice) that passed initial quality control. Among these sequenced 438 cells, 387 neurons (293 excitatory neurons, 94 inhibitory neurons) passed further quality control, yielding 2.40 ± 0.08 million (mean ± s.e.) reads and 8025 ± 70 (mean ± s.e.) detected genes per cell (Fig. S3). Patch-seq data were clustered and visualized following a similar method with SCANPY. Cells with a mapping rate of <60% and gene features of <4000 were not included in the analysis. We used the batch-balanced KNN method for batch correction194. Patch-seq data were mapped to the snRNA-seq data following a similar protocol as used previously28,96. Initially, we utilized the count matrix from 10x Genomics snRNA-seq data to identify the 3000 most variable genes through a dropout-based feature selection method195. For each gene, we calculated the fraction of near-zero counts (τ) and the mean log non-zero expression (μ) to establish a threshold for variability. μ and τ roughly followed the equation:\(\,\tau=\exp (-1.5\mu+0.002)\). An iterative approach was employed to determine a threshold based on the relationship between μ and τ, allowing us to select the desired number of variable genes. We then log-transformed all counts with log2(x + 1) transformation and averaged the log-transformed counts across all cells in each of the identified clusters in the snRNA-seq dataset to obtain reference transcriptomic profiles of each cluster (N × 3000 matrix). Out of these 3000 genes, 2575 are present in the mm10 reference genome that we used to align reads in our Patch-seq data. We applied the same log2(x + 1) transformation to the read counts of Patch-seq cells, and for each cell, computed Pearson correlation across the 2575 genes with all clusters. Each Patch-seq cell was assigned to a cluster to which it had the highest correlation. To illustrate the mapping results, each Patch-seq cell was positioned at the median UMAP coordinates of its ten nearest neighbors in snRNA-seq UMAP embeddings, as determined by Pearson correlation28,114.
DEG analysis and marker gene identification
To identify the DEGs for clusters and subclusters, we used SCANPY to rank genes using the T-test_overestim_var test with Benjamini–Hochberg correction. SCANPY generated logFC (with the base as 2) and FDR (False discovery rate) corrected P value and DEGs were identified based on p-value < 0.01 and |log2FC| > 1. This method could detect more DEGs in the clusters that have large numbers of cells, while in relatively small cell populations only those with much higher fold changes could be identified as DEGs.
Electrophysiological feature extraction and analysis
All cells that passed the electrophysiology data quality were included in the analysis. Electrophysiological data were analyzed with a python-based GUI named PatchView that we have developed196 or a custom-made IDL code. To access the IDL code, the raw electrophysiological data was exported to ASCII format using Nest-o-Patch. To highlight the differences in the intrinsic properties and firing patterns, the membrane responses to a hyperpolarizing current step, to the current step at the rheobase, and to a current step at 2X rheobase were shown for each cell type (Figs. 2–5). Notably, the values reported in this study were not corrected for the liquid junction potential, which was calculated to be ~15.8 mV based on the composition of the internal solution and ACSF. 20 electrophysiological features were extracted from the electrophysiological traces as outlined below, following a method similar to that described in previous studies (Fig. S14)96.
To calculate the time constant (Tau), all the traces in response to negative current steps (from the onset of the current step to the minimal potential during the stimulation period) were fitted with an exponential function as follows:
Here, f(t) is the voltage at the time t, τ is the membrane time constant, a and C are two constants. The τ was calculated for each trace. Boxplot was used to remove the outliers and the mean averaged across all traces after outlier removal was used as the time constant.
We calculated the sag ratio and rebound voltage using the hyperpolarizing trace elicited by the maximal negative current injection. The sag ratio was defined as the difference between the sag trough of the trace and the resting membrane potential (RMP) (ΔV1), divided by the difference between steady-state membrane voltage and the RMP (ΔV2). The trace from the stimulation offset to the peak of the depolarizing trace following the offset was fitted with an exponential function, and the potential difference between the maximal value of the fitting curve and the RMP was defined as the rebound value.
To calculate the input resistance and RMP, we measured the steady-state membrane potentials (determined by the average values of the last 100 ms of the hyperpolarization) in the traces elicited by the first five negative-going currents in the step protocol, and then plotted these values (y-axis) with each of the corresponding currents (x-axis). We fit the plot with a linear function shown as follows:
Here, f(c) steady-state voltages, c is the injected currents, Rm and Vrmp are constants that represent the slope of the linear function and the potential at the y-axis intersection, corresponding to the input resistance and the RMP respectively.
Rheobase is defined as the minimum current that could elicit any action potentials (APs) during the step-current injection. To extract the properties of action potential (AP), we used the first AP elicited by our step stimulation protocol. To calculate the threshold of the AP, the trace within 20 ms preceding the AP peak was used to calculate the third derivative and the data point with the maximal third derivative value was defined as the AP threshold197. The AP amplitude is the difference between the threshold potential and the peak potential of the AP. The AP half-width was defined as the time difference between the voltage at AP half-height on the rising and falling phase of the AP. The afterhyperpolarization (AHP) was defined as the potential difference between the AP threshold and the minimal value of the trough after the AP. The time difference from the stimulation onset to the first AP’s threshold at the rheobase sweep was defined as the AP delay time. The depolarization time was the time difference from the AP threshold to the overshoot. The time from the overshoot to the repolarizing voltage that equaled the threshold was the AP’s repolarization time. Spike-frequency adaptation was quantified with the membrane potential trace in response to the injection of the step current at 2X Rheobase. The interspike interval (ISI) between the first two APs divided by the ISI between the second and the third APs was the initial adaptation index. The ISI between the first two APs divided by the ISI between the last two APs was the last adaptation index.
The features used for E-type clustering analysis and UMAP embedding calculation are shown in Fig. S4. The features were standardized using StandardScaler from scikit-learn Python library (version 1.3.2). UMAP embeddings were generated from the standardized electrophysiological features using the umap-learn Python library (version 0.5.3) (Fig. 2c, Fig. 5c, Fig. S4b, f, h). K-means clustering analysis was performed on standardized electrophysiological features (using KMeans from scikit-learn library) to identify inherent groupings within the dataset. The results of this analysis were visualized on UMAP embeddings (Fig. S4b, h). A confusion matrix was used to show the performance of K-means clustering in differentiating cell types as labeled manually (Fig. S4b, h). To include the recording ___location (either VCN or DCN) as an additional feature in the inhibitory neuron clustering analysis, inhibitory neurons recorded in the VCN were assigned a value of 0, while those recorded in the DCN were assigned a value of 1. These values were then incorporated alongside the electrophysiological values for K-mean clustering analysis and to generate UMAP embedding (Fig. S4h and Fig. 5d). Different value assignments were tested and did not impact clustering analysis.
To determine whether tdTomato+ cells in Penk-Cre mice aligned with the electrophysiological profile of vertical cells (Fig. 5l), we extracted the electrophysiological features of those tdTomato+ cells recorded from Penk-Cre mice, and these features were then used to cluster tdTomato+ cells together with all other recorded inhibitory neurons (Fig. 5l). Visualizing tdTomato+ cells within the resulting UMAP embeddings revealed their relationship (co-clustering) with the population of recorded vertical neurons (Fig. 5l).
Morphological reconstruction and analysis
All cells that passed the morphological criteria were reconstructed and included in the analysis. Neuronal morphologies were reconstructed with Neurolucida (MicroBrightField) and analyzed with Neurolucida Explorer (including Sholl analysis of the dendritic arbors and the soma shape analysis). For visualization (Figs. 2a, 5a), reconstructions were imported into Canvas X (Canvas GFX). The CN contours for each morphology (Figs. 2a, 5a, S6d, S6h) were manually drawn using the drawing tools available in Canvas X. Other elements and cartoon within Fig. 1a and Fig. S1a were also created using Canvas X. For more comprehensive morphological feature analysis, each morphology file was converted into the SWC format with NLMorphologyConverter 0.9.0 (http://neuronland.org) for feature extraction using MorphoPy (https://github.com/berenslab/MorphoPy). See https://github.com/bcmjianglab/ cn_project/tree/main/patchseq/M-clusters for a detailed description of the morphological features.
The features used for M-type clustering analysis and UMAP embedding calculation are shown in Fig. S4. UMAP embeddings were generated from standardized morphological features using the umap-learn Python library (version 0.5.3) (Fig. 2d, Fig. 5c, Fig. S4b, d). K-means clustering analysis was performed on these features as did on electrophysiological features and the results were visualized on UMAP embeddings (Fig. S4b, d). The confusion matrix was used to show the performance of K-means clustering (Fig. S4b, d).
To determine whether tdTomato+ cells in Penk-Cre mice aligned with the morphological profile of vertical cells (Fig. 5l), we extracted the morphological features of those tdTomato+ cells recorded from Penk-Cre mice, and these features were then used to cluster tdTomato+ cells together with all other recorded inhibitory neurons (Fig. 5l). Visualizing tdTomato+ cells within the resulting UMAP embeddings revealed their relationship (co-clustering) with the population of recorded vertical neurons (Fig. 5l).
Sparse reduced-rank regression and simple linear correlation and regression
Sparse reduced-rank regression (RRR) was performed as previously reported96. We used 14 electrophysiological features extracted from 293 Patch-seq excitatory neurons (Fig. 2e) to predict the electrophysiological features from gene expression. Interneurons were excluded from the analysis due to data insufficiency. To enhance interpretability, we only focused on voltage-sensitive ion channel genes and 119 genes (source data) out of 330 genes in this category (https://www.genenames.org/) were chosen for the analysis (those expressed in less than 10 of the Patch-seq neurons were excluded). We optimized the hyperparameter combinations based on the model’s predictive performance (measured by cross-validated R-squared), and chose a model with rank = 5, zero ridge penalty (α = 1), and regularization strength (λ = 0.28) tuned to yield a selection of 25 genes, which achieved near-maximal predictive accuracy (Fig. S12b). Two biplots (bi-biplots) were used to show the RRR analysis results96, where the directions and magnitudes of lines representing individual genes and electrophysiological features reflect their correlations to two underlying components. Only features with a correlation above 0.4 are shown in the bibiplots. By comparing these directions and magnitudes between the two biplots, potential associations between specific genes and electrophysiological traits can be inferred.
To provide an additional, more direct assessment of the relationship between each ion channel gene and the electrophysiological features, the simple linear correlation and regression (including effect size and significance) between each of ion channel genes (n = 119, including 25 gene selected by the sparse RRR model) and the electrophysiological features were independently calculated (Fig. S12c). The correlation coefficients, significance and linear regression lines are visualized in dot plots and scaled plots (Fig. S12c–f). For all ion channel genes, only the top 25 with the highest correlation coefficients are displayed, and most of these overlap with the genes selected by the sparse RRR model. This substantial overlap serves as compelling validation for the accuracy and effectiveness of the sparse RRR model.
Registering the anatomic ___location of cells
To register each Patch-seq cell to the CN 3-dimensional (3D) ___location, we first generated a 3D CN reference space based on the Allen Mouse Brain Atlas and then aligned each brain slice used for our Patch-seq recordings to this reference space. We reconstructed a contour on each slice that delineates the entire CN area in that slice, and then iteratively searched for the contour within the 3D CN reference space that best matched the reconstructed contour (i.e., estimating the 3D ___location and orientation of that slice). Since our slices were cut in the para-sagittal plane, we limited our grid search parameters range: pitch angle (0, −25 rad) and roll angle (0, −20 rad), median to lateral shift (0, 850 µm), in-plane orientation (−90, 90). For each parameter set, we calculated the overlapping area of the reconstructed contour with the corresponding contour in the 3D CN reference space. The parameters that result in a maximal overlapping area were used as the final parameter for estimation. We visually inspected the mapping results to rule out erroneous registration. For some ambiguous slices (i.e., having very small CN regions), we further limited the search parameter range to improve the accuracy. The ___location of neurons in that slice was then projected into the 3D CN reference space. The 3D brain model with the entire CN (Fig. 1a) was generated using BrainRender198 based on the Allen Mouse Brain Atlas.
Machine learning algorithms to validate expert classification
Expert classification was performed by four individuals (TN, LT, JJ, MM), all with extensive expertise in studying CN cell types. Each individual independently assessed each cell based on its ___location, morphology, input resistance, and responses to a family of depolarizing and hyperpolarizing current steps. The results of these independent evaluations were then compared to established features in the CN literature4,5,6. Any disagreements were resolved through discussions in group meetings until a consensus identification was reached for each recorded cell. A random forest classifier based on electrophysiological or morphological features was then used to validate expert classification. Briefly, the electrophysiological or morphological features of each neuron were extracted, normalized, and scaled. To include the recording ___location (either VCN or DCN) within electrophysiological features during the classifier training process (for inhibitory neuron classification), inhibitory neurons recorded in the VCN were assigned a value of 0, while those recorded in the DCN were assigned a value of 1 (Fig. S4g). Different value assignments were tested and did not impact classifier performance. The data were split into training and testing datasets randomly, and the classifier was trained with the training data. The accuracy in distinguishing the cell types as labeled manually by the classifier was evaluated with the testing dataset. The accuracy varied with different training datasets used, so we trained the classifier 10,000 times, each with a different training dataset. The classifier settings with the top 100 performance outcomes were used to weigh the electrophysiological or morphological features for cell type discrimination. The overall accuracy in discriminating cell types by the classifier was reported as the mean accuracy within the chosen settings. The top 15 most important features contributing to the performance were used for UMAP visualizations (Fig. S4a, 4c,4e, 4g). A confusion matrix was used to show the overall accuracy in distinguishing the cell types as labeled manually by the classifier (Figs. 2c-d, 5c, d, S4f).
MetaNeighbor analysis and gene expression specificity analysis
To identify the gene families or sets whose differential expression could robustly predict cell types, MetaNeighbor analysis was conducted on our snRNA-seq dataset (either the whole dataset, projection neurons or inhibitory neurons only) utilizing pyMN tool, following established user tutorials199. In brief, we split our dataset into two halves and aimed to measure the replicability of cell type identity between them when considering only a given gene set or family. Cell type identities were held back from one set at a time and predicted from the other by neighbor voting based on a Spearman correlation subnetwork built from the expression of the gene set (or family) being tested. The accuracy of recovering cells of the same cell types was evaluated through cross-validation and reported as AUROC (Area Under Receiver-Operator Characteristics Curve) values. We then ranked the gene sets or families by the mean AUROC value. We first performed this analysis on gene sets annotated with mouse Gene Ontology (GO) terms (obtained via pyMN). To identify more specific gene categories, we also applied the test to all 620 gene families annotated in the HUGO Gene Nomenclature Committee (HGNC) database (https://www.genenames.org/cgi-bin/genegroup/download-all). Furthermore, we curated 342 cell adhesion molecule (CAM) genes or CAM-modifying enzyme genes into 12 families (Supplementary Data 5), following the strategy by Paul et al., 201754. We conducted additional tests on these curated families. Supplementary tests and AUROC scoring were also performed on 70 transcriptional factor families (1611 mouse transcription factors) sourced from AnimalTFDB4 (https://guolab.wchscu.cn/AnimalTFDB4/), the most comprehensive animal transcription factor database with precise identification and thorough annotation61. AUROC values for each family within these gene sets were documented in Supplementary Data 5. The gene sets or families with AUROC scores >0.75 (regarded as a stringent threshold) are considered as being discriminating54.
To identify cell type-specific gene expression patterns, we applied the tspex Python package (version 0.6.2) to our snRNA-seq dataset to calculate gene expression specificity (z-score) for each cell type200. We focused on cell adhesion molecules (CAMs) and transcription factors (TFs), identifying 135 CAMs and 299 TFs with a specificity z-score > 0.75 (Supplementary Data 6). Since CN neurons are comprised of three major classes arising from distinct lineages -projection neurons, inhibitory neurons, and granule cells/unipolar brush cells (UBCs)62,151,205 -we also calculated gene expression specificity within each of these major neuronal classes for TF and CAMs (Fig. 6)81,132,147.
Synergistic core identification
Based on the notion that cell type identity can be determined by a combination of a few key TFs that exhibit synergistic transcriptional regulation, we used a computational platform that performs a dynamic search for an identity core composed of TFs that possess an optimal synergistic coexpression pattern using an information-theoretic measure of synergy computed from the snRNA-seq dataset72. For each target subpopulation, a set of TFs that are preferentially expressed in the target cell type (i.e., specific TFs) is identified. Then most synergistic subset of specific TFs is sought by computing multivariate mutual information (MMI). Once a most synergistic core is found, it is extended by computing MMI with TFs expressed in both target and background cell types (non-specific TFs). Each TF core thus formed by a set of specific TFs and a few non-specific TFs given that non-specific TFs also contribute to the synergistic expression specifying cell type identity72. We performed the synergistic core analysis on principal projection neurons or inhibitory neurons using default parameters72. The synergistic TF cores for each cell type were reported in Supplementary Data 7.
FISH image analysis and quantification
Confocal images were captured from FISH-stained brain slices using LSM 710 or 880 confocal microscopes (Zeiss), and the image files (.lsm) containing image stacks were loaded into IDL for analysis of the distribution and co-localization of various mRNA transcripts. From each slice, the images were generated to capture either green fluorescence signals (FITC) or red signals (DIG). The maximum projected image was filtered with a Gaussian filter to remove background signals. To detect a cell with expression signals, cell regions were segmented using a customized IDL code (Fig. S15). A cell region with an area smaller than 80 µm2 or larger than 800 µm2 was not counted as a cell. A cell region with a circularity of less than 0.4 was not counted as a cell either. The circularity of a cell was calculated using the function: \({C}=4\pi {a}^{2}/{p}^{2}\). Here, C is the circularity, a is the area of a cell region and p is the perimeter of the cell region. Cell regions outside the CN area were not included for further analysis. The spatial position of a cell was set as the median of the x or y positions of the pixels within a detected cell region. The fluorescence intensity (F) of a detected cell (either green or red) was set as the mean intensity of all pixels within a detected cell region. After removing fluorescence intensity values within a cell region, the images of each channel were then smoothed by a 60 µm length smoothing window. The smooth images were counted as the background signals of each channel. The background signals (F0) of a detected cell were set as the mean values of the background intensities within a cell region. The intensity differences (ΔF) between F and F0 divided by F0 of a detected cell from both channels (green and red) were used to generate a 2-D scatter plot in Python, with each axis representing either channel. From this scatter plot, we used a method similar to the Flow Cytometry gated analysis to determine whether a cell was single-labeled or double-labeled (Fig. S15). The number of single-labeled and double-labeled cells was counted across 5–8 consecutive sagittal sections of CN. The distribution of labeled cells (single or double) across CN was quantified through the A-to-P and D-to-V axis with Jointplot from Seaborn python package. See https://github.com/bcmjianglab/cn_project/tree/main/FISH_analysis for more details about FISH image analysis.
ISH images in Fig. S8 were acquired from the Allen Mouse Brain Atlas (www.mouse.brain-map.org/)201. Single discriminatory markers were shown for each cell type, with several ISH images being repeated. ISH images were acquired with minor contrast adjustments as needed to maintain image consistency.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Original electrophysiological, morphological, processed RNA-sequencing data, and FISH/immunostaining imaging supporting the findings of this study have been deposited to the Mendeley Data (https://doi.org/10.17632/2n8m8k6b2g.1). The raw snRNA-seq and Patch-seq datasets have been deposited in the NCBI database with GEO accession number: GSE273145. Source data are provided with this paper.
Code availability
The code associated with this study is available at zenodo (https://doi.org/10.5281/zenodo.14036008), which links to a GitHub repository (https://github.com/bcmjianglab/cn_project).
References
Oertel, D., McGinley, M. & and Cao, X.-J. in Encylopedia of Neuroscience (Springer, 2009).
Grothe, B., Pecka, M. & McAlpine, D. Mechanisms of sound localization in mammals. Physiol. Rev. 90, 983–1012 (2010).
Chi, T., Ru, P. & Shamma, S. A. Multiresolution spectrotemporal analysis of complex sounds. J. Acoust. Soc. Am. 118, 887–906 (2005).
Rhode, W. S. & Greenberg, S. in The Mammalian Auditory Pathway: Neurophysiology (eds. Arthur N. Popper & Richard R. Fay) 94–152 (Springer, 1992).
Palmer, A. R. Physiology of the cochlear nerve and cochlear nucleus. Br. Med. Bull. 43, 838–855 (1987).
Yin, T. C. T., Smith, P. H. & Joris, P. X. Neural mechanisms of binaural processing in the auditory brainstem. Compr. Physiol. 9, 1503–1575 (2019).
Doucet, J. R. & Ryugo, D. K. Structural and functional classes of multipolar cells in the ventral cochlear nucleus. Anat. Rec. Part A Discov. Mol. Cell. Evolut. Biol. 288, 331–344 (2006).
Blackburn, C. C. & Sachs, M. B. Classification of unit types in the anteroventral cochlear nucleus: PST histograms and regularity analysis. J. Neurophysiol. 62, 1303–1329 (1989).
Hancock, K. E. & Voigt, H. F. Intracellularly labeled fusiform cells in dorsal cochlear nucleus of the gerbil. I. Physiological response properties. J. Neurophysiol. 87, 2505–2519 (2002).
Ngodup, T., Romero, G. E. & Trussell, L. O. Identification of an inhibitory neuron subtype, the L-stellate cell of the cochlear nucleus. Elife 9, e54350 (2020).
Xie, R. & Manis, P. B. GABAergic and glycinergic inhibitory synaptic transmission in the ventral cochlear nucleus studied in VGAT channelrhodopsin-2 mice. Front. Neural Circuits 8, 84 (2014).
Zheng, G. X. et al. Massively parallel digital transcriptional profiling of single cells. Nat. Commun. 8, 14049 (2017).
Macosko, E. Z. et al. Highly parallel genome-wide expression profiling of individual cells using nanoliter droplets. Cell 161, 1202–1214 (2015).
Saunders, A. et al. Molecular diversity and specializations among the cells of the adult mouse brain. Cell 174, 1015–1030.e1016 (2018).
Cadwell, C. R. et al. Electrophysiological, transcriptomic and morphologic profiling of single neurons using Patch-seq. Nat. Biotechnol. 34, 199–203 (2016).
Gates, T. S., Weedman, D. L., Pongstaporn, T. & Ryugo, D. K. Immunocytochemical localization of glycine in a subset of cartwheel cells of the dorsal cochlear nucleus in rats. Hear Res. 96, 157–166 (1996).
Hancock, K. E. & Voigt, H. F. Intracellularly labeled fusiform cells in dorsal cochlear nucleus of the gerbil. II. Comparison of physiology and anatomy. J. Neurophysiol. 87, 2520–2530 (2002).
Varecka, L., Wu, C.-H., Rotter, A. & Frostholm, A. GABAA/benzodiazepine receptor α6 subunit mRNA in granule cells of the cerebellar cortex and cochlear nuclei: Expression in developing and mutant mice. J. Comp. Neurol. 339, 341–352 (1994).
Campos, M. L., de Cabo, C., Wisden, W., Juiz, J. M. & Merlo, D. Expression of GABAA receptor subunits in rat brainstem auditory pathways: cochlear nuclei, superior olivary complex and nucleus of the lateral lemniscus. Neuroscience 102, 625–638 (2001).
Englund, C. et al. Unipolar brush cells of the cerebellum are produced in the rhombic lip and migrate through developing white matter. J. Neurosci. 26, 9184–9195 (2006).
Kozareva, V. et al. A transcriptomic atlas of mouse cerebellar cortex comprehensively defines cell types. Nature 598, 214–219 (2021).
Guo, C., Huson, V., Macosko, E. Z. & Regehr, W. G. Graded heterogeneity of metabotropic signaling underlies a continuum of cell-intrinsic temporal responses in unipolar brush cells. Nat. Commun. 12, 5491 (2021).
Borges-Merjane, C. & Trussell, L. O. ON and OFF unipolar brush cells transform multisensory inputs to the auditory system. Neuron 85, 1029–1042 (2015).
Irie, T., Fukui, I. & Ohmori, H. Activation of GIRK channels by muscarinic receptors and group II metabotropic glutamate receptors suppresses Golgi cell activity in the cochlear nucleus of mice. J. Neurophysiol. 96, 2633–2644 (2006).
Yaeger, D. B. & Trussell, L. O. Single granule cells excite Golgi cells and evoke feedback inhibition in the cochlear nucleus. J. Neurosci. 35, 4741–4750 (2015).
Manis, P. B., Kasten, M. R. & Xie, R. Classification of neurons in the adult mouse cochlear nucleus: linear discriminant analysis. PLoS One 14, e0223137 (2019).
Cadwell, C. R. et al. Cell type composition and circuit organization of clonally related excitatory neurons in the juvenile mouse neocortex. Elife 9, e52951 (2020).
Scala, F. et al. Layer 4 of mouse neocortex differs in cell types and circuit organization between sensory areas. Nat. Commun. 10, 4174 (2019).
McGinley, M. J., Liberman, M. C., Bal, R. & Oertel, D. Generating synchrony from the asynchronous: compensation for cochlear traveling wave delays by the dendrites of individual brainstem neurons. J. Neurosci. 32, 9301–9311 (2012).
Lu, H. W., Smith, P. H. & Joris, P. X. Mammalian octopus cells are direction selective to frequency sweeps by excitatory synaptic sequence detection. Proc. Natl Acad. Sci. USA 119, e2203748119 (2022).
Lauer, A. M., Connelly, C. J., Graham, H. & Ryugo, D. K. Morphological characterization of bushy cells and their inputs in the laboratory mouse (Mus musculus) anteroventral cochlear nucleus. PLoS One 8, e73308 (2013).
Romero, G. E. Central Circuitry Underlying The Medial Olivocochlear Efferent System (Ph.D. thesis) (Oregon Health & Science University School of Medicine, 2021).
Martin, M. R. Morphology of the cochlear nucleus of the normal and reeler mutant mouse. J. Comp. Neurol. 197, 141–152 (1981).
Webster, D. B. & Trune, D. R. Cochlear nuclear complex of mice. Am. J. Anat. 163, 103–130 (1982).
Cao, X. J., Shatadal, S. & Oertel, D. Voltage-sensitive conductances of bushy cells of the Mammalian ventral cochlear nucleus. J. Neurophysiol. 97, 3961–3975 (2007).
Manis, P. B. & Marx, S. O. Outward currents in isolated ventral cochlear nucleus neurons. J. Neurosci. 11, 2865–2880 (1991).
Rothman, J. S. & Manis, P. B. The roles potassium currents play in regulating the electrical activity of ventral cochlear nucleus neurons. J. Neurophysiol. 89, 3097–3113 (2003).
Cao, X. J. & Oertel, D. The magnitudes of hyperpolarization-activated and low-voltage-activated potassium currents co-vary in neurons of the ventral cochlear nucleus. J. Neurophysiol. 106, 630–640 (2011).
Wang, L. Y., Gan, L., Forsythe, I. D. & Kaczmarek, L. K. Contribution of the Kv3.1 potassium channel to high-frequency firing in mouse auditory neurones. J. Physiol. 509, 183–194 (1998).
Perney, T. M. & Kaczmarek, L. K. Localization of a high threshold potassium channel in the rat cochlear nucleus. J. Comp. Neurol. 386, 178–202 (1997).
Brew, H. M. & Forsythe, I. D. Two voltage-dependent K+ conductances with complementary functions in postsynaptic integration at a central auditory synapse. J. Neurosci. 15, 8011–8022 (1995).
Johnston, J. et al. Initial segment Kv2.2 channels mediate a slow delayed rectifier and maintain high-frequency action potential firing in medial nucleus of the trapezoid body neurons. J. Physiol. 586, 3493–3509 (2008).
Oertel, D., Wu, S. H., Garb, M. W. & Dizack, C. Morphology and physiology of cells in slice preparations of the posteroventral cochlear nucleus of mice. J. Comp. Neurol. 295, 136–154 (1990).
Xie, R. & Manis, P. B. Radiate and planar multipolar neurons of the mouse anteroventral cochlear nucleus: intrinsic excitability and characterization of their auditory nerve input. Front Neural Circuits 11, 77 (2017).
Doucet, J. R. & Ryugo, D. K. Projections from the ventral cochlear nucleus to the dorsal cochlear nucleus in rats. J. Comp. Neurol. 385, 245–264 (1997).
Kolston, J., Osen, K. K., Hackney, C. M., Ottersen, O. P. & Storm-Mathisen, J. An atlas of glycine- and GABA-like immunoreactivity and colocalization in the cochlear nuclear complex of the guinea pig. Anat. Embryol. 186, 443–465 (1992).
Bauer, C. K. & Schwarz, J. R. Ether-à-go-go K(+) channels: effective modulators of neuronal excitability. J. Physiol. 596, 769–783 (2018).
Zou, A. et al. Distribution and functional properties of human KCNH8 (Elk1) potassium channels. Am. J. Physiol. Cell Physiol. 285, C1356–C1366 (2003).
Rudy, B. & McBain, C. J. Kv3 channels: voltage-gated K+ channels designed for high-frequency repetitive firing. Trends Neurosci. 24, 517–526 (2001).
Apostolides, P. F. & Trussell, L. O. Chemical synaptic transmission onto superficial stellate cells of the mouse dorsal cochlear nucleus. J. Neurophysiol. 111, 1812–1822 (2014).
Saint Marie, R. L., Benson, C. G., Ostapoff, E. M. & Morest, D. K. Glycine immunoreactive projections from the dorsal to the anteroventral cochlear nucleus. Hear. Res. 51, 11–28 (1991).
Wickesberg, R. E. & Oertel, D. Delayed, frequency-specific inhibition in the cochlear nuclei of mice: a mechanism for monaural echo suppression. J. Neurosci. 10, 1762–1768 (1990).
Cant, N. B. in The Mammalian Cochlear Nuclei 91–105 (Springer, 1993).
Paul, A. et al. Transcriptional architecture of synaptic communication delineates GABAergic neuron identity. Cell 171, 522–539 e520 (2017).
Sun, S. et al. Hair cell mechanotransduction regulates spontaneous activity and spiral ganglion subtype specification in the auditory system. Cell 174, 1247–1263.e1215 (2018).
Shrestha, B. R. et al. Sensory neuron diversity in the inner ear is shaped by activity. Cell 174, 1229–1246.e1217 (2018).
Hobert, O. & Kratsios, P. Neuronal identity control by terminal selectors in worms, flies, and chordates. Curr. Opin. Neurobiol. 56, 97–105 (2019).
Arendt, D. et al. The origin and evolution of cell types. Nat. Rev. Genet. 17, 744–757 (2016).
Hobert, O. Terminal Selectors of Neuronal Identity. Curr. Top. Dev. Biol. 116, 455–475 (2016).
Deneris, E. S. & Hobert, O. Maintenance of postmitotic neuronal cell identity. Nat. Neurosci. 17, 899–907 (2014).
Shen, W.-K. et al. AnimalTFDB 4.0: a comprehensive animal transcription factor database updated with variation and expression annotations. Nucleic Acids Res. 51, D39–D45 (2022).
Shekhar, K. et al. Comprehensive classification of retinal bipolar neurons by single-cell transcriptomics. Cell 166, 1308–1323 e1330 (2016).
Yao, Z. et al. A high-resolution transcriptomic and spatial atlas of cell types in the whole mouse brain. Nature 624, 317–332 (2023).
Okawa, S. et al. Transcriptional synergy as an emergent property defining cell subpopulation identity enables population shift. Nat. Commun. 9, 2595 (2018).
Hnisz, D. et al. Super-enhancers in the control of cell identity and disease. Cell 155, 934–947 (2013).
Wilson, N. K. et al. Combinatorial transcriptional control in blood stem/progenitor cells: genome-wide analysis of ten major transcriptional regulators. Cell Stem Cell 7, 532–544 (2010).
Mazzoni, E. O. et al. Synergistic binding of transcription factors to cell-specific enhancers programs motor neuron identity. Nat. Neurosci. 16, 1219–1227 (2013).
Greig, L. C., Woodworth, M. B., Galazo, M. J., Padmanabhan, H. & Macklis, J. D. Molecular logic of neocortical projection neuron specification, development and diversity. Nat. Rev. Neurosci. 14, 755–769 (2013).
Colasante, G. et al. Rapid conversion of fibroblasts into functional forebrain GABAergic interneurons by direct genetic reprogramming. Cell Stem Cell 17, 719–734 (2015).
Huang, M. et al. Ptf1a, Lbx1 and Pax2 coordinate glycinergic and peptidergic transmitter phenotypes in dorsal spinal inhibitory neurons. Dev. Biol. 322, 394–405 (2008).
Chanda, S. et al. Generation of induced neuronal cells by the single reprogramming factor ASCL1. Stem Cell Rep. 3, 282–296 (2014).
Okawa, S. & Del Sol, A. A general computational approach to predicting synergistic transcriptional cores that determine cell subpopulation identities. Nucleic Acids Res 47, 3333–3343 (2019).
Ninkovic, J. et al. The transcription factor Pax6 regulates survival of dopaminergic olfactory bulb neurons via crystallin alphaA. Neuron 68, 682–694 (2010).
Rajakumari, S. et al. EBF2 determines and maintains brown adipocyte identity. Cell Metab. 17, 562–574 (2013).
Bobowski-Gerard, M. et al. Functional genomics uncovers the transcription factor BNC2 as required for myofibroblastic activation in fibrosis. Nat. Commun. 13, 5324 (2022).
Liu, H., Aramaki, M., Fu, Y. & Forrest, D. Retinoid-related orphan receptor beta and transcriptional control of neuronal differentiation. Curr. Top. Dev. Biol. 125, 227–255 (2017).
Chabbert, D. et al. Postnatal Tshz3 deletion drives altered corticostriatal function and autism spectrum disorder-like behavior. Biol. psychiatry 86, 274–285 (2019).
Vissers, J. H. A. et al. The scalloped and Nerfin-1 transcription factors cooperate to maintain neuronal cell fate. Cell Rep. 25, 1561–1576 e1567 (2018).
Parsons, MichaelJ. et al. The regulatory factor ZFHX3 modifies circadian function in SCN via an at motif-driven axis. Cell 162, 607–621 (2015).
Zhang, Z. et al. Zfhx3 is required for the differentiation of late-born D1-type medium spiny neurons. Exp. Neurol. 322, 113055 (2019).
Butts, J. C. et al. A single-cell transcriptomic map of the developing Atoh1 lineage identifies neural fate decisions and neuronal diversity in the hindbrain. Dev. Cell 59, 2171–2188 e2177 (2024).
Saul, S. M. et al. Math5 expression and function in the central auditory system. Mol. Cell Neurosci. 37, 153–169 (2008).
Cai, X. et al. Bhlhb5::flpo allele uncovers a requirement for Bhlhb5 for the development of the dorsal cochlear nucleus. Dev. Biol. 414, 149–160 (2016).
Farago, A. F., Awatramani, R. B. & Dymecki, S. M. Assembly of the brainstem cochlear nuclear complex is revealed by intersectional and subtractive genetic fate maps. Neuron 50, 205–218 (2006).
Di Bonito, M. & Studer, M. Cellular and molecular underpinnings of neuronal assembly in the central auditory system during mouse development. Front. Neural Circuits 11, 18 (2017).
Li, H. et al. Classifying drosophila olfactory projection neuron subtypes by single-cell RNA sequencing. Cell 171, 1206–1220 e1222 (2017).
de Wit, J. & Ghosh, A. Specification of synaptic connectivity by cell surface interactions. Nat. Rev. Neurosci. 17, 4–4 (2016).
Kolodkin, A. L. & Tessier-Lavigne, M. Mechanisms and molecules of neuronal wiring: a primer. Cold Spring Harb. Perspect. Biol. 3, a001727 (2011).
Rawson, R. L., Martin, E. A. & Williams, M. E. Mechanisms of input and output synaptic specificity: finding partners, building synapses, and fine-tuning communication. Curr. Opin. Neurobiol. 45, 39–44 (2017).
Kurusu, M. et al. A screen of cell-surface molecules identifies leucine-rich repeat proteins as key mediators of synaptic target selection. Neuron 59, 972–985 (2008).
Schroeder, A. et al. A modular organization of LRR protein-mediated synaptic adhesion defines synapse identity. Neuron 99, 329–344.e327 (2018).
Mutalik, S. P. & Gupton, S. L. Glycosylation in axonal guidance. Int. J. Mol. Sci. 22, 5143 (2021).
Trottein, F. et al. Glycosyltransferase and sulfotransferase gene expression profiles in human monocytes, dendritic cells and macrophages. Glycoconj. J. 26, 1259–1274 (2009).
Dani, N. & Broadie, K. Glycosylated synaptomatrix regulation of trans-synaptic signaling. Dev. Neurobiol. 72, 2–21 (2012).
Kobak, D. et al. Sparse reduced‐rank regression for exploratory visualisation of paired multivariate data. J. R. Stat. Soc. Ser. C 70, 980–1000 (2021).
Scala, F. et al. Phenotypic variation of transcriptomic cell types in mouse motor cortex. Nature 598, 144–150 (2021).
Bal, R. & Oertel, D. Hyperpolarization-activated, mixed-cation current (I(h)) in octopus cells of the mammalian cochlear nucleus. J. Neurophysiol. 84, 806–817 (2000).
Bal, R. & Oertel, D. Potassium currents in octopus cells of the mammalian cochlear nucleus. J. Neurophysiol. 86, 2299–2311 (2001).
Golding, N. L., Ferragamo, M. J. & Oertel, D. Role of intrinsic conductances underlying responses to transients in octopus cells of the cochlear nucleus. J. Neurosci.: Off. J. Soc. Neurosci. 19, 2897–2905 (1999).
Eatock, R. A., Xue, J. & Kalluri, R. Ion channels in mammalian vestibular afferents may set regularity of firing. J. Exp. Biol. 211, 1764–1774 (2008).
Watanabe, T., Shimazaki, T. & Oda, Y. Coordinated expression of two types of low-threshold K(+) channels establishes unique single spiking of Mauthner cells among segmentally homologous neurons in the zebrafish hindbrain. eNeuro 4, https://doi.org/10.1523/ENEURO.0249-17.2017 (2017).
Kalluri, R., Xue, J. & Eatock, R. A. Ion channels set spike timing regularity of mammalian vestibular afferent neurons. J. Neurophysiol. 104, 2034–2051 (2010).
Heidenreich, M. et al. KCNQ4 K+ channels tune mechanoreceptors for normal touch sensation in mouse and man. Nat. Neurosci. 15, 138–145 (2012).
Lau, D. et al. Impaired fast-spiking, suppressed cortical inhibition, and increased susceptibility to seizures in mice lacking Kv3.2K+ channel proteins. J. Neurosci. 20, 9071–9085 (2000).
Johnston, J., Forsythe, I. D. & Kopp-Scheinpflug, C. Going native: voltage-gated potassium channels controlling neuronal excitability. J. Physiol. 588, 3187–3200 (2010).
Petitpre, C. et al. Neuronal heterogeneity and stereotyped connectivity in the auditory afferent system. Nat. Commun. 9, 3691 (2018).
Connor, J. A. & Stevens, C. F. Prediction of repetitive firing behaviour from voltage clamp data on an isolated neurone soma. J. Physiol. 213, 31–53 (1971).
Kanold, P. O. & Manis, P. B. Transient potassium currents regulate the discharge patterns of dorsal cochlear nucleus pyramidal cells. J. Neurosci. 19, 2195–2208 (1999).
Shibata, R. et al. A-Type K+ current mediated by the Kv4 channel regulates the generation of action potential in developing cerebellar granule cells. J. Neurosci. 20, 4145–4155 (2000).
Rhode, W. S., Oertel, D. & Smith, P. H. Physiological response properties of cells labeled intracellularly with horseradish peroxidase in cat ventral cochlear nucleus. J. Comp. Neurol. 213, 448–463 (1983).
Trussell, L. O. Synaptic mechanisms for coding timing in auditory neurons. Annu. Rev. Physiol. 61, 477–496 (1999).
Manis, P. et al. The Endbulbs of Held. In: Synaptic Mechanisms in the Auditory System. (Eds. Trussell, L. O., Popper, A. N., & Fay, R. R.) 61–93 (Springer, 2011).
Wang, Y.-X., Wenthold, R. J., Ottersen, O. P. & Petralia, R. S. Endbulb synapses in the anteroventral cochlear nucleus express a specific subset of AMPA-type glutamate receptor subunits. J. Neurosci. 18, 1148–1160 (1998).
Petralia, R. S., Rubio, M. E., Wang, Y. X. & Wenthold, R. J. Differential distribution of glutamate receptors in the cochlear nuclei. Hear Res 147, 59–69 (2000).
Rubio, M. E. et al. The number and distribution of AMPA receptor channels containing fast kinetic GluA3 and GluA4 subunits at auditory nerve synapses depend on the target cells. Brain Struct. Funct. 222, 3375–3393 (2017).
Petralia, R. S., Wang, Y. X., Zhao, H. M. & Wenthold, R. J. Ionotropic and metabotropic glutamate receptors show unique postsynaptic, presynaptic, and glial localizations in the dorsal cochlear nucleus. J. Comp. Neurol. 372, 356–383 (1996).
Zhao, Y., Chen, S., Swensen, A. C., Qian, W. J. & Gouaux, E. Architecture and subunit arrangement of native AMPA receptors elucidated by cryo-EM. Science 364, 355–362 (2019).
Gardner, S. M., Trussell, L. O. & Oertel, D. Correlation of AMPA receptor subunit composition with synaptic input in the mammalian cochlear nuclei. J. Neurosci.: Off. J. Soc. Neurosci. 21, 7428–7437 (2001).
Gardner, S. M., Trussell, L. O. & Oertel, D. Time course and permeation of synaptic AMPA receptors in cochlear nuclear neurons correlate with input. J. Neurosci.: Off. J. Soc. Neurosci. 19, 8721–8729 (1999).
Futai, K., Okada, M., Matsuyama, K. & Takahashi, T. High-fidelity transmission acquired via a developmental decrease in NMDA receptor expression at an auditory synapse. J. Neurosci.: Off. J. Soc. Neurosci. 21, 3342–3349 (2001).
Bellingham, M. C., Lim, R. & Walmsley, B. Developmental changes in EPSC quantal size and quantal content at a central glutamatergic synapse in rat. J. Physiol. 511 3, 861–869 (1998).
Trussell, L. O. in The Oxford Handbook of the Auditory Brainstem (ed Karl Kandler) (Oxford University Press, 2019).
Goyer, D. et al. Slow cholinergic modulation of spike probability in ultra-fast time-coding sensory neurons. eNeuro 3, ENEURO.0186-16.2016 (2016).
Xie, R. & Manis, P. B. Target-specific IPSC kinetics promote temporal processing in auditory parallel pathways. J. Neurosci. 33, 1598–1614 (2013).
Lim, W., Mayer, B. & Pawson, T. Cell signaling (Garland Science, 2014).
Kopp-Scheinpflug, C. & Tempel, B. L. Decreased temporal precision of neuronal signaling as a candidate mechanism of auditory processing disorder. Hear. Res. 330, 213–220 (2015).
Griffiths, T. D. Central auditory processing disorders. Curr. Opin. Neurol. 15, 31–33 (2002).
Zeng, H. & Sanes, J. R. Neuronal cell-type classification: challenges, opportunities and the path forward. Nat. Rev. Neurosci. 18, 530–546 (2017).
Young, E. D. Parallel processing in the nervous system: evidence from sensory maps. Proc. Natl. Acad. Sci. USA 95, 933–934 (1998).
Clayton, K. K. et al. Auditory corticothalamic neurons are recruited by motor preparatory inputs. Curr. Biol. 31, 310–321 e315 (2021).
Daigle, T. L. et al. A suite of transgenic driver and reporter mouse lines with enhanced brain-cell-type targeting and functionality. Cell 174, 465–480.e422 (2018).
Wang, V. Y., Rose, M. F. & Zoghbi, H. Y. Math1 expression redefines the rhombic lip derivatives and reveals novel lineages within the brainstem and cerebellum. Neuron 48, 31–43 (2005).
de Agustín-Durán, D., Mateos-White, I., Fabra-Beser, J. & Gil-Sanz, C. Stick around: cell–cell adhesion molecules during neocortical development. Cells 10, 118 (2021).
Webber, A. & Raz, Y. Axon guidance cues in auditory development. Anat. Rec. Part A, Discov. Mol. Cell. Evolut. Biol. 288, 390–396 (2006).
Wong, K. et al. Auditory brainstem implants: recent progress and future perspectives. Front. Neurosci. 13, 10 (2019).
Martin, M. R. & Rickets, C. Histogenesis of the cochlear nucleus of the mouse. J. Comp. Neurol. 197, 169–184 (1981).
Schweitzer, L. & Cant, N. B. Differentiation of the giant and fusiform cells in the dorsal cochlear nucleus of the hamster. Brain Res. 352, 69–82 (1985).
Tolbert, L. P., Morest, D. K. & Yurgelun-Todd, D. A. The neuronal architecture of the anteroventral cochlear nucleus of the cat in the region of the cochlear nerve root: horseradish peroxidase labelling of identified cell types. Neuroscience 7, 3031–3052 (1982).
Liberman, M. C. Central projections of auditory-nerve fibers of differing spontaneous rate. I. Anteroventral cochlear nucleus. J. Comp. Neurol. 313, 240–258 (1991).
Brownell, W. E. Organization of the cat trapezoid body and the discharge characteristics of its fibers. Brain Res 94, 413–433 (1975).
Roos, M. J. & May, B. J. Classification of unit types in the anteroventral cochlear nucleus of laboratory mice. Hear. Res. 289, 13–26 (2012).
Trussell, L. O. & Oertel, D. in The Mammalian Auditory Pathways: Synaptic Organization and Microcircuits (eds Douglas L. Oliver, Nell B. Cant, Richard R. Fay, & Arthur N. Popper) 73–99 (Springer International Publishing, 2018).
Ruan, H. B., Zhang, N. & Gao, X. Identification of a novel point mutation of mouse proto-oncogene c-kit through N-ethyl-N-nitrosourea mutagenesis. Genetics 169, 819–831 (2005).
Spritz, R. A. & Beighton, P. Piebaldism with deafness: molecular evidence for an expanded syndrome. Am. J. Med. Genet. 75, 101–103 (1998).
Morris, S. A. & Daley, G. Q. A blueprint for engineering cell fate: current technologies to reprogram cell identity. Cell Res. 23, 33–48 (2013).
Graf, T. & Enver, T. Forcing cells to change lineages. Nature 462, 587–594 (2009).
Fujiyama, T. et al. Inhibitory and excitatory subtypes of cochlear nucleus neurons are defined by distinct bHLH transcription factors, Ptf1a and Atoh1. Development 136, 2049–2058 (2009).
Chessum, L. et al. Helios is a key transcriptional regulator of outer hair cell maturation. Nature 563, 696–700 (2018).
Sun, S. et al. Dual expression of Atoh1 and Ikzf2 promotes transformation of adult cochlear supporting cells into outer hair cells. Elife 10, https://doi.org/10.7554/eLife.66547 (2021).
Park, D., Bae, S., Yoon, T. H. & Ko, J. Molecular mechanisms of synaptic specificity: spotlight on hippocampal and cerebellar synapse organizers. Mol. Cells 41, 373–380 (2018).
Howell, D. M. et al. Molecular guidance cues necessary for axon pathfinding from the ventral cochlear nucleus. J. Comp. Neurol. 504, 533–549 (2007).
Kolson, D. R. et al. Temporal patterns of gene expression during calyx of held development. Dev. Neurobiol. 76, 166–189 (2016).
Kim, Y. J. et al. Dcc mediates functional assembly of peripheral auditory circuits. Sci. Rep. 6, 23799 (2016).
Nakamura, P. A., Hsieh, C. Y. & Cramer, K. S. EphB signaling regulates target innervation in the developing and deafferented auditory brainstem. Dev. Neurobiol. 72, 1243–1255 (2012).
Zhang, W. et al. Netrin-G2 and netrin-G2 ligand are both required for normal auditory responsiveness. Genes, Brain Behav. 7, 385–392 (2008).
Toyoshima, M. et al. Deficiency of neural recognition molecule NB-2 affects the development of glutamatergic auditory pathways from the ventral cochlear nucleus to the superior olivary complex in mouse. Dev. Biol. 336, 192–200 (2009).
Li, H. et al. Aberrant responses to acoustic stimuli in mice deficient for neural recognition molecule NB-2. Eur. J. Neurosci. 17, 929–936 (2003).
Shimoda, Y. & Watanabe, K. Contactins: emerging key roles in the development and function of the nervous system. Cell Adhes. Migr. 3, 64–70 (2009).
Huang, S. et al. A non-coding variant in 5’ untranslated region drove up-regulation of pseudo-kinase EPHA10 and caused non-syndromic hearing loss in humans. Hum. Mol. Genet. 32, 720–731 (2022).
Mohseni, M. et al. Exome sequencing utility in defining the genetic landscape of hearing loss and novel-gene discovery in Iran. Clin. Genet. 100, 59–78 (2021).
Mercati, O. et al. CNTN6 mutations are risk factors for abnormal auditory sensory perception in autism spectrum disorders. Mol. Psychiatry 22, 625–633 (2017).
Choucair, N. et al. Evidence that homozygous PTPRD gene microdeletion causes trigonocephaly, hearing loss, and intellectual disability. Mol. Cytogenet. 8, 39 (2015).
Völker, L. A. et al. Neph2/Kirrel3 regulates sensory input, motor coordination, and home-cage activity in rodents. Genes, Brain Behav. 17, e12516 (2018).
Bouilly, J. et al. DCC/NTN1 complex mutations in patients with congenital hypogonadotropic hypogonadism impair GnRH neuron development. Hum. Mol. Genet. 27, 359–372 (2018).
Bates, T. C. et al. Genetic variance in a component of the language acquisition device: ROBO1 polymorphisms associated with phonological buffer deficits. Behav. Genet. 41, 50–57 (2011).
Wells, H. R. R., Newman, T. A. & Williams, F. M. K. Genetics of age-related hearing loss. J. Neurosci. Res. 98, 1698–1704 (2020).
Alsaber, R., Tabone, C. J. & Kandpal, R. P. Predicting candidate genes for human deafness disorders: a bioinformatics approach. BMC Genom. 7, 180 (2006).
Pfister, M. H. et al. Clinical evidence for dystrophin dysfunction as a cause of hearing loss in locus DFN4. Laryngoscope 109, 730–735 (1999).
Kalay, E. et al. A novel locus for autosomal recessive nonsyndromic hearing impairment, DFNB63, maps to chromosome 11q13.2–q13.4. J. Mol. Med. 85, 397–404, (2007).
Pagnamenta, A. T. et al. Characterization of a family with rare deletions in CNTNAP5 and DOCK4 suggests novel risk loci for autism and dyslexia. Biol. psychiatry 68, 320–328 (2010).
Hannula-Jouppi, K. et al. The axon guidance receptor gene ROBO1 is a candidate gene for developmental dyslexia. PLoS Genet. 1, e50 (2005).
Leão, R. M. The ion channels and synapses responsible for the physiological diversity of mammalian lower brainstem auditory neurons. Hear. Res. 376, 33–46 (2019).
Karcz, A., Allen, P. D., Walton, J., Ison, J. R. & Kopp-Scheinpflug, C. Auditory deficits of Kcna1 deletion are similar to those of a monaural hearing impairment. Hear. Res 321, 45–51 (2015).
Tomlinson, S. E. et al. Clinical, genetic, neurophysiological and functional study of new mutations in episodic ataxia type 1. J. Neurol. Neurosurg. Psychiatry 84, 1107–1112 (2013).
Ison, J. R., Allen, P. D. & Oertel, D. Deleting the HCN1 subunit of hyperpolarization-activated ion channels in mice impairs acoustic startle reflexes, gap detection, and spatial localization. J. Assoc. Res. Otolaryngol. 18, 427–440 (2017).
Bleakley, L. E. et al. Cation leak underlies neuronal excitability in an HCN1 developmental and epileptic encephalopathy. Brain 144, 2060–2073 (2021).
Kharkovets, T. et al. KCNQ4, a K+ channel mutated in the form of dominant deafness, is expressed in the inner ear and the central auditory pathway. Proc. Natl. Acad. Sci. USA 97, 4333–4338 (2000).
Beisel, K. W. et al. Differential expression of KCNQ4 in inner hair cells and sensory neurons is the basis of progressive high-frequency hearing loss. J. Neurosci. 25, 9285–9293 (2005).
Middlebrooks, J. C. et al. Mutation in the kv3.3 voltage-gated potassium channel causing spinocerebellar ataxia 13 disrupts sound-localization mechanisms. PLoS One 8, e76749 (2013).
Yang, Y., Ramamurthy, B., Neef, A. & Xu-Friedman, M. A. Low somatic sodium conductance enhances action potential precision in time-coding auditory neurons. J. Neurosci. 36, 11999–12009 (2016).
Yu, F. H. & Catterall, W. A. Overview of the voltage-gated sodium channel family. Genome Biol. 4, 207 (2003).
Krishnaswami, S. R. et al. Using single nuclei for RNA-seq to capture the transcriptome of postmortem neurons. Nat. Protoc. 11, 499–524 (2016).
Yang, H. & Xu-Friedman, M. A. Relative roles of different mechanisms of depression at the mouse endbulb of Held. J. Neurophysiol. 99, 2510–2521 (2008).
Ngodup, T. et al. Activity-dependent, homeostatic regulation of neurotransmitter release from auditory nerve fibers. Proc. Natl. Acad. Sci. USA 112, 6479–6484 (2015).
Jiang, X. et al. Principles of connectivity among morphologically defined cell types in adult neocortex. Science 350, aac9462 (2015).
Cadwell, C. R. et al. Multimodal profiling of single-cell morphology, electrophysiology, and gene expression using Patch-seq. Nat. Protoc. 12, 2531–2553 (2017).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Liao, Y., Smyth, G. K. & Shi, W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930 (2014).
Yaylaoglu, M. B. et al. Comprehensive expression atlas of fibroblast growth factors and their receptors generated by a novel robotic in situ hybridization platform. Dev. Dyn. 234, 371–386 (2005).
Wolf, F. A., Angerer, P. & Theis, F. J. SCANPY: large-scale single-cell gene expression data analysis. Genome Biol. 19, 15 (2018).
Luecken, M. D. & Theis, F. J. Current best practices in single-cell RNA-seq analysis: a tutorial. Mol. Syst. Biol. 15, e8746 (2019).
Wolock, S. L., Lopez, R. & Klein, A. M. Scrublet: computational identification of cell doublets in single-cell transcriptomic data. Cell Syst. 8, 281–291.e289 (2019).
Korsunsky, I. et al. Fast, sensitive and accurate integration of single-cell data with Harmony. Nat. Methods 16, 1289–1296 (2019).
Polański, K. et al. BBKNN: fast batch alignment of single-cell transcriptomes. Bioinformatics 36, 964–965 (2019).
Kobak, D. & Berens, P. The art of using t-SNE for single-cell transcriptomics. Nat. Commun. 10, 5416 (2019).
Ming Hu, X. J. PatchView: a Python package for Patch-clamp data analysis and visualization. J. Open Source Softw. 7, 4706 (2022).
Henze, D. A. & Buzsaki, G. Action potential threshold of hippocampal pyramidal cells in vivo is increased by recent spiking activity. Neuroscience 105, 121–130 (2001).
Claudi, F. et al. Visualizing anatomically registered data with brainrender. eLife 10, e65751 (2021).
Crow, M., Paul, A., Ballouz, S., Huang, Z. J. & Gillis, J. Characterizing the replicability of cell types defined by single-cell RNA-sequencing data using MetaNeighbor. Nat. Commun. 9, 884 (2018).
Li, H. et al. Fly Cell Atlas: a single-nucleus transcriptomic atlas of the adult fruit fly. Science 375, eabk2432 (2022).
Lein, E. S. et al. Genome-wide atlas of gene expression in the adult mouse brain. Nature 445, 168–176 (2007).
Acknowledgements
We thank the members of Jiang and Trussell labs for technical assistance, discussions, and comments on the manuscript. Research reported in this publication is supported by research grants R01 MH122169, R01 MH120404, R01 NS110767 (to X.J.), R01 DC004450, R35NS116798 (to L.O.T.), and R01 DC017797 (to M.J.M.) from the National Institutes of Health. Research reported in this publication was also supported by the Main Street America Fund, a Shared Instrumentation grant from the NIH (1S10OD016167), by the Eunice Kennedy Shriver National Institute of Child Health & Human Development of the National Institutes of Health under Award Number P50HD103555 for use of the Microscopy Core facilities and the RNA In Situ Hybridization Core facility at Baylor College of Medicine, and by the NEI Core Grant for Vision Research (EY-002520-37) for use of the Bioengineering Core. We thank Drs. Lisa Goodrich and Arpiar Saunders for the comments on the manuscript and Drs. Dmitry Kobak, Philipp Berens, Yves Bernaerts, and Satoshi Okawa for helping with data analysis. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Author information
Authors and Affiliations
Contributions
J.J., T.N., and X.J. performed the Patch-seq experiments, J.J., Q.M., and S.L. performed the snRNA-seq. M.C.L. and J.J. performed the FISH staining. J.J. performed the immunostaining and confocal imaging. J.J., M.H., X.J., and T.N. analyzed the data. J.J. and X.J. prepared figures. X.J., L.T., and M.M. supervised all experiments and data analyses. X.J. and L.T. wrote the manuscript with input from all co-authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Jing, J., Hu, M., Ngodup, T. et al. Molecular logic for cellular specializations that initiate the auditory parallel processing pathways. Nat Commun 16, 489 (2025). https://doi.org/10.1038/s41467-024-55257-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-024-55257-z
