
Abstract
Observations show an increase in the seasonal cycle amplitude of CO2 in northern latitudes over the past half century. Although multiple drivers contribute, observations and inversion models cannot quantitatively account for the factors contributing to the increased CO2 amplitude and older versions of Earth System Models (ESMs) do not simulate it. Here we show that several current generation ESMs are closer to the observed CO2 amplitude and highlight that in the Community Earth System Model (CESM) agricultural nitrogen (N) fertilization increases CO2 amplitude by 1-3 ppm throughout the Northern Hemisphere and up to 9 ppm in agricultural hotspots. While agricultural N fertilization is the largest contributor to the enhanced amplitude (45%) in Northern Hemisphere land-atmosphere carbon fluxes in CESM, higher CO2 concentrations and warmer temperatures also contribute, though to a lesser extent (40% and 18% respectively). Our results emphasize the fundamental role of agricultural management in Northern Hemisphere carbon cycle feedbacks and illustrate that agricultural N fertilization should be considered in future carbon cycle simulations.
Similar content being viewed by others

Introduction
Increased seasonal amplitude of atmospheric CO2 measured in remote locations signal that anthropogenic activities have changed terrestrial carbon exchange with the atmosphere1. Observations illustrate that the seasonal amplitude of CO2 increased in the Northern Hemisphere, with an increase of 15% over the past 50 years at Mauna Loa, Hawaii, and more than 30% at Point Barrow, Alaska, where the seasonal cycles are much stronger2,3. These enhancements have largely been attributed to changes in high latitude ecosystems associated with climate change2,4,5,6 and agriculture7,8.
Several mechanisms may be responsible for observed changes in the amplitude of seasonal CO2 fluxes. For example, warmer summers and longer growing seasons across ecosystems in the high latitudes may accelerate rates of photosynthesis and accentuate draw-down of atmospheric CO2 in the boreal summer2,4,5,6,9,10,11. Forest regrowth in mid-latitudes and forest expansion across high latitudes may similarly increase photosynthetic drawdown of growing season CO22, as does the CO2 fertilization of photosynthesis4,5. Although higher respiratory CO2 release during the fall may also increase CO2 amplitude, increased photosynthesis is the dominant driver of the increased seasonal CO2 amplitude2,6. All these mechanisms – longer growing seasons, forest expansion, and CO2 fertilization of photosynthesis – emphasize increases in plant productivity in high latitude ecosystems, a greening trend that has been observed from space12. However, warmer temperatures and higher CO2 concentrations cannot fully explain the increased CO2 seasonal amplitude2,7,8.
The large increase in agricultural productivity associated with intensified management has also contributed to the increased Northern Hemisphere seasonal CO2 amplitude7,8 and may help to explain the missing contribution to increased CO2 seasonal amplitude. Agricultural practices have intensified over the past half century, initiated by the Green Revolution, increasing overall yield per area primarily through hybridization, irrigation, and fertilization13. Atmospheric circulation transports CO2 from temperate regions into high latitudes14, so agricultural intensification in the mid-latitudes may contribute to the large increases in seasonal CO2 amplitude observed at high latitude sites. While the combination of agricultural management and climate may explain the enhanced seasonal amplitude of the atmospheric CO2, research has yet to explicitly quantify the relative contributions of each. Existing efforts use a synthesis of literature and expert opinion15, which is limited by the lack of explicit agricultural management representation in models and therefore quantitative attribution. Additionally, no previous work has disentangled specific contributions of individual agricultural management practices like irrigation and fertilization. Here we investigate changes in the seasonal amplitude of atmospheric CO2 exchange that are simulated by ESMs and investigate the potential role of explicit agricultural management and climate change in driving changes in land-atmosphere CO2 exchange over the observational record. We illustrate that nitrogen fertilization contributes more to the increased CO2 amplitude than irrigation or climate change.
Results
Changes in CO2 amplitude simulated by Earth system models
The Coupled Model Intercomparison Project version 6 (CMIP6) ensemble of models brackets the amplitude of the seasonal cycle of atmospheric CO2 observations at Point Barrow Alaska, whereas nearly all the CMIP5 ensemble of models underestimate the amplitude (Fig. 1). Earth system models are often used to attribute the underlying mechanisms contributing to the observed carbon cycle changes because large-scale fluxes of carbon between the atmosphere and biosphere are not directly observable and inversion models cannot discern process contributions. However, the capability to attribute changes in the magnitude and trend of CO2 seasonality has been limited because older versions of ESMs used in CMIP5 underestimate the observed amplitude and the change in amplitude over the observational record at Point Barrow (Fig. 1a) and throughout the Northern Hemisphere high latitudes3. A variety of reasons for model underestimation of CO2 amplitude and trends have been proposed, including poor representation of high latitude vegetation and soil processes, inaccurate responses to CO2 fertilization, and an oversimplification in representing key terrestrial processes like nutrient limitation11; these biases have been used as the basis of an emergent constraint on the response of northern ecosystems to elevated CO211,16. However, the robustness of emergent constraints may be affected by the absence of atmospheric transport or representation of other important terrestrial processes in most models, like changes in crop productivity which exert a strong influence on the CO2 seasonality7,8. Since the seasonal CO2 amplitude is an integrated signal of change in terrestrial ecosystem processes, improving process representation to more accurately capture ecosystem dynamics may increase confidence in future carbon cycle projections, including terrestrial carbon sequestration and carbon-climate feedbacks. Newer iterations of models used for CMIP6 updated carbon cycle processes and better capture the amplitude in the seasonal cycle of CO2 at Point Barrow (Fig. 1b). Given these broad improvements in the CMIP6 models, we take a deeper look at atmospheric CO2 dynamics that are simulated by successive generations of the Community Earth System Model (CESM).

The CMIP5 (a) and CMIP6 (b) values plotted for each model are the change in the mean CO2 amplitude over the last ten years of the CMIP5 historical simulations (1995–2005) versus the change in amplitude at the end of the simulation relative to the 1970–1980 mean. Observations are averaged for the same timeframe as model simulations. X-axis error bars represent the standard deviation in the CO2 amplitude across years.
The magnitude of the seasonal CO2 cycle has improved in successive generations of the CESM (Fig. 2). The CESM117 was one of two CMIP5 models that included nitrogen limitation on plant growth. However, like most models participating in the CMIP5 experiment, CESM1 underestimated the mean CO2 amplitude for 1995–2005 compared to observations3 at Kumukahi, Mauna Loa, Niwot Ridge, and Point Barrow (Fig. 2a–d), and underestimated trends in CO2 amplitude compared to Point Barrow (Fig. 2h). The magnitude of CO2 amplitude in CESM version 218 is more comparable to observations than CESM1 (Figs. 2; also see Fig. S1 and Table S1), although CESM2 does not clearly improve CO2 amplitude trends over time. Both model versions underestimate the CO2 amplitude and the change in amplitude through time at high elevation sites Mauna Loa and Niwot Ridge (Fig. 2b, c, f, g), suggesting some inaccuracies in vertical atmospheric transport14,19. For example, CESM2 underestimates observed amplitude at higher altitude sites compared to nearby sites at sea level (e.g., comparing Kumukahi, Fig. 2a, and Mauna Loa, Fig. 2b). Analysis verifies that the seasonality of atmospheric CO2 increases throughout the free troposphere in CESM2, although the seasonality is weaker compared to observations as altitude increases (Fig. S2) highlighting the need for improvements to the representation of vertical CO2 transport. There have been numerous developments between CESM1 and CESM2 that have improved carbon cycle fluxes (e.g., photosynthetic capacity, soil decomposition, plant nitrogen use) and may contribute to differences in CO2 amplitude18,20, with the representation of agriculture21 one of the key contributions to changes in CO2 amplitude (Fig. S3).

Plots illustrate the annual cycle (a–d), averaged for 1995-2005 and trends (e–h) at Cape Kumukahi, HI (19.7oN,155.0oW; altitude = 0.3 m), Mauna Loa, HI (19.5oN, 155.6oW, altitude = 3397.0 m), Niwot Ridge, CO (40.1oN, 105.6oW, altitude = 3523.0 m), and Point Barrow, AK (71.3oN, 156.61oW, altitude = 11.0 m) from NOAA’s CO2 flask network (black), CESM1 (orange), and CESM2 (purple) simulations. Shaded area in a-d illustrates the standard deviation across years. Simulation data are from the model grid cell and atmospheric level nearest the ___location of observation.
Agricultural N fertilization and the amplitude of atmospheric CO2
The role of human activities in emissions-driven ESM simulations is typically represented through external model inputs of fossil fuel emissions and land use change, and few ESMs represent land management processes associated with agriculture22. Agriculture is an important contributor to seasonal carbon fluxes in the extratropics7,8,23, which underscores the potential importance of representing agriculture in ESMs24. However, in the CMIP5 ensemble few ESMs included a representation of agricultural management, and none included agricultural N fertilization (only two represent N limitation). In contrast, five of the fourteen CMIP6 models contributing ‘esm-hist’ simulations reported cropland N fertilizer fluxes (UKESM-1.0, CESM2, MIROC-ES2L, EC-Earth3-CC, NorESM).
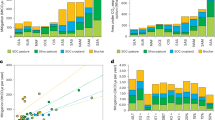
For agricultural systems, which are designed to maximize plant productivity, the addition of industrial N fertilizer is a fundamental aspect of modern crop management and has sustained human population growth24,25,26,27,28. It is arguably the most important contribution to increased crop yields over the past century21,29. Nitrogen is a key nutrient required for plant growth, and its availability limits terrestrial carbon gain through primary production30,31. Using the new representation of agricultural management20,21 in CESM2, we disentangle the contributions of increasing industrial N fertilizer use by comparing simulations with and without industrial N fertilizer in agricultural regions and illustrate that fertilizer contributes ~3 ppm of the Northern Hemisphere seasonal amplitude of CO2 (Figs. 3a, Fig. S3) and up to 9 ppm in key agricultural regions for 1996–2015 (Fig. 3a). Considering the large impact of agricultural N fertilization on photosynthesis and growth21,23, its large, local effect on seasonal carbon fluxes is perhaps not surprising. However, the large contribution of industrial N fertilization to the seasonal CO2 amplitude has not been previously identified due to difficulty in disentangling the signal in observational data and inversion models, as well as the limited representation in process models.

Panel a illustrates the change in mean amplitude of surface CO2 in CESM2 relative to a simulation without agricultural N fertilization for 1996–2015 (a). Panels b-e compare control CLM5 simulations (fully transient forcings and management) to simulations with individual forcings turned off to illustrate the change in land-only CLM5 simulation amplitude of net biome production (NBP), the net flux of carbon from terrestrial ecosystems to the atmosphere for 1996–2015 due to agricultural N fertilization (b), CO2 fertilization (c), irrigation (d), and climate change (e). The CO2 fertilization experiment uses 1850 CO2 concentrations; and the climate change experiment cycles over 1900–1920 climate. All differences illustrated here are the mean of 1996-2015.
Other contributions to seasonal carbon flux amplitude
Agricultural N fertilization causes similar spatial patterns of change in net biome production (NBP) from land-only simulations (Fig. 3b) and CO2 amplitude from coupled simulations using CESM2 (Fig. 3a). Net biome production represents the net land-air CO2 flux that determines terrestrial contributions to atmospheric CO2 concentrations in the CESM2-esm-hist simulations. The seasonal cycle of NBP, therefore, can be used as a proxy for CO2 amplitude32. Net biome production, however, does not capture the effects of atmospheric transport, as evident in the large changes in CO2 amplitude north of agricultural regions in East Asia from the CESM2-esm-hist simulations (Fig. 3a) and smaller changes in non-agricultural regions throughout the Northern Hemisphere, compared to the local distribution of NBP fluxes in Fig. 3b. Additionally, while ocean CO2 fluxes are an important component of the global carbon cycle, they are only a small contribution to the seasonal cycle of CO232. Given the dominant role of NBP in driving patterns of change in the CO2 annual cycle, we leverage data available from the Land Use Model Intercomparison Project (LUMIP) to identify the relative contributions of forcings (CO2 and climate) and agricultural management (fertilization and irrigation) on terrestrial carbon fluxes to the atmosphere. Analysis focuses on the Community Land Model version 5 (CLM5, the terrestrial component of CESM2) since it is the only model to contribute simulations with and without fertilizer and irrigation.
Agriculture is a key component of the seasonal CO2 amplitude7,8, and in our simulations, industrial N fertilizer has the largest impact on the seasonal amplitude of terrestrial carbon fluxes (0.54 g C m−2 d−1 in 2010–2015, contributing to a 45 ± 2.5% enhancement from the century prior (1910–1915) compared to the enhancement over the same time in the control CLM5 simulation; Figs. 3b, 4). Until now, increasing atmospheric CO2 concentrations was considered the dominant contribution to the increased amplitude in high latitude carbon fluxes4,32. Our results highlight the importance of elevated CO2 in increasing the seasonal amplitude of carbon fluxes throughout the Northern Hemisphere (0.39 g C m−2 d−1 in 2010–2015, a 40 ± 2.0% increase; Figs. 3c, 4). However, the impact emerges later (Fig. 4b) and is smaller in magnitude (peaks ≤ 2 g C m−2 d−1) than agricultural N fertilization (peaks > 2 g C m−2 d−1) in most regions (Fig. 3b, c). While the carbon cycle response to CO2 fertilization is a large uncertainty in previous generations of ESMs33, the response of CLM5 to elevated CO2 is within the observed range of responses in Free-Air CO2 Enrichment (FACE) experiments34.

Plots illustrate the 1996-2015 average annual cycle (a) and historical trends in amplitude (b) of NBP from land-only CESM2 (e.g., CLM5) simulations, averaged north of 30oN. The CLM5 simulation (purple) includes transient atmospheric forcing and active crop management (irrigation and fertilization). Additional perturbation simulations show the impacts of removing agricultural N fertilization (blue), not irrigating (yellow), preindustrial climate (orange), and preindustrial CO2 (green). Shaded area in a illustrates the standard deviation across years.
Although climate change has a smaller influence on Northern Hemisphere seasonal amplitude of NBP fluxes (18 ± 1.4% by 2010-2015; Fig. 3c, d), it has a strong latitudinal gradient and may act to further increase the CO2 amplitude in high latitudes (Fig. 3e)7,32. The effect of climate change across much of the boreal region is similar in magnitude to the effect of CO2 fertilization, but with little or no changes in mid and low latitudes (Fig. 3e). Warmer temperatures are often associated with a longer growing season in high latitudes and can increase the CO2 amplitude through increased ecosystem productivity that draws down CO2, and CO2 releases due to increased respiration through higher decomposition rates6. Previous work indeed highlights that the increasing Northern Hemisphere seasonal carbon amplitude is primarily due to increased gross primary productivity (GPP) that sequesters carbon during the growing season, with smaller contributions from ecosystem respiration2,6. Additionally, disentangling climate drivers highlights that warmer temperatures contribute to the increased amplitude more than precipitation in most high latitude regions32. In low- and mid-latitudes, however, other studies4,35,36 found that warmer temperatures decreased the seasonal amplitude of carbon fluxes due to increased water stress.
While other agricultural management practices are critical for increased crop production and can influence the carbon cycle37, they contribute less to the seasonal amplitude of carbon fluxes than N fertilization. For example, although irrigation is an important aspect of agricultural management, we find that its effect on Northern Hemisphere seasonal carbon fluxes is small (3.7 ± 0.46%) due to the relatively small proportion of crop area that is irrigated (Figs. 3d, 4). Irrigation contributions to local carbon fluxes, for example in the Indo-Gangetic Plain, are similar in magnitude to fertilization (Fig. 3d), suggesting that future expansion of irrigation38 may have increasingly important impacts on carbon fluxes. Land conversion from forest to cropland is another important component of agricultural management. Indeed, land cover change significantly impacts the carbon cycle39,40 and accounts for 18% of the total anthropogenic carbon emissions over the last 50 years41. However, it does not strongly contribute to changes in the seasonal cycle of Northern Hemisphere carbon fluxes in CLM5 or other models4,32, but it may contribute to the trends in amplitude change. The design of the LUMIP simulations does not allow for disentangling interactions among the drivers of change, although these are likely to be small relative to the direct effects isolated in this analysis. For example, previous work illustrates that interaction between irrigation and fertilization contributes ~10% of their combined effect on crop yields in CLM21.
Discussion
The increased use of irrigation, fertilizers, and high-yield cultivars over the past fifty years has tripled crop production with important carbon cycle consequences8,42. Our work illustrates that industrial N fertilization has a large impact on seasonal CO2 amplitude that has not been previously isolated, accounting for a large fraction of the agricultural contribution to the amplitude7,8,24. While CO2 fertilization and climate are also important drivers of changes in seasonal carbon flux amplitude, especially in high latitudes2,4,32, industrial N fertilization has a larger impact throughout the Northern Hemisphere, contributing to contemporary mean amplitude increases as large as 9 ppm CO2 in some regions, and increasing land-atmosphere carbon fluxes by 45% across the entire Northern Hemisphere. The magnitude of change due to industrial N fertilization on land-atmosphere carbon fluxes in 2010–2015 (0.54 g C m−2 d−1) is similar to that of CO2 and climate combined (0.55 g C m−2 d−1).
Although the role of agriculture has historically been debated, ESMs that are typically used to disentangle the drivers of changing CO2 amplitude do not include detailed representations of agriculture or management and therefore suggest that agriculture has a marginal contribution. Other studies2,7,8 that include agriculture illustrate that intensified agricultural production contributes to 17–45% of the increase in CO2 amplitude, with lower estimates from studies that do not represent agricultural N fertilizer2 or identify impacts from four key crop types7 rather than all crops. Our results align with the high-end estimate8 that accounts for global agricultural intensification and illustrate that agricultural N fertilization is an important component of seasonal carbon cycle fluxes that is currently overlooked by most ESMs. Given the key role of agricultural N fertilization in regulating carbon fluxes, we strongly encourage that models used for future carbon cycle simulations include a representation of agricultural N fertilization.
Additionally, emergent constraints on photosynthesis11,16 typically assume that climate change is the primary driver of increased seasonal CO2 amplitude. Our findings underscore the need to reconsider this assumption, given the important role of agricultural fertilization on seasonal CO2 amplitude changes. There are, of course, other contributions to increased CO2 amplitude, like shifts towards younger forests in high latitudes43 or other agricultural management practices44 that are not incorporated in these scenarios and might further improve model fidelity with observations. Ultimately, this work highlights that agricultural N fertilization is a profound impact of human activities on seasonal CO2 amplitude.
Methods
Coupled model intercomparison project
Analysis of CMIP simulations uses esm-hist simulations, which are emissions-driven transient historical simulations, where CO2 is calculated and anthropogenic sources are prescribed45. Data from esm-hist simulations used in our analysis includes all available CO2 data (variable ‘co2’) from the monthly atmospheric data (Amon) contributed to the Coupled Model Intercomparison Project (CMIP), versions 5 and 6. Data are available through the Earth System Grid Federation (ESGF; https://esgf-node.llnl.gov/projects/esgf-llnl/).
Observations for model evaluation
Model simulations were evaluated against observational data of CO2 concentrations, which were obtained from the National Oceanic and Atmospheric Administration’s CO2 flask network46 (https://www.esrl.noaa.gov/gmd/ccgg/). Data from Point Barrow, Alaska (71.3oN, 156.61oW, altitude = 11.0 m); Niwot Ridge, Colorado (40.1oN, 105.6oW, altitude = 3523.0 m); Cape Kumukahi, Hawaii (19.7oN,155.0oW; altitude = 0.3 m); and Mauna Loa, Hawaii (19.5oN, 155.6oW, altitude = 3397.0 m) were extracted for use in this analysis and compared to the nearest model grid points and atmospheric layers. Data for 24 additional sites were used for extended model evaluation (see Fig. S1).
Simulations of NEE from CLM5 were evaluated at six agricultural flux tower sites (Fig. S4). Sites were selected by combing three flux networks (Ameriflux, FluxNet, and National Ecological Observatory Network (NEON) flux tower networks) to determine locations where observations were collected in agricultural ecosystems. Sites were further filtered to include only those that grew crops simulated by CLM (corn, wheat, soy, cotton, rice, sugarcane) and included more than three years of data with no major gaps, which limited analysis to sites growing corn. Meteorological forcing and NEE evaluation data from the towers were used from the NCAR-NEON system47, the Plumbing of Land Surface Models (PLUMBER) project48, or downloaded from Ameriflux (https://ameriflux.lbl.gov/sites/site-search/) and FluxNet (https://fluxnet.org/data/fluxnet2015-dataset/). The data from these networks have limited information on management, such as crop rotations, planting dates, and rates and timing of fertilizer applications. Thus, simulations at each site use a single crop type, prognostic planting dates based on growing degree days, and fertilizer rates from the dataset used in global simulations21. Since management has strong influence on crop growth and productivity, mismatches between simulations and observations are expected. Nevertheless, our simulations show overall much better agreement with NEE observations when N fertilizer is accounted for than otherwise (Fig. S4), supporting our hypothesis at the site scale.
Community Earth System Model
The Community Earth System Model is an open-source community model that couples atmosphere, ocean, sea ice, land, land ice, and river transport components of the Earth system to simulate biogeophysical and biogeochemical processes for past, present, and future climates18. The CESM2 includes numerous improvements to previous CESM versions, including low-latitude precipitation, shortwave cloud forcing, and global land carbon accumulation18 which is affected by improvements to the representation of agriculture, photosynthetic capacity, plant nitrogen use, and vertically-resolved soil decomposition. Of particular relevance for this work is the new capability of the Community Land Model version 5 (CLM5)20, the land component of CESM2, to explicitly simulate agriculture.
The agriculture module simulates global transient agricultural expansion as six major crop types (corn, soy, wheat, cotton, rice, sugarcane) and represents key land management practices (irrigation, fertilization), all of which can impact carbon, nitrogen, water, and energy fluxes between the land and atmosphere. Specifically, simulated crops use the same physiology as natural vegetation (e.g, photosynthesis, stomatal conductance, etc.) but with crop-specific parameters and crop-specific temperature-triggered phenological phases (planting, leaf emergence, grain fill, and harvest), as described in Lombardozzi et al21. Industrial fertilizer is prescribed by crop type, year, and country based on fertilization rates from the observationally derived Land Use Harmonization Version 2 (LUH2)49 and is applied to all existing and new crop area each year, while irrigation is applied to all irrigated crop fractions if soil moisture is less than a specified threshold. Compared to observations, CLM5 underestimates global crop yields over the past 20 years due to not representing processes related to agricultural intensification (e.g, increased planting density, improved cultivars, etc.), which may contribute to an underestimation of simulated NBP and CO2 amplitude. Additional details about the crop model can be found in Lombardozzi et al21.
Both CESM2 and CLM5 have been extensively evaluated against available observations, which notably illustrate that the cumulative land carbon sink in CLM5 and CESM2 simulations are similar to observationally based estimates18,50. Specifically, Lawrence et al20. use the International Land Benchmarking system (ILAMB)51 to evaluate CLM5 net ecosystem exchange (NEE) and GPP against FLUXNET201552 data and also evaluate GPP using FLUXCOM53 and WECANN54 datasets. Additionally, Lombardozzi et al.47 evaluated CLM5 NEE and GPP at 48 National Ecological Observatory Network sites. Evaluation of CLM5 at six agricultural flux tower sites is included in Fig. S4 and illustrates that CLM5 simulations of NEE with fertilizer are similar in magnitude to observations and reduces model error compared to not including fertilizer, although the model does not capture the interannual variability in NEE amplitude observed at the sites, likely due to limited site-level information on agricultural management (crop rotations, timing and rates of fertilizer applications, planting dates, etc.) and limitations in model representation of agricultural management. Global net ecosystem carbon balance is evaluated against the Global Carbon Project41 and Hoffman et al.55 datasets, and response to elevated CO2 and nitrogen additions is evaluated against meta-analyzes of FACE and nitrogen addition experiments34. Specific to the Arctic-boreal zone, Birch et al.56 found that the timing of leaf onset and offset in CLM5 is typically later than observed. Additionally, CLM5 overestimates GPP and ecosystem respiration in this region due to inaccuracies in the representation of phenology, allocation, and photosynthetic capacity parameterizations, potentially contributing to inaccuracies in the simulated magnitude of NBP and CO2 amplitude.
Simulations from CESM218 are a large focus of the analysis because this is the only model contributing simulations to CMIP6 and LUMIP projects that includes a comprehensive representation of agriculture, including spatiotemporally changing distributions (based on the Land Use Harmonization version 2, LUH2)20,49, as well as explicit crop types and phenological phases, and fertilization and irrigation management (described above). Within CESM, the transient land use datasets are produced using the CLM Land Use Data Tool (http://www.cgd.ucar.edu/iam/projects/thesis/thesis-clm-landuse-tool.html), which merges present-day land cover distribution with historical and future transitions described in LUH249, translating these into changing distributions of the plant categories and management information used in CESM20. All simulations used in this analysis were run from 1850 through 2014 at nominal 1o horizontal resolution.
The impact of individual forcings on terrestrial carbon fluxes were identified using additional CESM2 simulations. To understand the impact of agricultural management, two simulations were branched from the CESM2-esm-hist CMIP6 simulation in 1970 (data available at the UCAR/NCAR Climate Data Gateway doi:10.5065/q6br-jr78), when the impact of agriculture is apparent on carbon cycle processes21. The first of these turned off the explicit representation of agriculture so that all crop areas are represented as generic C3 crops, where crop phenology is simulated as C3 grasses and do not include irrigation or fertilization (referred to as ‘generic crop’; this is the CESM1 representation of agriculture). The second uses the explicit representation of agriculture but removes industrial N fertilization from all existing and new crop area (referred to as ‘no fertilization’). To ensure that changes in CO2 fluxes were minimally impacted by model drift, each simulation equilibrated carbon fluxes in 1970 by cycling over a single year of forcing for ten years. The CESM2 simulated these alternative representations of agriculture in a CO2 emissions-forced historical scenario following the esm-hist experimental protocol.
Land Use Model Intercomparison Project
We additionally leveraged land-only CLM5 simulations available through the CMIP6 Land Use Model Intercomparison Project57 (LUMIP; data available through ESGF: https://esgf-node.llnl.gov/projects/cmip6/) to understand the impacts of anthropogenic forcings (CO2 and climate), agricultural management (irrigation and fertilization), and land cover change on carbon fluxes. We used data from land-hist (fully transient historical, including transient CO2, N-deposition, aerosol deposition, etc.), and other simulations that were the same as land-hist except with a single factor held constant: land-cCO2 (constant CO2), land-cClim (constant climate), land-crop-noIrrig (irrigated area held at 1850 levels), and land-crop-noFert (fertilization rates and area held at 1850 levels/distribution; see Table 2 in Lawrence et al57. for detailed description of LUMIP and these simulations). The LUMIP simulations use the land component of ESMs run with GSWP3v1 climate forcing data (http://hydro.iis.u-tokyo.ac.pf/GSWP3), and we focus analysis on CLM5, the land model used within CESM2, because it is the only LUMIP model that submitted simulations with agricultural fertilization and irrigation. Because CO2 concentrations are not available from land-only simulations, we use NBP, the net carbon flux from the land surface (the carbon flux variable that is passed to the atmosphere in coupled simulations), as a proxy for the atmospheric CO2 concentration dynamics. We recognize that land-only simulations cannot account for the impacts of circulation and oceanic CO2 uptake on the distribution and seasonal cycling of CO2.
Amplitude and trend calculations
Seasonality in terrestrial carbon fluxes is due to many factors, including GPP, ecosystem respiration, and carbon losses from fires and land use change. Our analysis focuses on the seasonal amplitude of CO2 from coupled simulations (CMIP and CESM) and NBP fluxes from land-only simulations (CLM5). Net biome production is the terrestrial carbon flux from CLM5 that is passed to the atmosphere in coupled simulations. We quantify the seasonal amplitude of CO2 and NBP as the absolute difference between the annual maximum and minimum monthly averaged fluxes, representing the net source or sink of carbon fluxes to the atmosphere. Note that maximum NBP values indicate the peak net carbon sink. The linear trend in amplitude through time was removed from the calculations of the mean annual cycle. All amplitude analysis focuses on the Northern Hemisphere, > 30 oN, unless otherwise specified. The Northern Hemisphere has larger land area and more dramatic changes in the seasonal cycle of carbon fluxes.
The trends in amplitude for CESM1 and CESM2 were evaluated against trends in observational data (see description above) from the National Oceanic and Atmospheric Administration’s CO2 flask network46 (https://www.esrl.noaa.gov/gmd/ccgg/). The model data were available from 1970 through 2020 by combining historical (through 2005 for CESM1 and 2014 for CESM2) and future (RCP8.5 for CESM1 and SSP5-85 for CESM2) scenarios. Each station has a different record length within this time, so we truncate the model timeseries to have the same start and end year as the observational record for each station. Some stations have missing data for individual years within the middle of their time series, and our approach effectively assumes zero anomalies from climatology for these years. Regression slopes (b) and intercepts (a) were calculated using a least squares linear regression approach. Uncertainties on the slope (b) were calculated by resampling the residuals from the original regression slope. The original data were given by CO2(y) = a + by + e(y), where y is year and e represents the residuals from the original regression slope for each year. For a timeseries of length N, we randomly sample, with replacement, N values from the distribution of e(y), generate a new time series CO2’(y) = a + by + e’(y) where e’(y) are the randomly resampled residuals, and compute the least squared regression slope of this new time series. We repeat this 1000 times to generate a distribution of 1000 estimates of the regression slope, centered on the original, and estimate the uncertainty on the regression slope as the 2.5th and 97.5th percentile range of these bootstrapped slopes, encompassing 95% of the resampled slopes.
Data availability
Coupled Model Intercomparison Project (CMIP), versions 5 and 6 data are available through the Earth System Grid Federation (ESGF): https://esgf-node.llnl.gov/projects/esgf-llnl/. Land-only CLM5 simulations used here are available from the CMIP6 Land Use Model Intercomparison Project57 (LUMIP) through ESGF: https://esgf-node.llnl.gov/projects/cmip6/. Observational data of CO2 concentrations were obtained from the National Oceanic and Atmospheric Administration’s CO2 flask network46: https://www.esrl.noaa.gov/gmd/ccgg/. Data from the CESM2-esm-hist simulation without agricultural fertilization are available through the UCAR/NCAR Climate Data Gateway doi:10.5065/q6br-jr78.
Code availability
Code for the CESM2 model used in this work is available online at www.cesm.ucar.edu:/models/cesm2.
References
Bacastow, R. B., Keeling, C. D. & Whorf, T. P. Seasonal amplitude increase in atmospheric CO 2 concentration at Mauna Loa, Hawaii, 1959–1982. J. Geophys. Res. Atmospheres 90, 10529–10540 (1985).
Forkel, M. et al. Enhanced seasonal CO 2 exchange caused by amplified plant productivity in northern ecosystems. Science 351, 696–699 (2016).
Graven, H. D. et al. Enhanced seasonal exchange of CO2 by Northern ecosystems since 1960. Science 341, 1085–1089 (2013).
Bastos, A. et al. Contrasting effects of CO2 fertilization, land-use change and warming on seasonal amplitude of Northern Hemisphere CO2 exchange. Atmospheric Chem. Phys. 19, 12361–12375 (2019).
Piao, S. et al. Net carbon dioxide losses of northern ecosystems in response to autumn warming. Nature 451, 49–52 (2008).
Welp, L. R. et al. Increasing summer net CO2 uptake in high northern ecosystems inferredfrom atmospheric inversions and comparisons to remote-sensing NDVI. Atmospheric Chem. Phys. 16, 9047–9066 (2016).
Gray, J. M. et al. Direct human influence on atmospheric CO2 seasonality from increased cropland productivity. Nature 515, 398–401 (2014).
Zeng, N. et al. Agricultural green revolution as a driver of increasing atmospheric CO2 seasonal amplitude. Nature 515, 394–397 (2014).
Elmendorf, S. C. et al. Plot-scale evidence of tundra vegetation change and links to recent summer warming. Nat. Clim. Change 2, 453–457 (2012).
Liu, J., Wennberg, P. O., Parazoo, N. C., Yin, Y. & Frankenberg, C. Observational constraints on the response of high‐latitude northern forests to warming. AGU Adv 1, e2020AV000228 (2020).
Winkler, A. J., Myneni, R. B., Alexandrov, G. A. & Brovkin, V. Earth system models underestimate carbon fixation by plants in the high latitudes. Nat. Commun. 10, 885 (2019).
Ju, J. & Masek, J. G. The vegetation greenness trend in Canada and US Alaska from 1984–2012 Landsat data. Remote Sens. Environ. 176, 1–16 (2016).
Pingali, P. L. Green Revolution: Impacts, limits, and the path ahead. Proc. Natl. Acad. Sci. 109, 12302–12308 (2012).
Lin, X. et al. Siberian and temperate ecosystems shape Northern Hemisphere atmospheric CO2 seasonal amplification. Proc. Natl. Acad. Sci. 117, 21079–21087 (2020).
Liu, Z. et al. Seasonal CO2 amplitude in northern high latitudes. Nat. Rev. Earth Environ. 5, 802–817 (2024).
Wenzel, S., Cox, P. M., Eyring, V. & Friedlingstein, P. Projected land photosynthesis constrained by changes in the seasonal cycle of atmospheric CO2. Nature 538, 499–501 (2016).
Hurrell, J. W. et al. The community earth system model: a framework for collaborative research. BAMS 94, 1339–1360 (2013).
Danabasoglu, G. et al. The community earth system moDEL VERsion 2 (CESM2). J. Adv. Model. Earth Syst. 12, e2019MS001916 (2020).
Keppel-Aleks, G. et al. Atmospheric carbon dioxide variability in the community earth system model: evaluation and transient dynamics during the twentieth and twenty-first centuries. J. Clim. 26, 4447–4475 (2013).
Lawrence, D. M. et al. The community land model version 5: description of new features, benchmarking, and impact of forcing uncertainty. J. Adv. Model. Earth Syst. 11, 4245–4287 (2019).
Lombardozzi, D. L. et al. Simulating agriculture in the community land model version 5. J. Geophys. Res. Biogeosciences 125, e2019JG005529 (2020).
Pongratz, J. et al. Models meet data: challenges and opportunities in implementing land management in Earth system models. Glob. Change Biol. 24, 1470–1487 (2018).
Guanter, L. et al. Global and time-resolved monitoring of crop photosynthesis with chlorophyll fluorescence. Proc. Natl. Acad. Sci. 111, E1327–33 (2014).
Corbin, K. D. et al. Assessing the impact of crops on regional CO2 fluxes and atmospheric concentrations. Tellus B Chem. Phys. Meteorol. 62, 521 (2010).
Galloway, J. N. et al. Nitrogen cycles: past, present, and future. Biogeochemistry 70, 153–226 (2004).
Gruber, N. & Galloway, J. N. An Earth-system perspective of the global nitrogen cycle. Nature 451, 293–296 (2008).
Fowler, D. et al. The global nitrogen cycle in the twenty-first century. Philos. Trans. R. Soc. B Biol. Sci. 368, 20130164 (2013).
Robertson, G. P. & Vitousek, P. M. Nitrogen in agriculture: balancing the cost of an essential resource. Annu. Rev. Environ. Resour. 34, 97–125 (2009).
Stewart, W. M. & Roberts, T. L. Food security and the role of fertilizer in supporting it. Procedia Eng 46, 76–82 (2012).
LeBauer, D. S. & Treseder, K. K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 89, 371–379 (2008).
Thomas, R. Q., Canham, C. D., Weathers, K. C. & Goodale, C. L. Increased tree carbon storage in response to nitrogen deposition in the US. Nat. Geosci. 3, 13–17 (2009).
Piao, S. et al. On the causes of trends in the seasonal amplitude of atmospheric CO2. Glob. Change Biol. 24, 608–616 (2017).
Arora, V. K. et al. Carbon–concentration and carbon–climate feedbacks in CMIP5 earth system Models. J. Clim. 26, 5289–5314 (2013).
Wieder, W. R. et al. Beyond static benchmarking: using experimental manipulations to evaluate land model assumptions. Glob. Biogeochem. Cycles 33, 1289–1309 (2019).
Buermann, W. et al. Widespread seasonal compensation effects of spring warming on northern plant productivity. Nature 562, 110–114 (2018).
Wang, K. et al. Causes of slowing‐down seasonal CO2 amplitude at Mauna Loa. Glob. Change Biol. 26, 4462–4477 (2020).
Lal, R. Agricultural activities and the global carbon cycle. Nutr. Cycl. Agroecosystems 70, 103–116 (2004).
Rosa, L. et al. Potential for sustainable irrigation expansion in a 3 °C warmer climate. Proc. Natl. Acad. Sci. 117, 29526–29534 (2020).
Houghton, R. A. et al. Carbon emissions from land use and land-cover change. Biogeosciences 9, 5125–5142 (2012).
Lawrence, P. J., Lawrence, D. M. & Hurtt, G. C. Attributing the carbon cycle impacts of cmip5 historical and future land use and land cover change in the community earth system model (CESM1). J. Geophys. Res. Biogeosciences 123, 1732–1755 (2018).
Friedlingstein, P. et al. Global carbon budget 2019. Earth Syst. Sci. Data 11, 1783–1838 (2019).
MacBean, N. & Peylin, P. Agriculture and the global carbon cycle. Nature 515, 351–352 (2014).
Welp, L. R., Randerson, J. T. & Liu, H. P. Seasonal exchange of CO2 and δ18 O‐CO2 varies with postfire succession in boreal forest ecosystems. J. Geophys. Res. Biogeosciences 111, 2005JG000126 (2006).
Sanderman, J., Hengl, T. & Fiske, G. J. Soil carbon debt of 12,000 years of human land use. Proc. Natl. Acad. Sci. 114, 9575–9580 (2017).
Eyring, V. et al. Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) experimental design and organization. Geosci. Model Dev. 9, 1937–1958 (2016).
NOAA ESRL GML CCGG Group. Earth system research laboratory carbon cycle and greenhouse gases group flask-air sample measurements of CO2 at global and regional background sites, 1967-Present. https://doi.org/10.15138/WKGJ-F215 (2019).
Lombardozzi, D. L. et al. Overcoming barriers to enable convergence research by integrating ecological and climate sciences: the NCAR–NEON system Version 1. Geosci. Model Dev. 16, 5979–6000 (2023).
Best, M. J. et al. The plumbing of land surface models: benchmarking model performance. J. Hydrometeorol. 16, 1425–1442 (2015).
Hurtt, G. C. et al. Harmonization of global land-use change and management for the period 850–2100 (LUH2) for CMIP6. Geosci. Model Dev. 13, 5425–5464 (2020).
Bonan, G. B. et al. Model structure and climate data uncertainty in historical simulations of the terrestrial carbon cycle (1850–2014). Glob. Biogeochem. Cycles 33, 1310–1326 (2019).
Collier, N. et al. The international land model benchmarking (ILAMB) System: design, theory, and implementation. J. Adv. Model. Earth Syst. 10, 2731–2754 (2018).
Pastorello, G. et al. The FLUXNET2015 dataset and the ONEFlux processing pipeline for eddy covariance data. Sci. Data 7, 225 (2020).
Jung, M. et al. Scaling carbon fluxes from eddy covariance sites to globe: synthesis and evaluation of the FLUXCOM approach. Biogeosciences 17, 1343–1365 (2020).
Alemohammad, S. H. et al. Water, Energy, and Carbon with Artificial Neural Networks (WECANN): a statistically based estimate of global surface turbulent fluxes and gross primary productivity using solar-induced fluorescence. Biogeosciences 14, 4101–4124 (2017).
Hoffman, F. M. et al. Causes and implications of persistent atmospheric carbon dioxide biases in Earth system models. J. Geophys. Res. Biogeosciences 119, 141–162 (2014).
Birch, L. et al. Addressing biases in Arctic–boreal carbon cycling in the community land model version 5. Geosci. Model Dev. 14, 3361–3382 (2021).
Lawrence, D. M. et al. The Land Use Model Intercomparison Project (LUMIP) contribution to CMIP6: rationale and experimental design. Geosci. Model Dev. 9, 2973–2998 (2016).
Acknowledgements
The CESM project is supported primarily by the National Science Foundation (NSF). This material is based upon work supported by the NSF National Center for Atmospheric Research, which is a major facility sponsored by the NSF under Cooperative Agreement 1852977. Computing and data storage resources, including the Cheyenne (doi:10.5065/D6RX99HX) and Derecho (doi:10.5065/qx9a-pg09) supercomputers, were provided by the Computational and Information Systems Laboratory (CISL) at NCAR. We acknowledge the World Climate Research Program, which, through its Working Group on Coupled Modeling, coordinated and promoted CMIP5 and CMIP6. We thank the climate. Federation (ESGF) for archiving the data and providing access, and the multiple funding agencies who support CMIP5, CMIP6, and ESGF. We would like to thank Pieter Tans and Edward Dlugokencky for the use of NOAA Flask data to evaluate model simulations, and Katie Dagon, Deepak Cherian, Thomas Martin, and Anderson Banihirwe for help troubleshooting code and downloading CMIP6 data. We would also like to thank Heather Graven for providing processed observational data from HIPPO and IGY campaigns. DLL and WRW were supported by the U.S. Department of Agriculture NIFA Award 2015-67003-23485 and the National Science Foundation DISES Award 2409246. DLL was also supported by the U.S. Department of Agriculture NIFA Award 2021-0455.
Author information
Authors and Affiliations
Contributions
D.L.L. designed the research, carried out the analysis, and wrote the paper. I.R.S., J.L., Z.L., and Y.S. contributed analysis for model evaluation. All authors, including W.R.W., G.K.-A., J.L., Z.L., Y.S., I.R.S., D.M.L., G.B.B., X.L., C.D.K., P.F., and K.L. contributed to editing and revising the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review.
Peer review information
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Lombardozzi, D.L., Wieder, W.R., Keppel-Aleks, G. et al. Agricultural fertilization significantly enhances amplitude of land-atmosphere CO2 exchange. Nat Commun 16, 1742 (2025). https://doi.org/10.1038/s41467-025-56730-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-025-56730-z
