
Abstract
Targeting macrophage inhibitory receptors like signal regulatory protein α (SIRPα) is a promising avenue in cancer treatment. Whereas the ligand of SIRPα, CD47, is widely expressed on tumor cells, its simultaneous presence on all normal cells raises concerns about toxicity and efficacy. This study identifies CD200R1, which binds CD200 on specific tumor types and limited normal cells, as an alternative inhibitory checkpoint for phagocytosis. Blocking or removing CD200R1 from macrophages or CD200 from tumor cells increases phagocytosis and suppresses tumor growth. In humans, CD200R1 is mainly expressed in immunosuppressive macrophages and is induced by interleukin-4. Unlike SIRPα that utilizes phosphatases Src homology 2 ___domain phosphatase (SHP)−1 and SHP-2, CD200R1 mediates its inhibitory effect via the kinase Csk. Combined CD200R1-CD200 and SIRPα-CD47 blockade further boosts phagocytosis and reduces tumor growth of CD200-expressing tumors, compared to either blockade alone. Thus, targeting CD200R1-CD200 is a promising strategy for immune checkpoint blockade in macrophages, either alone or alongside blockade of other checkpoints.
Similar content being viewed by others

Introduction
Immunotherapy is one of the most significant advancements in anti-cancer treatment of recent years1,2. Most efforts have concentrated on blocking inhibitory immune checkpoints on T cells, such as PD-1 and CTLA-4. These strategies have shown impressive results against solid tumors like melanoma and renal cancer but have had limited success with hematological malignancies. However, promising results against hematological cancers have emerged from blocking inhibitory checkpoints on phagocytes, particularly macrophages3,4.
Notably, the blockade of SIRPα, an inhibitory receptor on macrophages that recruits the phosphatases SHP-1 and SHP-2, or of its ligand CD47 that is widely expressed on cancer cells, has been shown to enhance tumor cell phagocytosis and suppress tumor growth in mice3,4. Phase 1 and 2 clinical trials have also suggested the anti-tumor efficacy of SIRPα-CD47 blockade in hematological malignancies such as diffuse large B cell lymphoma, follicular lymphoma, Sezary’s syndrome and some solid tumors5,6,7,8,9.
Despite these advances, SIRPα-CD47 blockade has demonstrated limited efficacy against other tumors, including myelodysplastic syndrome and acute myelogenous leukemia, and toxicity issues such as anemia, thrombocytopenia and lymphopenia frequently arose due to CD47’s broad expression on normal cells7,10. Suggested approaches to address these issues are to block alternative macrophage inhibitory checkpoints or to combine SIRPα-CD47 blockade with blockade of other checkpoints to lower treatment dosage7.
Macrophage activation is regulated by a range of activating and inhibitory receptors3,4,11. When activating receptors, such as Fc receptors (FcRs), integrin CD11b and SLAMF7, are predominantly engaged by ligands on potential target cells, macrophages are activated and exhibit phagocytosis. In contrast, prevalent engagement of inhibitory receptors like SIRPα by ligands on potential target cells suppresses activation and phagocytosis. Most, if not all, macrophage subtypes have a phagocytic capacity, including the so-called inflammatory (M1) and anti-inflammatory (M2) macrophages, especially in the context of phagocytosis inhibitory checkpoint blockade12,13.
CD200R1 is a transmembrane receptor expressed on macrophages and other immune cells such as T cells, mast cells, and other myeloid cells14,15. It recognizes CD200, a ligand expressed in a restricted fashion on B cell malignancies, including chronic lymphocytic leukemia (CLL) and multiple myeloma (MM), and some solid tumors like melanoma and lung cancer16,17,18,19. In several tumor types, higher expression of CD200 correlated with a worse prognosis16,17,18,19. Engagement of CD200R1 by CD200 was reported to inhibit T cell activation, mast cell degranulation, and Toll-like receptor (TLR) signals in myeloid cells, seemingly through recruitment of Ras-GTPase-activating protein (Ras-GAP), which inhibits the Ras-Erk pathway20,21,22,23,24,25. Identification of CD200R1 as an inhibitory axis in T cells led to the testing of blocking CD200 mAb samalizumab and blocking CD200R1 mAb 23ME-00610 in phase 1 clinical trials of human cancer, with encouraging results including limited toxicity and some clinical responses25,26.
While the CD200R1-CD200 interaction has been reported to influence inflammatory responses by macrophages—possibly leading to indirect consequences on phagocytosis14,21,27,28—a direct impact on the phagocytosis process, in particular of cancer cells, has not been assessed. The molecular mechanism through which CD200R1 may suppress activating pro-phagocytic receptors also remains to be elucidated. Additionally, the therapeutic potential of targeting the CD200R1-CD200 pathway, alone or in combination with other modalities, including SIRPα-CD47 blockade, to enhance phagocytosis has yet to be investigated.
In this study, we discovered that the antibody-mediated blockade or loss of CD200R1 on macrophages or CD200 on tumor cells increased phagocytosis and enhanced macrophage-dependent anti-tumor responses in vivo. Acute blockade of the CD200R1-CD200 interaction resulted in a rapid (within 15 min) augmentation of actin polarization towards tumor cells, suggesting that the impact of CD200R1-CD200 on phagocytosis was direct. In humans, CD200R1 was induced by IL-4 and predominantly expressed in anti-inflammatory (M2-type) macrophages. Unlike SIRPα that acts through SHP-1 and SHP-2, CD200R1 mediated its inhibitory impact on phagocytosis by way of Csk, a protein tyrosine kinase that inactivates Src family kinases. When combined, blockade of CD200R1-CD200 and SIRPα-CD47 worked synergistically to enhance phagocytosis and inhibit tumor growth. Therefore, CD200R1 is a phagocytosis inhibitory checkpoint in macrophages that is triggered by CD200 expressed on tumor cells. Blocking this pathway may provide useful strategies for eliminating CD200-positive malignancies.
Results
CD200R1, a CD11b-interacting protein, inhibits phagocytosis
Several studies have demonstrated that inhibitory receptors, such as SIRPα and PD-1, mediate their inhibitory functions in part through physical association, direct or indirect, with the activating receptors that they suppress29,30,31. In macrophages, SIRPα, an immunoreceptor tyrosine-based inhibitory motif (ITIM)-containing receptor, closely associates with CD11b, the αM subunit of Mac-1, a key effector in phagocytosis30,31. CD11b serves not only as a complement receptor but also as a co-receptor for pro-phagocytic receptors such as FcRs and SLAMF73.
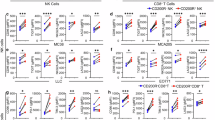
We aimed to identify CD11b-associated inhibitory receptors, beyond SIRPα, that control phagocytosis when engaged by ligands expressed on cancer cells and act through a molecular mechanism distinct from that of SIRPα. CD11b was immunoprecipitated from mouse bone marrow-derived macrophages (BMDMs), and the associated proteins were analyzed using mass spectrometry. A few inhibitory receptors were identified in CD11b immunoprecipitates from wild-type BMDMs, but not from CD18 (the β2 subunit of Mac-1)-deficient BMDMs, which lack CD11b (Fig. 1a and Supplementary Table 1). Some of these receptors, namely SIRPα, Ly-9, CD84 and CD200R1, recognize protein ligands potentially expressed at the surface of cancer cells. Most contain ITIMs or immunoreceptor tyrosine-based switch motifs (ITSMs), including SIRPα and Ly-9, both known to inhibit phagocytosis3,4,32. CD84 is related to Ly-9. The notable exception was CD200R1, which contains a NPXY (asparagine-proline-any residue-tyrosine) motif and has not been reported to suppress pro-phagocytic receptors14,15. The co-immunoprecipitation of CD200R1 and CD11b was validated by protein immunoblots (Fig. 1b).

a Mass spectrometry analysis of CD11b immunoprecipitates from BMDMs. Left: experimental workflow for isolating the CD11b-interacting proteins. Right: relative abundance of CD11b-interacting proteins, with a focus on cell surface receptors with inhibitory potential, indicated by a color gradient. ITIM, immunoreceptor tyrosine-based inhibitory motif; ITSM, immunoreceptor tyrosine-based switch motif; NPXY, asparagine, proline, any residue, tyrosine. IP, immunoprecipitation. b Immunoblot analysis of CD11b and CD200R1 interaction in immunoprecipitates from WT, Itgb2−/− and Cd200r1−/− BMDMs. Relative abundance is shown at the bottom of each panel. c Flow cytometry analysis of CD200 expression on parental (top) and Tac expression on Tac+ (bottom) mouse tumor cell lines. Red curves represent staining with CD200 or Tac mAbs. Filled curves, control (Ctrl) mAbs. d Microscopy-based phagocytosis assay of non-opsonized (−) or IgG-opsonized (+) Tac+ WEHI-231, Tac+ A20, Tac+ J558 and Tac+ TUBO cells by WT BMDMs, in presence of CD200 mAb OX-90 (rat IgG2a) or Ctrl mAb 2A3 (rat IgG2a). Tumor cells were opsonized with Tac mAb 7G7 (mouse IgG2a), (n = 3). e Time-course pHrodo-based phagocytosis assay using IgG-opsonized Tac+ WEHI-231 cells and WT BMDMs. WEHI-231 cells were labeled with pHrodo red dye, and BMDMs were labeled with CSFE. Phagocytosis over time (0-4 h) was analyzed using an IncuCyte Live Cell Analyzer. Left: representative images at 2 h (scale bar, 100 μm; arrows, BMDMs with engulfed tumor cells). Right: quantification of cumulative phagocytosis (top) and time-specific increase in phagocytosis (Δ phagocytosis; bottom) over 0-4 h, (n = 3). f Confocal microscope-based conjugate formation and actin polarization assay of IgG-opsonized WEHI-231 cells labeled with CSFE (green) and co-incubated with WT BMDMs labeled with Cell Trace Violet (CTV; blue), in presence of CD200 mAb or Ctrl mAb. Actin (red) was detected by β-actin mAb. Left: representative images (scale bar, 10 μm; arrows, BMDMs with fully polarized actin). Scale, 10 μm. Right: quantification of conjugate formation (top) and of conjugates with fully polarized actin (bottom), (n = 3). g As per Fig. 1d, except that phagocytosis of complement (C3bi)-opsonized WEHI-231 cells by WT BMDMs in the presence of blocking CD200 mAb OX-90, blocking CD11b mAb 5C6 (rat IgG2b), Ctrl mAb 2A3 or Ctrl mAb LTF-2 (rat IgG2b), (n = 3). h As per Fig. 1d, expect that phagocytosis of IgG-opsonized WEHI-231 cells by WT BMDMs in the presence of CD200R1 mAb OX-131 (mouse IgG1, Fc-silent) or Ctrl mAb (mouse IgG1, Fc-silent), (n = 3). Data are from three (a–h) independent experiments. Each symbol represents one mouse. Data are presented as mean ± s.e.m. Statistical analysis: two-tailed t-test (d–f, h) with multiple comparisons (e); One-way ANOVA test with multiple comparisons (g); ns, not significant. See also Supplementary Fig. 1 and Supplementary Table 1.
To test the involvement of CD200R1 in phagocytosis, the effect of antibody-mediated blockade of CD200R1-CD200 on phagocytosis of CD200+ mouse tumor cell lines, including B cell lines WEHI-231, A20 and J558, as well as breast cancer cell line TUBO, was studied (Fig. 1c and Supplementary Fig. 1a, b). Variants of these cell lines expressing the Tac molecule (human CD25) were also created, to enable opsonization by mouse IgG2a Tac mAb 7G728 (Fig. 1c and Supplementary Fig. 1b). CD200 mAb OX-90, but not a control (Ctrl) mAb, inhibited the binding of a CD200R1-Fc fusion protein to WEHI-231 cells, confirming its effectiveness as a blocking mAb (Supplementary Fig. 1c). In a 2-h-microscopy-based phagocytosis assay, OX-90 enhanced the ability of wild-type C57BL/6 mouse BMDMs to phagocytose IgG-opsonized CD200+ tumor cells, compared to Ctrl mAb (Fig. 1d). While WEHI-231, A20, and TUBO did not show increased phagocytosis in the absence of IgG opsonization, J558 did, as will be further discussed below (Fig. 1d).
Similar effects were observed in a pHrodo-based assay, over a wide range of co-incubation times as short as 1 h that were analyzed by Incucyte (1–4 h; Fig. 1e) or flow cytometry (2h; Supplementary Fig. 1d–f). Confocal microscopy assays revealed that blocking CD200 mAbs also augmented the proportion of macrophages displaying actin polarization around the tumor cells within 15 min of co-incubation (Fig. 1f). However, there was no increase in conjugate formation between the macrophages and the tumor cells.
The ability of CD200 blockade to enhance phagocytosis of IgG-opsonized WEHI-231 was eliminated in BMDMs lacking FcRγ (Fcer1g−/−), confirming that phagocytosis was mediated by FcRs30 (Supplementary Fig. 1g). Likewise, the increased phagocytosis of non-opsonized J558 cells was negated in BMDMs without SLAMF7 (Slamf7−/−), validating that this response was mediated by SLAMF728 (Supplementary Fig. 1h). CD200 blockade also improved the ability of WT BMDMs to phagocytose WEHI-231 cells opsonized with complement (C3bi), which directly triggers CD11b, compared to the Ctrl mAb16 (Fig. 1g). This effect was prevented by a blocking CD11b mAb. Additionally, CD200 blockade enhanced the phagocytic capacity of mouse peritoneal and liver macrophages (Supplementary Fig. 1i). CD200R1 mAb OX-131, which blocks CD200R1 rather than CD200, also exhibited a phagocytic enhancement27 (Fig. 1h).
Therefore, CD200R1 was one of a few inhibitory receptors other than SIRPα that were co-immunoprecipitated with CD11b in macrophages. Antibody-mediated blockade of CD200R1-CD200 increased phagocytosis of tumor cells initiated by several pro-phagocytic receptors, including CD11b, FcRs and SLAMF7. The rapidity of these effects suggested that the inhibitory impact of CD200R1-CD200 on the functions of pro-phagocytic receptors may be direct.
Genetic evidence that CD200R1-CD200 suppresses phagocytosis
To confirm these results, we examined the impact of ablating expression of CD200R1 in macrophages, CD200 on tumor cells, or both (Fig. 2a; Supplementary Fig. 2a,b). Like the effects seen with the CD200 mAb, Cd200r1−/− BMDMs showed an increased phagocytosis of IgG-opsonized WEHI-231 cells in the presence of the Ctrl mAb, compared to WT BMDMs (Fig. 2b). There was no additional increase by adding the CD200 mAb. Furthermore, we generated a variant of WEHI-231 cells that lacked CD200 (Fig. 2a and Supplementary Fig. 2b). IgG-opsonized Cd200−/− WEHI-231 cells elicited enhanced phagocytosis by WT BMDMs compared to Cd200+/+ WEHI-231 cells (Fig. 2c). There was no further increase when BMDMs were deficient in CD200R1.

a Flow cytometry analysis of CD200R1 expression (left) on WT (top) or Cd200r1−/− (bottom) BMDMs, and CD200 expression (right) on Cd200+/+ (top) or Cd200−/− (bottom) WEHI-231 cells, as measured by flow cytometry. Red curves, CD200R1 or CD200 mAbs. Filled curves, Ctrl mAb. b As per Fig. 1d, except that phagocytosis of IgG-opsonized WEHI-231 cells in the presence of WT or Cd200r1−/− BMDMs was studied, (n = 3). c As per Fig. 2b, except that phagocytosis of IgG-opsonized Cd200+/+ and Cd200−/− WEHI-231 cells, in the presence of WT or Cd200r1−/− BMDMs, (n = 3). Data are from three independent experiments (a–c). Each symbol represents one mouse. Data are mean ± s.e.m. Statistical analysis: two-way ANOVA test, with multiple comparisons (b, c), ns not significant. See also Supplementary Fig. 2.
Thus, the absence of CD200R1 on macrophages or CD200 on tumor cells enhanced phagocytosis similarly to blockade of the CD200R1-CD200 interaction.
Blockade of CD200R1-CD200 inhibits tumor growth in vivo
To assess the effects of CD200R1-CD200 blockade in vivo, we used RAG-1-deficient mice (Rag1−/−), which lack T cells and B cells, and were inoculated with WEHI-231 or A20 cells. RAG-1-deficient mice were utilized, to prevent tumor cell rejection via T cell-mediated mechanisms, as all the available CD200+ tumor cell lines originated from mouse strains with a genetic background distinct from C57BL/6. In addition, this approach enabled us to avoid contributions of T cells and B cells to the anti-tumor effects of CD200R1-CD200 blockade. Once tumor growth was established, mice were treated with CD200 mAb or Ctrl mAb, plus Tac mAb 7G7 for opsonization.
Mice injected intravenously with WEHI-231 developed liver tumors (Fig. 3a–d, and Supplementary Fig. 3a). When euthanized at day 15, they displayed enlargement, white discoloration and tumor cell infiltration of the liver compared to those without tumor cell injection (Fig. 3b–d, Supplementary Fig. 3a). Compared to Ctrl mAb, treatment with CD200 mAb resulted in a notable decrease in these parameters (Fig. 3b–d). Flow cytometry analyzes of liver cells showed that the total number of WEHI-231 cells (defined as GFP+) was reduced by ~90% with the CD200 mAb compared to Ctrl mAb (Supplementary Fig. 3b, c). The total counts of immune cells, including macrophages, neutrophils, and NK cells, were also reduced (Supplementary Fig. 3c). Most immune cells in tumors were macrophages.

a–g Tac+ WEHI-231 tumor (with or without luciferase expression) were injected intravenously into Rag1−/− mice, followed by intraperitoneal injection of Tac mAb combined with CD200 mAb or Ctrl mAb every 2 days. Mice were euthanized on day 15 (b–d) or monitored over time using luminescence, if cells were expressing luciferase (e–g), (n = 12, b, c, n = 10, e–g). a Schematic representation of the experimental workflow. I.V., intravenously; I.P., intraperitoneally. b Representative photographs of mice euthanized on day 15, with or without tumor injection, and treated with the indicated mAbs. Scale bar, 1 cm. c Liver weight of mice injected or not with tumor cells. d Hematoxylin and eosin staining of liver sections, showing blood vessels (blue asterisks) and tumor cell aggregates adjacent to blood vessels (outlined in white). Scale bars, 100 µm (10× magnification), and 20 µm (40× magnification). e Representative luminescence images on day 12. f Tumor progression over time as measured by luminescence. sec, second, st, steradian. g Kaplan–Meier analysis of survival. h–j Tac+ A20 cells were injected subcutaneously in Rag1−/− mice followed by intraperitoneal injection of Tac mAb combined with CD200 mAb or Ctrl mAb, every 2 days, (n = 8). h Schematic representation of the experimental workflow. S.C, subcutaneously. i Tumor volume over time. j Tumor weight. Data are from three c or two d–j independent experiments, respectively. Each symbol represents one mouse. Data are presented as mean ± s.e.m. Statistical analysis: one-way ANOVA test, with multiple comparisons (c); two-tailed t-test (f, i, j). log-rank (Mantel–Cox) test (g). ns, not significant. See also Supplementary Figs. 3 and 4.
To evaluate the growth of WEHI-231 tumors in liver over time, we developed a variant of WEHI-231 that expressed luciferase (Supplementary Fig. 3a). Tumor mass was assessed by measuring luminescence. Compared to Ctrl mAb, the CD200 mAb caused a significant inhibition of tumor growth over time (Fig. 3e, f). Mouse survival was also improved (Fig. 3g). Importantly, the inhibitory effect of CD200 mAb on tumor growth was dependent on IgG opsonization of tumor cells, as it was not seen in the absence of mAb 7G7, compared to in the presence of mAb 7G7 (Supplementary Fig. 3d). It was also reliant on host macrophages, as its impact disappeared when mice were treated with clodronate-containing liposomes, which depleted >90% of liver macrophages, compared to control liposomes (Supplementary Fig. 3e–g).
In the case of A20 cells, RAG-1-deficient mice were injected subcutaneously with tumor cells and tumor size was ascertained over time using a caliper (Fig. 3h; Supplementary Fig. 4a). CD200 mAb yielded a reduction in A20 tumor size over time, compared to Ctrl mAb (Fig. 3i). Upon euthanasia, there was a decrease in tumor weight (Fig. 3j). Lastly, there was a diminution in the numbers of tumor cells and immune cells, most of the latter being macrophages (Supplementary Fig. 4b, c).
In summary, blockade of CD200R1-CD200 with CD200 mAb suppressed mouse tumor cell growth in vivo. This effect was dependent on IgG opsonization of tumor cells and required macrophages.
Distribution of CD200R1-CD200 in normal and malignant human cells
Although Cd200r1 is broadly expressed in mouse macrophages (www.immgen.org), whether this is also the case for human macrophages needs to be clarified. Unprimed blood monocyte-derived human macrophages exhibited low levels of CD200R1 (Fig. 4a and Supplementary Fig. 5a). However, IL-4, which induces differentiation into anti-inflammatory M2a-type macrophages31, augmented expression by ~5- to 10-fold. Interferon (IFN)-γ, which drives the differentiation into inflammatory M1-type macrophages, reduced expression, although the difference did not reach statistical significance33. Little to no effects were seen with IL-10 or transforming growth factor (TGF)-β. These observations were supported by analyzes of published RNA sequencing data, in which we noted that CD200R1 was among the most up-regulated transcripts following IL-4 treatment (Fig. 4b). They were also confirmed by our evaluation of a separate study with bulk RNA sequencing (Supplementary Fig. 5b)34. Additionally, they aligned with previous studies indicating that CD200R1 is a marker for M2a-type macrophages35.

a Flow cytometry analysis of CD200R1 expression on human blood monocyte-derived macrophages, either unprimed or primed for 1 day with the indicated stimuli. Red curves, CD200R1 mAb. Filled curves, Ctrl mAb. Number in histograms indicated mean fluorescence intensity (MFI). b Fold changes of RNA expression of human blood monocyte-derived macrophages treated with or without IL-4. CD200R1 is highlighted in red. Fold changes (log2; x-axis) and adjusted p-values (log10; y-axis) are shown. The red line represents a 4-fold change threshold. Data are from dataset GSE195440. c Frequency and relative expression levels of CD200R1 RNA across various human TAM subtypes, distinguished by gene expression profiles36,37. Data are from single-cell RNA sequencing (scRNA-seq) datasets GSE15476336 and GSE14677137 of TAMs from patients with the cancers indicated in Supplementary Fig. 5c, and colon carcinoma. d RNA expression profiles of human hematological malignancies, determined by microarray analysis. Left: UMAP plots of samples with tumor diagnosis are shown and colored by cluster identity. Middle: CD200 and CD47 RNA expression levels overlaid onto the UMAP (color gradient shows relative expression). Right: mean expression of CD200 and CD47 RNA in common subtypes of hematological malignancies and normal immune cells (color gradient). Data are from published dataset HEMAP56 (n = 7092). e Single-cell RNA sequencing analysis of human melanoma. Left: t-SNE profiles of melanoma malignant or non-malignant cells. Malignant melanoma cells are clustered by patient sample (top), while infiltrating immune cells are clustered by cell type (bottom). The middle and right: normalized expression of selected markers overlaid onto the t-SNE space, with a color gradient reflecting relative expression. Macrophages are outlined by a blue line. Data are from published dataset GSE11597857. f Same as d, except that expression of CD200 and CD47 RNA was analyzed in normal hematopoietic cells. g Flow cytometry analysis of expression of CD200 and CD47 on resting or activated human B cells, CD4+ T cells, and CD8+ T cells. Data are from three independent experiments (a, g). Each symbol represents one gene (b), one cell (c), one healthy donor or patient (d). Statistical analysis: Wald test followed by multiple comparisons (b); Kruskal–Wallis H test followed by Dunn’s post-hoc test with multiple comparisons (c, left); two-tailed Mann–Whitney U test (c, right panels). See also Supplementary Fig. 5.
To assess expression of CD200R1 in macrophages from human tumors, we took advantage of published RNA sequencing datasets. In these studies, the authors identified several tumor-associated macrophages (TAMs) subtypes, including SPP1+ (secreted phosphoprotein 1) TAMs, ISG15+ (interferon-secreted gene 15+) TAMs, INHBA+ (inhibin subunit beta A+) TAMs and C1QC+ (complement C1q C-chain) TAMs36,37 (Supplementary Fig. 5c). Amongst these, we found that C1QC+ TAMs showed the highest frequency of CD200R1 RNA expression, compared to other subtypes such as SPP1+, ISG15+ and INHBA+ TAMs (Fig. 4c). In contrast to CD200R1, SIRPA exhibited broader RNA expression across these various TAM subtypes (Supplementary Fig. 5d).
Of note, the authors of these datasets indicated that C1QC+ TAMs had an M2 gene expression signature with a high phagocytosis score characterized by expression of MRC1 (mannose receptor C-type 1), CD163, MERTK (MER Proto-Oncogene, tyrosine kinase) and CIQB (complement C1q B-chain), compared to the other subsets36,37 (Supplementary Fig. 5c). Gene ontology analyzes within the C1QC+ TAM population also revealed that CD200R1-positive macrophages expressed higher levels of genes associated with phagocytosis, compared to CD200R1-negative macrophages (Supplementary Fig. 5e).
In the case of CD200, databases of human hematological tumors demonstrated that CD200 RNA was most prevalent in B cell malignancies, such as CLL, MM, pre-B cell acute lymphoblastic leukemia (ALL), and Hodgkin’s lymphoma (HL), as well as in certain T cell malignancies like angioimmunoblastic T cell lymphoma (AITL) (Fig. 4d and Supplementary Fig. 5f). In contrast, CD47 RNA was broadly detected in all hematological tumors (Fig. 4d and Supplementary Fig. 5f). As an example of expression of CD200 in solid tumors, CD200 RNA was identified in tumor cells from ~50% of patients in a cohort of 14 melanoma cases, in keeping with previous reports18 (Fig. 4e). In contrast, melanoma cells universally expressed CD47. Analysis of the immune cells infiltrating these melanoma tumors also revealed that macrophages expressed CD200R1 and SIRPA.
In normal hematopoietic cells, CD200 also had a restricted expression pattern. CD200 was more highly detected in activated CD4+ T cells, resting B cells and activated B cells, with no expression in red blood cells, platelets and minimal or no expression in several cell types including resting CD4+ T cells, resting CD8+ T cells and activated CD8+ T cells (Fig. 4f). This expression pattern was confirmed by flow cytometry (Fig. 4g). By opposition, CD47 was broadly expressed in all normal hematopoietic cell types (Fig. 4f, g).
Therefore, human CD200R1 exhibited a narrower expression profile in macrophages compared to human SIRPα, being induced by IL-4 in blood-derived macrophages from healthy donors and primarily expressed in M2-type TAMs with high phagocytic capacity in human cancer tissues; likewise, human CD200 displayed a more restricted expression pattern compared to CD47, being present in B cell malignancies, specific T cell malignancies, and their normal counterparts, as well as in some solid tumors like melanoma.
Blockade of human CD200 enhances phagocytosis and reduces growth of human tumor cells
In line with the detection of CD200 RNA in specific human tumor types, CD200 was found on the surface of several human B cell lines, including NCI-H929 (MM), SLVL (B cell lymphoma), 721.221 (B lymphoblastoid cells), and KG-1a (acute myelogenous leukemia) (Fig. 5a). Additionally, it was detected on the human melanoma cell line SK-MEL-28 and the small cell lung cancer cell line NCI-H209 (Fig. 5a). The effects of CD200 blockade by therapeutic human CD200 mAb samalizumab on phagocytosis of these cells were assessed26. To ensure that samalizumab did not engage FcRs, we engineered an Fc-silent version of the mAb. When combined with IgG-opsonization, samalizumab enhanced phagocytosis by IL-4-pre-treated human macrophages, compared to Ctrl mAb (Fig. 5b). Little to no increase was observed in the absence of IgG-opsonization (Fig. 5b). The increase in phagocytosis in response to CD200 blockade by samalizumab was nearly equivalent to that seen with CD47 blockade by CD47 mAb B6H12 (Fig. 5c).

a Flow cytometry analysis of CD200 (top) or tumor antigens (CD20, CD38, CD123, DLL3 and SLAMF7; bottom) on various human tumor cell lines. Red curves, CD200 mAb or tumor antigen-specific mAbs. Filled curves, Ctrl mAbs. b As per Fig. 1d, except using IL-4-primed human blood monocyte-derived macrophages and human tumor cells in the presence of samalizumab (human IgG1, Fc-silent) or Ctrl mAb MOPC21 (human IgG1, Fc-silent). IgG opsonization was performed using CD20 mAb rituximab (SLVL, 721.221), CD38 mAb daratumumab (NCI-H929), CD123 mAb talacotuzumab (KG-1a), DLL3 mAb rovalpituzumab (NCI-H209), or SLAMF7 mAb elotuzumab (SK-MEL-28) (n = 3 or 4). c As per Fig. 5b, except using the indicated cell lines and samalizumab (human IgG1, Fc-silent), CD47 mAb B6H12 (human IgG1, Fc-silent), or Ctrl mAb MOPC21 (human IgG1, Fc-silent). Tumor cells were opsonized as detailed for Fig. 5b, (n = 3). d–f Subcutaneous injection of SLVL cells in NSG mice, followed by intraperitoneal injection of rituximab (mouse IgG2a version) combined with samalizumab or Ctrl mAb every 2 days, (n = 8). d Schematic representation of the experimental workflow. e Tumor volume over time. f Tumor weight. g–i Subcutaneous injection of 721.221 cells in NSG mice, followed by intraperitoneal injection of rituximab (mouse IgG2a version) combined with samalizumab or Ctrl mAbs every 2 days, (n = 10). g Schematic representation of the experimental workflow. h Tumor volume over time. i Tumor weight. Data are from three to four (a, b), three (c), or two (d-i) independent experiments. Each symbol represents one human sample or one mouse. Data are presented as mean ± s.e.m. Statistical analysis: two-tailed t-test (f, i), with multiple comparisons (b, c, e, h). ns, not significant. See also Supplementary Fig. 6.
To evaluate the impact of samalizumab on human tumor cell growth in vivo, SLVL or 721.221 cells were inoculated subcutaneously into NSG mice—deficient in T cells, B cells, and NK cells. Mice were then treated or not with samalizumab, in combination with anti-CD20 mAb rituximab for IgG opsonization. Compared to the Ctrl mAb, samalizumab significantly reduced the size and weight of tumors caused by SLVL or 721.221 cells and led to a decrease in the number of tumor cells and immune cells, which consisted of macrophages and to a lesser extent neutrophils, recovered from tumor tissues (Fig. 5d–i and Supplementary Fig. 6a–h).
Put together, these results showed that CD200 mAb samalizumab augmented phagocytosis and inhibited growth of human tumor cells in vivo, when combined with tumor-opsonizing mAbs. The impact of CD200 blockade on phagocytosis of human tumor cells was quantitatively analogous to that of CD47 blockade.
CD200R1 inhibits phagocytosis via Csk
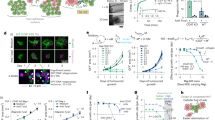
While SIRPα inhibits phagocytosis through ITIMs that recruit SHP-1 and SHP-2, CD200R1 lacks such motifs3,4. Instead, CD200R1 contains three conserved cytoplasmic tyrosines, Y286, Y289, and Y297, one of which, Y297, is part of the NPXY motif (Fig. 6a)14,15. When phosphorylated at Y297, the CD200R1 NPXY motif has been reported to bind the adaptor molecules Dok-1 and Dok-2, which subsequently enlisted the enzyme Ras-GAP, to suppress mast cell degranulation and myeloid cell cytokine production20,21,22,23,24.

a Partial sequences of the cytoplasmic ___domain of CD200R1 from different species. The conserved tyrosines (Y286, Y289, and Y297; based on mouse amino acid numbering) are in red, whereas the conserved NPxY motif is boxed. Identical residues are depicted by asterisks (*), while conserved and semi-conserved amino acids are highlighted by colons (:) and periods (.), respectively. b Phagocytosis of IgG-opsonized WEHI-231 cells by WT BMDMs expressing GFP alone, or Cd200r1−/− BMDMs expressing GFP alone or CD200R1 variants, in the presence of CD200 mAb or Ctrl mAb, (n = 3). c Mass spectrometry analyzes of cytoplasmic proteins with inhibitory potential interacting with synthetic biotinylated CD200R1 peptides, with or without phosphorylation at Y286 or Y297 in pull-down assays. Peptides are depicted at the top. Interactors identified by phosphorylated peptides are shown below. Negative regulators of immune cell activation, either adaptors, kinases, phosphatases or Ras-GAP, are indicated. d Immunoblot analysis of Dok-1 and Dok-2 expression (left) and phagocytosis of IgG-opsonized WEHI-231 cells (right) by BMDMs from WT or Dok1−/−Dok2−/− mice. β-actin as loading Ctrl (left). Normalized protein abundance (in %) relative to actin is shown below the top panel, (n = 4). e Same as Fig. 6d except that WT BMDMs transduced with Ctrl or Csk-specific siRNAs [Csk knockdown (CskKD)] were used. Two different Csk-specific siRNAs were studied. Csk expression (left) and phagocytosis (right) were studied, (n = 4). f Summary of fold-changes in phagocytosis for the various genetically deficient BMDMs in response to CD200 mAb, compared to WT BMDMs, (n = 3 or 4). g WT or Dok1−/−Dok2−/− BMDMs were stimulated or not for 1 min with biotinylated CD200R1 mAb OX-110 and streptavidin. Cell lysates were immunoprecipitated with α-Dok-1, α-Dok-2, α-Csk, or normal rabbit serum (NRS), and probed by immunoblotting with antibodies targeting phosphotyrosine (pTyr), Dok-1, Dok-2 or Csk. h WT or Dok1−/−Dok2−/− BMDMs were stimulated or not for 30 s with biotinylated CD200R1 mAb OX-110 and streptavidin. Cell lysates were probed with α-pLyn (Tyr 507) or α-Lyn Abs (top). A quantification of multiple independent experiments is shown at the bottom, (n = 3). Data are from three (b, e, g, h) or four d independent experiments, two (pY297 peptide) and three (pY286 peptide) (c) independent experiments. Each symbol represents one mouse (b, d, e). Data are mean ± s.e.m. Statistical analysis: two-way ANOVA test, with multiple comparisons (b, d, e, h); One-way ANOVA test, with multiple comparisons (f). ns not significant. See also Supplementary Figs. 7, 8 and Supplementary Table 2.
To investigate how CD200R1 suppressed phagocytosis, we created mutants of mouse CD200R1 with tyrosine-to-phenylalanine (Y-to-F) substitutions at positions Y286, Y289, or Y297. Additionally, we engineered a CD200R1 variant missing most of the cytoplasmic ___domain (Δintra-cytoplasmic; ΔIC)23,25,26,38. All mutations, along with WT CD200R1, were introduced into Cd200r1−/− BMDMs, where they expressed at levels approximately 10-fold higher than endogenous CD200R1 on WT BMDMs (Supplementary Fig. 7a). In comparison to Cd200r1−/− BMDMs expressing only GFP, those transduced with WT CD200R1 exhibited reduced phagocytosis, like WT BMDMs expressing GFP alone (Fig. 6b). This inhibitory effect was alleviated by CD200 mAb OX-90. Moreover, it was abrogated by the Y286F, Y297F, and ΔIC mutations, but not by the Y289F mutation.
To assess if Y286 and Y297 were recruiting inhibitors of phagocytosis, we used pull-down assays with synthetic peptides, phosphorylated or not, and lysates of mouse BMDMs, followed by mass spectrometry (Fig. 6c). The phosphorylated peptides, but not the unphosphorylated ones, recovered several proteins with inhibitory potential (Fig. 6c and Supplementary Table 2). These inhibitory proteins included the adaptors Dok-1, Dok-2, and Shc, Ras-GAP, the lipid phosphatase SHIP-1, protein tyrosine phosphatases SHP-1, and SHP-2, and the protein tyrosine kinase Csk.
Based on these findings, we examined the phagocytic function of BMDMs genetically lacking Dok-1 and Dok-2, Shc, Ras-GAP, SHIP-1, or SHP-1 and SHP-2. We also generated Csk-deficient BMDMs (Csk knock-down; CskKD) by transducing WT BMDMs with Csk-targeting small interfering (si) RNAs. We predicted that if any of these molecules were downstream effectors of CD200R1-mediated inhibition, we would observe increased phagocytosis without CD200 blockade, in addition to a reduction in the ability of CD200 blockade to enhance phagocytosis.
Compared to Ctrl BMDMs, the deficient BMDMs exhibited defective expression of the target proteins, but normal expression of various cell surface markers (Fig. 6d, e and Supplementary Figs. 7b–g and 8a–d). BMDMs lacking Shc, Ras-GAP, or SHP-1 and SHP-2 did not show any increase in phagocytosis in the absence of CD200 blockade or any impairment in the capacity of CD200 mAb to enhance phagocytosis, compared to the Ctrl mAb (Supplementary Fig. 8a–d). While SHIP-1-deficient BMDMs exhibited a small, albeit not statistically significant, increase in phagocytosis relative to Ctrl BMDMs in the absence of CD200 blockade, a further increase was still observed with the CD200 mAb, suggesting the possible involvement of SHIP-1 in CD200R1-independent inhibition of phagocytosis (Supplementary Fig. 8c). In contrast, BMDMs lacking Dok-1 and Dok-2, or those deficient in Csk, displayed increased phagocytic activity in the absence of the CD200 blockade compared to WT BMDMs. They also lost the ability of CD200 mAb to enhance phagocytosis (Fig. 6d, e). A summary of these data is shown in Fig. 6f.
As Dok-1 and Dok-2 bind NPXY motifs and have tyrosine phosphorylation sites that can recruit Csk, we tested if these adaptors were responsible for recruiting Csk to CD200R1. To this end, we analyzed the tyrosine phosphorylation of Dok-1 and Dok-2, along with their association with Csk in response to engagement of CD200R1. Compared to unstimulated cells, WT BMDMs stimulated with non-blocking CD200R1 mAb OX-110 exhibited prominent tyrosine phosphorylation of Dok-1 and Dok-2, which were also detected in Csk immunoprecipitates (Fig. 6g). No Dok-1 or Dok-2 phosphorylation or association with Csk was observed in BMDMs lacking Dok-1 and Dok-2 (Fig. 6g). As reported in earlier studies with the adaptor PAG38, although Dok-1 and Dok-2 were present in Csk immunopreciptates, we did not reciprocally observe Csk in immunoprecipitates of Dok-1 or Dok-2. This is likely related to the poor quality of the Csk antibodies available for immunoblot analyzes. In keeping with the involvement of Csk in Dok-1- and Dok-2-mediated inhibition, the CD200R1 mAb-induced phosphorylation of Lyn at the inhibitory tyrosine, Y507, which is the target of Csk, was abolished in BMDMs lacking Dok-1 and Dok-2, in comparison to Ctrl BMDMs (Fig. 6h).
Thus, the capacity of CD200R1 to inhibit phagocytosis involved a mechanism distinct from SIRPα that required the kinase Csk, which was recruited to CD200R1 via Dok-1 and Dok-2.
Synergy between CD200R1-CD200 and SIRPα-CD47 blockade in anti-tumor immunity
Since CD200R1 and SIRPα utilized different mechanisms of inhibition, we examined if combined blockade of CD200R1-CD200 and SIRPα-CD47 cooperated to augment phagocytosis. Mouse cell lines WEHI-231, A20, and J558 expressed both CD200 and CD47, albeit in different relative amounts (Fig. 7a). Single agent blockade of CD200R1-CD200 or SIRPα-CD47 with CD200 mAb or SIRPα mAb 27, respectively, yielded an increase in phagocytosis of IgG-opsonized WEHI-231, A20, and J558 cells, compared to Ctrl mAbs (Fig. 7b). Although doubling the concentrations of the individual mAbs did not increase phagocytosis further, combined blockade of CD200 and SIRPα did.

a Flow cytometry analysis of CD200 (red curves; top) and CD47 (lavender curves; bottom) expression on J558, A20 and WEHI-231 cells (left). Filled curves, Ctrl mAbs. The right panel shows relative expression levels of CD200 and CD47. b As per Fig. 1d, except that mAbs were used in combination: CD200 mAb OX-90 (rat IgG2a), SIRPα mAb 27 (mouse IgG2a, Fc-silent), Ctrl mAb 2A3 (rat IgG2a), and Ctrl mAb MOPC21 (mouse IgG2a, Fc-silent), (n = 3). c–f Luciferase+ Tac+ GFP+ WEHI-231 cells were injected intravenously into Rag1−/− mice, followed by intraperitoneal injection of Tac mAb combined with the indicated mAbs every 2 days starting from day 4, (n = 5). c Schematic representation of the experimental workflow. Tumor progression was measured over time using luminescence. Representative photographs of mice (d) and quantification (e). f Kaplan–Meier curves of survival. g Phagocytosis of normal activated human T cells or B cells by autologous human macrophages, in the presence of samalizumab (human IgG1, Fc-silent), CD47 mAb B6H12 (human IgG1, Fc-silent) or Ctrl mAb (human IgG1, Fc-silent). T cells were not opsonized (implying phagocytosis was mediated by SLAMF7), whereas B cells were opsonized with rituximab (human IgG1, Fc-active) (n = 3). Data are from three (a, b, g) or two (c–f) independent experiments. Each symbol represents one mouse or donor. Data are mean ± s.e.m. Statistical analysis: two-way ANOVA test, with multiple comparisons (b, e); log-rank (Mantel-Cox) test (f); one-way ANOVA test, with multiple comparisons (g). ns, not significant.
We also assessed synergic potential in vivo, using the WEHI-231 model expressing luciferase detailed in Fig. 3, with the exception that mice were treated with CD200 mAb, SIRPα mAb, or both. When used individually, CD200R1-CD200 blockade and SIRPα-CD47 blockade inhibited tumor growth and improved survival, compared to Ctrl mAbs (Fig. 7c–f). A greater impact was seen with CD200R1-CD200 blockade, compared to SIRPα-CD47 blockade. However, a much greater suppression was noted when CD200R1-CD200 blockade and SIRPα-CD47 blockade were combined.
In addition to causing anemia and thrombocytopenia, SIRPα-CD47 blockade can induce loss of lymphocytes, including CD8+ chimeric antigen receptor (CAR)-T cells, thereby potentially interfering with T cell-dependent anti-tumor responses39. Thus, we also compared the effects of blocking CD200R1-CD200 or SIRPα-CD47 individually on the phagocytosis of normal human T cells. In keeping with the expression patterns of CD200 and CD47 in human cells (Fig. 4f, g), blockade of SIRPα-CD47, but not of CD200R1-CD200, augmented phagocytosis of activated CD8+ T cells, which expressed CD47 but little to no CD200 (Fig. 7g). Either blockade strategy augmented phagocytosis of activated CD4+ T cells and B cells, which expressed both CD200 and CD47 (Fig. 7g).
Thus, combined blockade of CD200R1-CD200 and SIRPα-CD47 resulted in greater phagocytosis and inhibition of tumor growth in vivo compared to blocking of either checkpoint alone; moreover, unlike SIRPα-CD47 blockade, CD200R1-CD200 blockade did not enhance phagocytosis of activated CD8+ T cells.
Discussion
In this study, we found that engagement of CD200R1 on macrophages by CD200 on tumor cells inhibited phagocytosis in vitro and promoted tumor growth in vivo. CD200R1 suppressed phagocytosis initiated by several pro-phagocytic receptors, including FcRs, SLAMF7, and CD11b, presumably because of physical proximity between CD200R1 and CD11b, as suggested by the co-immunoprecipitation of CD200R1 with CD11b and as reported for SIRPα30,31. This inhibitory effect was demonstrated in several types of mouse macrophages, as well as in human blood monocyte-derived macrophages treated with IL-4. The effect was reversed by either antibody blockade of CD200R1 or CD200, or through genetic depletion of CD200R1 in macrophages or CD200 in tumor cells. Notably, within 15 min of acute disruption of the CD200R1-CD200 interaction, macrophages had enhanced actin polarization toward tumor cells, indicating that CD200R1’s influence on phagocytosis was rapid and likely direct.
There are several notable differences between CD200R1-CD200 and SIRPα-CD47. First, in humans, the expression patterns of CD200 and CD47 differed, both in tumor cells and in normal cells. While CD47 was essentially expressed on all tumor and normal cells, CD200 was restricted to a narrower range of cells, primarily B cell malignancies, some T cell neoplasia, and solid tumors such as melanoma, lung, breast, and prostate cancer18,19. Unlike CD47, CD200 was not found in red blood cells, platelets, and activated CD8+ T cells.
Second, expression of CD200R1 and SIRPα also showed significant differences. In human cells, CD200R1 expression was induced by IL-4 in human blood monocyte-derived macrophages, indicating its regulation by specific cytokines tied to immune responses. This regulation may be different in mice, where expression of Cd200r1 seems to be constitutive in macrophages (www.immgen.org). In human tissues, CD200R1 was primarily identified in C1QC+ TAMs, which were described to have M2-type features with high phagocytic capacity36,37. In contrast to human CD200R1, human SIRPα was constitutively expressed on blood monocyte-derived macrophages and was more uniformly present across TAMs. Although macrophages in the tumor microenvironment are often viewed as being poorly phagocytic, it should be noted that these cells, like pro-inflammatory macrophages, are capable of phagocytosis, especially when phagocytosis inhibitory checkpoints are blocked. This was already reported for SIRPα-CD47 blockade13,40. This notion was further supported by our data herein showing that IL-4-treated human macrophages displayed augmented phagocytosis in response to CD200 or CD47 blockade.
Third, the expression of CD200R1 and SIRPα diverged among immune cells other than macrophages. In addition to macrophages, human CD200R1 was reported to be present in T cells, NK cells, innate lymphoid cells (ILCs), mast cells, and dendritic cells (DCs)41,42. In contrast, SIRPα was additionally found in neutrophils, DCs, and NK cells. While the inhibition of tumor growth by CD200 mAbs in RAG-1-deficient mice and NSG mice indicated that the effects of CD200R1-CD200 blockade in our studies were independent of T cells, B cells, and, in NSG mice, NK cells, it remains possible that other immune cell types expressing CD200R1 will influence the overall effects of CD200R1-CD200 blockade in human patients. Along these lines, inhibitory effects of CD200R1 during T cell activation have been reported41,42. While the current study focussed on macrophages and phagocytosis, future studies, including with other mouse models and, especially, clinical trials in humans, will be needed for addressing the contributions of other macrophage functions and other types of immune cells to the responses of CD200R1-CD200 blockade in vivo.
Fourth, whereas SIRPα-CD47 suppresses phagocytosis via SHP-1 and SHP-2, CD200R1-CD200 inhibited phagocytosis through Csk43. This contrasts with the ability of CD200R1 to suppress mast cell degranulation and myeloid cell cytokine production, which were reportedly regulated by Ras-GAP20,21,22,23,24,25. The capability of CD200R1 to suppress phagocytosis also depended on two intracellular tyrosines in CD200R1, Y286 and Y297. Although the mechanism of action of Y286 remains to be clarified, one possibility, in keeping with the pull-down assays, is that Y286 was involved in recruiting the Src family kinase Lyn, which in turn was responsible for phosphorylating Y297, thereby enabling binding of Dok-1 and Dok-2 to phosphorylated Y297 and subsequent recruitment of Csk to Dok-1 and Dok-2. The divergence in molecular mechanisms between CD200R1 and SIRPα may explain the additive effects of combining CD200R1-CD200 and SIRPα-CD47 blockade in enhancing phagocytosis and suppressing tumor growth.
One of the questions emerging from our findings is whether Csk prevented or terminated phagocytosis. It is plausible that if Csk were already at the plasma membrane prior to engagement of a pro-phagocytic receptor, it would prevent activation before the activating signals are initiated. This is the case in T cells for PAG-associated Csk, which forms a complex prior to antigen-induced activation38. However, it is conceivable that the situation is different if the recruitment of Csk to the membrane occurs in response to cell activation, as was the case for CD200R1 in macrophages. In this case, CD200R1-associated Csk may terminate already initiated activating signals.
At first glance, the several differences between CD200R1-CD200 and SIRPα-CD47 may indicate that SIRPα-CD47 blockade would be effective against a wider range of tumor types, compared to CD200R1-CD200 blockade. Nonetheless, they also imply a higher potential for toxicity with SIRPα-CD47 blockade, due to the broader distribution of CD47 on normal cells. Thus, even though CD200R1-CD200 blockade may offer clinical efficacy against a narrower spectrum of tumor types, it may also have lower toxicity. For CD200+ tumors, the combination of CD200R1-CD200 blockade with SIRPα-CD47 blockade may also allow for lowering the dose of each blocking agent, thereby further reducing associated toxicities.
Despite all these distinctions, there is one key similarity between blockade of CD200R1-CD200 and blockade of SIRPα-CD47 that is therapeutically relevant. Like SIRPα-CD47 blockade, the CD200R1-CD200 blockade required the concomitant use of FcR-engaging anti-tumor antibodies to trigger phagocytosis, unless tumor cells expressed natural pro-phagocytic ligands like SLAMF7. Therefore, for most cancers, blockade of CD200R1-CD200, like that of SIRPα-CD47, will require combination therapies involving tumor-opsonizing mAbs. This is not surprising as blockade of inhibitory receptors on immune cells consistently requires co-engagement of activating receptors, either naturally or by therapeutic ligands, to evoke immune cell activation.
Data of phase 1 clinical trials with blocking CD200 or CD200R1 mAbs as T cell immune checkpoint blockers have been reported in patients with a variety of cancer types, including CLL, MM, and solid tumors25,26. The mAbs were well tolerated, indicating their safety. Additionally, in the samalizumab CD200 mAb trial with CLL and MM, over 60% of patients showed a noticeable clinical response, although only a small fraction experienced a reduction in tumor volume of 50% or more, and no complete responses were recorded. Such outcomes are common for single-agent therapies, particularly if immune checkpoint blockers primarily act on macrophages rather than T cells. The simultaneous use of Fc-active anti-tumor antibodies to trigger FcRs may be needed to achieve greater efficacy in clinical trials of CD200R1-CD200 blockade.
Previously, macrophage biology was dominated by the M1/M2 paradigm, with M1 macrophages being pro-inflammatory and anti-tumoral, and M2 macrophages promoting tumor progression31,33. However, single-cell RNA sequencing studies revealed that macrophages exhibit substantial heterogeneity and plasticity36,37. Indeed, in many preclinical mouse tumor models, TAMs exhibit immunosuppressive, pro-tumoral phenotypes36,37. However, in our models, macrophages seemingly played an anti-tumoral role, at least in the presence of phagocytic inhibitory checkpoint blockers. This is consistent with findings with other macrophage-targeting immunotherapies, such as SIRPα-CD47 blockade, which enhanced macrophage-mediated phagocytosis and suppressed tumor growth in preclinical models3,4,5. This contrasting functionality of macrophages is also reflected in ongoing clinical trials targeting different aspects of macrophage biology. For instance, anti-CSF1R antibody therapies aim to deplete immunosuppressive TAMs, while SIRPα-CD47 blockade therapies seek to enhance macrophage-mediated phagocytosis3,4,5,44.
In conclusion, our findings indicated that the CD200R1-CD200 checkpoint functions as a suppressor of phagocytosis in macrophages and serves as a promising macrophage target for treating CD200+ malignancies. Given the distinct molecular mechanism of action associated with CD200R1, the blockade of CD200R1-CD200 may be well-suited for combination with other inhibitory checkpoint blockers that target macrophages, potentially enhancing their efficacy while reducing toxicity. These insights also highlight the crucial need for sustained investigations into the mechanisms that regulate immune cell functions, which can then be better leveraged for developing effective and safer therapeutic strategies against human diseases.
Methods
Ethical statement
This research complies with all relevant institutional and national ethical regulations. All procedures involving human participants were approved by the IRCM Research Ethics Board, and informed consent was obtained from all participants. Animal studies were approved by the IRCM Animal Care Committee (protocol 2023-09) and conducted in accordance with the guidelines of the Canadian Council on Animal Care (CCAC).
Mice
Mice lacking SLAMF7 (Slamf7 −/−) or FcRγ (Fcer1g−/−) were generated in our laboratory, as described in refs. 12,31,40. The following mice were obtained from The Jackson Laboratory (Bar Harbor, ME): C57BL/6J, NSG (NOD.Cg-Prkdc scid Il2rg tm1Wjl/SzJ), RAG-1-deficient (Rag1−/−), CD18-deficient (Itgb2−/−), and Lys2-Cre. CD200R1-deficient mice (Cd200r1−/−), Dok-1- and Dok-2-deficient mice (Dok1−/−Dok2−/−), conditional SHIP-1-deficient mice (Inpp5dfl/fl), conditional Shc-deficient mice (Shc1fl/fl), conditional Ras-GAP-deficient mice (Rasa1fl/fl), conditional SHP-1-deficient mice (Ptpn6fl/fl), conditional SHP-2-deficient mice (Ptpn11fl/fl) and UBC-Cre-ERT2+ transgenic mice were reported45,46,47,48,49,50,51,52. Mice were maintained in the C57BL/6 background, unless specified. They were kept in a specific pathogen-free (SPF) environment. Either males or females were used, between 6 and 12 weeks of age. Littermates were used as controls in most experiments, except for some studies in which WT syngeneic, age and sex-matched mice were used. Housing was in Allentown PNC (positive/negative control) Model PNC 160 individually ventilated cages with Edstrom™ watering (Allentown Inc, Allentown, NJ, USA) in ventilated racks under negative pressure, ensuring a controlled environment at 20–23 °C and 40–60% humidity. An automated 12-h light/12-h dark cycle light system was used. Animal handling was performed in HEPA-filtered biosafety cabinets in a 100% filtered air ventilation room.
Cells
Mouse BMDMs and thioglycolate-elicited peritoneal macrophages were generated as described12,31. In brief, BMDMs were generated in medium supplemented with 30% (vol/vol) L929 cell-conditioned medium, which served as a source of colony-stimulating factor-1 (CSF-1). To deplete genes in UBC-creERT2 BMDMs in vitro, 0.5 μg/mL of 4 OH-tamoxifen (Sigma-Aldrich, St. Louis, MO) was added to the BMDM culture on day 2 and removed on day 3. To obtain thioglycolate-elicited peritoneal macrophages, mice were intraperitoneally injected with 1 mL of thioglycolate broth (BD Biosciences, Franklin Lakes, NJ). After 4 days, mice were euthanized, and peritoneal cells were collected via peritoneal lavage. Cells were first plated in a Petri dish for macrophage attachment. After 30 min, the suspended cells were removed, and the attached cells (macrophages) were harvested with 2 mM ethylene diamine tetra acetic acid (EDTA)-phosphate-buffered saline (PBS) and plated overnight into 24-well plates. Phagocytosis assays were performed the next day. To obtain liver macrophages, liver perfusion was done as described53. Liver was then dissected and digested in medium containing collagenase (1 mg/mL; Sigma-Aldrich) and DNase I (0.03 mg/mL; Sigma-Aldrich) at 37 °C for 30 min with constant agitation. Crude liver non-parenchymal cells (LNPCs) were enriched by centrifuging the suspension at 50 × g for 3 min at room temperature (RT), followed by centrifugation at 400 × g for 10 min at RT. Liver macrophages were generated by culturing LNPCs in medium supplemented with CSF-1 for 7 days.
Human blood samples were collected from healthy donors, above 18-year-old, both male and female, through the IRCM blood donor program. All participants provided written informed consent prior to participation, as approved by the IRCM Human Ethics Board. Peripheral blood mononuclear cells (PBMCs) were differentiated into macrophages by culture for 7 days in medium supplemented with 10% human serum and 10 ng/ml CSF-1, as reported40. Human monocyte blood-derived macrophages were either unprimed or primed with IFN-γ (100 ng/ml, PeproTech, Cranbury, NJ), IL-4 (10 ng/ml, PeproTech), IL-10 (50 ng/ml, PeproTech), or TGF-β (50 ng/ml, PeproTech) for 24 h prior to the phagocytosis assay. Human B cells, CD4+ T cells, and CD8+ T cells were isolated freshly or from frozen PBMCs using a negative selection method (STEMCELL Technologies, Vancouver, BC, Canada), achieving a purity of >90%. B cells were subsequently activated with lipopolysaccharide (LPS, 1 μg/mL; Sigma-Aldrich) for 2 days. CD4+ and CD8+ T cells were activated with Fc-silent anti-CD3ε (3 μg/mL, OKT-3) for 2 days.
WEHI-231, A20, J558, L1210, 70z/3, BW5147.3, YAC-1, S194/5, MPC-11, P815, B16-F10, CMT-93, L929, 293 T, NCI-H209, SK-MEL-28, and Phoenix-ECO cell lines were purchased from American Type Culture Collection (Rockville, MA). KG-1a and SLVL cell lines were purchased from RIKEN BioResource Center (Kyoto, Japan). NCI-H929 cell line was purchased from European Collection of Authenticated Cell Cultures (Salisbury, UK). 721.221 cell line was purchased from International Histocompatibility Working Group (Seattle, WA). The TUBO cell line was a gift from Dr. John Stagg, University of Montreal, Montreal, Canada. All cells were negative for Mycoplasma. They were certified as authentic by commercial providers, and identity was confirmed by assaying expression of relevant cell surface markers using flow cytometry.
DNA constructs, retroviral infection, and transfections
cDNAs encoding mutated versions of CD200R1 (Y286F, Y289F, Y297F, ΔIC) were generated by overlap-extension PCR. After cloning of WT or mutated cDNAs in the indicated retroviral vectors, plasmids were transfected in Phoenix-Eco cells, and viral supernatants were recovered. Supernatants were then used for spin infection of primary cells, as described31. Empty plasmids were used as controls. BMDMs were infected with retroviruses (produced with pFB-GFP) encoding the indicated CD200R1 variants. GFP-positive cells were sorted after 48 h of retroviral infection and propagated for an additional 24 h in growth medium before experimentation.
WEHI-231, A20, J558, and TUBO derivatives expressing Tac (human CD25) were generated using retroviruses, encoding a cytoplasmic ___domain-deleted version of Tac fused to the transmembrane ___domain of 2B4 (cloned into pFB-neo or pFB-GFP), followed by cell sorting31. Polyclonal Tac-expressing cells were isolated by cell sorting, whereas monoclonal populations were established by limiting dilution. CD200-deficient (Cd200−/−) WEHI-231 cells were generated using CRISPR-Cas9-mediated genome editing, with the guide RNA (gRNA) sequences 5′ TCTCCACCTACAGCCTGATT 3′ [Mm.Cas9.CD200.1.AE; Integrated DNA Technologies (IDT), San Diego, CA] or 5′ TTGTAGTGAAGGTGTACTAT 3′ (Mm.Cas9.CD200.1.AL; IDT). Cells transduced with scrambled gRNA (Alt-R® CRISPR-Cas9 Negative Control crRNA #1; IDT) were generated as control. In brief, gRNAs (10 μM) were assembled with Cas9 nuclease (62 μM, IDT) through incubation at RT for 15 min. Then, WEHI-231 cells (1 × 106) were resuspended in 20 μL buffer R, containing gRNA-Cas9 complexes, and cell electroporation was done using the Neon electroporation system (1000 V, 30 ms, 2 pulses). CD200-negative cells were sorted after 48 h. Csk expression was knocked-down using two different Csk-specific short-interfering (si) RNAs (mm.Ri.Csk.13.1, mm.Ri.Csk.13.2; IDT), which were transduced in BMDMs with Lipofectamine RNAi MAX (Invitrogen, Burlington, Ontario, Canada). Universal negative control siRNAs (Sigma-Aldrich) were used as control. In brief, 2 × 105 BMDMs were plated in a 6-well-plate. Then, 4 μL of 10 μM siRNAs were mixed with 12 μL Lipofectamine RNAi MAX in 200 μl Opti-MEM and added to BMDMs. Fresh medium was added after 6 h. Cells were used for phagocytosis or immunoblotting assays after 2 days.
Antibodies and fusion proteins
Recombinant mAbs were generated as described in ref. 12. In brief, cDNAs encoding the variable fragments of heavy chain (VH) and light chain (VL) of mAbs were synthesized by GeneArt (Invitrogen). Then, VH cDNAs were cloned in-frame into the expression vector pAb-hIgG1 or pAb-mIgG2a. Fc-silent versions carried the LALAPG mutation, i.e., leucine 234-to-alanine 234; leucine 235-to-alanine 235; proline 329-to-glycine 329. These point mutations, based on human IgG1 amino acid numbering, prevent binding to FcRs. The VL cDNAs were cloned in-frame into the expression vector pAb-hIgK or pAb-mIgK. Recombinant mAbs were expressed in CHO cells and purified with protein A Sepharose (Cytiva). Samalizumab sequences were obtained from patent US7408041B2, whereas rituximab sequences were from patent US5736137 (IDEC_C2B8). Control mAb MOPC21, CD47 mAb B6H12, and SLAMF7 mAb elotuzumab were described elsewhere12. Coomassie blue staining of protein gels was used to verify protein production and quality.
For phagocytosis assays and in vivo assays, the following mAbs were used: CD200 mAb OX-90 (BioXCell, Lebanon, NH); CD200R1 mAb OX-131 (Absolute Antibody, Boston, MA), SIRPα mAb no. 2712; rat IgG2b isotype Ctrl mAb LTF-2 (BioXCell); rat IgG2a isotype Ctrl mAb 2A3 (BioXCell); Ctrl mAb MOPC-21 (in-house generated); and blocking CD11b mAb 5C6 (BioXCell). For cell opsonization, the following mAbs were used: Tac mAb 7G7 (BioXCell); human CD20 mAb rituximab biosimilar (BioXCell); human CD123 mAb talacotuzumab (MedChemExpress, Princeton, NJ); human CD38 mAb daratumumab (MedChemExpress); human DLL3 mAb rovalpituzumab (MedChemExpress). For cell stimulation, the following mAbs were used: biotinylated CD200R1 mAb OX110 (Invitrogen) or human CD3ε mAb OKT-3 (hIgG1, Fc-silent). For flow cytometry, we used the following fluorophore-conjugated mAbs from Biolegend (San Diego, CA) or eBioscience (Waltham, MA): PD-L1 mAb 10 F.9G2; CD38 mAb 90; CD47 mAb miap301; CD19 mAb 6D5; F4/80 mAb BM8; CD200R1 mAb OX-110; CD200 mAb OX-90; CD45 mAb 30-F11; MHC class I H-2Kd mAb SF1-1.1.1; NK1.1 mAb PK136; Ly6G mAb 1A8; CD11a mAb M17/4; CD11b mAb M1/70; CD11c mAb N418; CD18 mAb M18/2; SIRPα mAb P84; CD64 mAb VX54-5/7.1; CD16/32 mAb 93; SLAMF7 mAb 4G2; CD20 mAb SA271G2; B220 mAb RA3-6B2; IgM mAb RMM-1; rat IgG2a isotype Ctrl mAb RTK2758; rat IgG1 isotype Ctrl mAb RTK2071; mouse IgG1 isotype Ctrl MOPC-21; Armenian hamster IgG isotype Ctrl mAb HTK888; rat IgG2b isotype Ctrl mAb RTK4530.
Immunoblotting was done using antibodies directed against Dok-1, Dok-2, Shc, Csk, SHP-1, SHP-2 or SHIP-1 Ab, which were generated in the Veillette laboratory38,54; CD11b (EPR1344, Abcam, Ontario, CA), CD200R1 (AF2554, R&D Systems, Minneapolis, MN), Ras-GAP (B4F8; Santa Cruz Biotechnologies, Dallas, TX), β-actin (C4, Santa Cruz Technologies), or p-Lyn (Tyr507) (Cell Signaling Technology, Danvers, MA) or Lyn (Cell Signaling Technology).
Flow cytometry
Cells were harvested and washed with 2% fetal bovine serum (FBS)-containing PBS (flow cytometry buffer). In most cases, to prevent non-specific binding to FcRs, FcRs were blocked with a mix of mouse IgG2a (mAb 7G7) and CD16/32 mAb 2.4G2 for 30 min on ice. Afterwards, cells were stained with fluorophore-conjugated antibodies (1:200 dilution, 1 × 107 cells/ml) for 30 min on ice. After being washed with flow cytometry buffer, samples were analyzed using a Fortessa flow cytometer (BD Biosciences).
To assess blocking of the CD200R1-CD200 interaction by CD200 mAb OX-90, 2.5 × 105 WEHI-231 cells were pre-incubated with CD200 mAb or Ctrl mAb for 30 min on ice. Subsequently, cells were incubated with Fc-silent variants of CD200R1-Fc or Ctrl Fc, carrying the LALAPG mutations, and labeled with Alexa Fluor 647, following the manufacturer’s instructions (Invitrogen). After incubation, cells were washed with flow cytometry buffer, and binding of the fusion proteins was analyzed by flow cytometry.
In vitro phagocytosis assays
Phagocytosis was assessed using either a microscopy-based assay or a pHrodo-based assay, as detailed31. For the microscopy-based assay, 5 × 104 macrophages were plated in a 24-well tissue culture plate. The following day, the culture medium was replaced with serum-free DMEM medium for 2 h. Meanwhile, the target cells were opsonized (or not) with antibodies at RT for 30 min, then labeled with 2.5 mM carboxyfluorescein succinimidyl ester (CFSE) fluorescent dye (Invitrogen). 2.5 × 105 CFSE-labeled target cells were co-incubated with macrophages for 2 h, in the presence of blocking or Ctrl mAb at 10 μg/ml. Bright field and fluorescent images were captured using an inverted microscope (Carl Zeiss Axiovert S100 TV). Phagocytosis efficiency was determined as the number of macrophages containing CFSE+ target cells per 100 macrophages. For phagocytosis using the combination of CD200 mAb and SIRPα mAb, the 1× concentration of each mAb was 10 μg/ml. To opsonize target cells with complement (C3bi), WEHI-231 cells were incubated with C5-deficient human serum (Sigma-Aldrich) for 1 h at 37 °C, before the microscopy-based phagocytosis assay.
For the pHrodo-based assay, 1 × 106 target cells were pre-labeled with 50 ng/ml pHrodo Red SE pH indicator (Invitrogen) in 100 μl PBS at RT for 30 min. Subsequently, 2.5 × 105 pHrodo-labeled target cells were co-incubated with macrophages for 2 h, in the presence of blocking or Ctrl mAb at 10 μg/ml. After 2 h, target cells and macrophages were harvested with Accutase (Innovative Cell Technologies, San Diego, CA), stained with F4/80 APC-Cy7, and processed for flow cytometry. Macrophages were identified as F4/80+ and GFPintermediate (macrophage autofluorescence). The pHrodo signal was acquired using the PE channel. Phagocytosis efficiency was calculated as the percentage of F4/80+ cells containing pHrodo+ cells. When monitoring phagocytosis over time using the Incucyte Live Cell Analyzer (Sartorius, Göttingen, Germany), 1 × 106 target cells were pre-labeled with 500 ng/ml pHrodo Red SE pH indicator (Invitrogen) in 100 μl PBS at RT for 30 min. Subsequently, 1 × 105 pHrodo-labeled target cells were co-incubated with macrophages (pre-labeled with CSFE) for 4 h, in the presence of blocking antibodies or Ctrl mAb at 10 μg/ml. Images were captured at 0, 1, 2, 3, and 4 h.
Conjugate formation and actin polarization
Conjugate formation and actin polarization assays were performed as previously described31. Briefly, 2 × 105 BMDMs were labeled with Cell Trace Violet (CTV) and plated overnight. The following day, 1 × 106 target cells were labeled with CSFE and co-incubated with BMDMs in serum-free DMEM medium in the presence of either CD200 mAb or Ctrl mAb for 15 min at 37 °C. After incubation, the dishes were washed, and cells were then fixed in PBS containing 4% paraformaldehyde solution, followed by permeabilization in PBS containing 0.1% Triton X-100. Subsequently, cells were blocked in PBS containing 5% BSA for 30 min at 4 °C. Following this, cells were washed and incubated with β-actin mAb AC-74 (Sigma-Aldrich) in PBS containing 5% BSA for 2 h at 4 °C. Subsequently, the cells were incubated with Alexa Fluor 647 anti-mIgG (Jackson ImmunoResearch Labs, West Grove, PA) for 1 h at 4 °C. Images were acquired using an LSM710 confocal microscope (Carl Zeiss). The degree of conjugation between BMDMs and target cells, relative to total BMDMs, as well as the percentage of cells with fully polarized actin at conjugation sites were quantified.
In vivo mouse tumor models
Tumor injections and treatments: For WEHI-231 and A20, pools of three clones of Tac+ GFP+ cells (1 × 106 each) were injected intravenously (WEHI-231) or subcutaneously (A20) in 6-10-week-old Rag1−/− mice. Alternatively, for WEHI-231, a polyclonal population of luciferase+ Tac+ GFP+ WEHI-231 cells (1 × 106) was used. At the indicated times after tumor cell injection, mice received intraperitoneal injections every 2 days of Tac mAb 7G7 or Ctrl mAb (200 µg), along with either CD200 mAb OX-90 (200 µg), SIRPα mAb 27 (200 μg), both mAbs, or Ctrl mAb 2A3 (200 µg). For macrophages depletion in vivo, mice were injected I.V. with 100 μl of clodronate liposomes or control liposomes (Liposoma BV, Amsterdam, Netherlands) every 4 days, starting from 1 day prior to tumor transplantation and until the end of the experiment.
For SLVL and 721.221 cells, cells (5 × 106) were injected subcutaneously in 6–10-week-old NSG mice. Prior to injection, SLVL tumor cells were resuspended in a 1:1 mixture of PBS and pre-warmed Matrigel (Corning Life Sciences, Glendale, AZ). At the indicated times after tumor cell injection, mice were treated with rituximab (mIgG2a variant, 200 μg) plus Fc-silent samalizumab or control mAb (200 μg), intraperitoneally.
Analysis: For A20, SLVL, and 721.221, tumor size was monitored with a caliper and was calculated using the formula (length × width2)/2. Mice were euthanized when tumor diameter reached 1.5 cm or more. For WEHI-231, mice were either euthanized at day 15 for liver examination or were monitored for tumor growth using luminescence. Upon euthanasia, the liver was dissected, weighed, and digested. Then, cells were fractionated by differential centrifugation, and red blood cells were lysed. After counting total cell numbers, populations of tumor and immune cells were identified by flow cytometry. For histology, liver tissues were fixed in 10% formalin, embedded in paraffin, sectioned, and stained with hematoxylin and eosin by the IRCM Histology Core Facility as described in ref. 55.
For mice injected with luciferase+ WEHI-231 cells, tumor progression was recorded every two days starting from day 4 post-inoculation. To this end, mice received 200 µL of a 15 mg/mL D-luciferin substrate (PerkinElmer, Woodbridge, ON) in PBS via intraperitoneal injection, followed by imaging using the IVIS™ Bioluminescence Imaging System (PerkinElmer). For survival analysis, mice were euthanized when the luminescence in the abdominal region reached 5 × 107 photons/s/cm2/steradian.
Immunoprecipitations, immunoblots, peptide pull-down assays, and mass spectrometry
For immunoprecipitation of CD11b and CD200R1 followed by immunoblotting, BMDMs from the WT, Cd200r1−/− and Itgb2 −/− mice were lysed by 1× Brij99 lysis buffer (150 mM NaCl;1% Brij99 (polyethylene [20]-oleylether); 25 mM HEPES (Gibco)), supplemented with protease and phosphatase inhibitors. The cell lysates were then centrifuged to remove insoluble material. Immunoprecipitation was performed using pre-cleared cell lysates and CD11b mAb M1/70 (Biolegend) or CD200R1 mAb OX110 (Biolegend), followed by capture with protein G sepharose (Cytiva). Beads were washed with lysis buffer containing 300 mM NaCl. For mass spectrometry, immunoprecipitates of CD11b from WT and Itgb2−/− mice were eluted using 0.1 M glycine (pH 2.8) buffer followed by neutralizing using 1 M Tris-HCl (pH 9.0). Three technical replicates were performed for each sample, and immunoprecipitates from Itgb2−/− mice were used as negative control.
Synthetic peptides corresponding to the cytoplasmic ___domain of mouse CD200R1 (residues 281 to 301), phosphorylated or not on the indicated tyrosines and containing biotin at the amino terminus, were obtained commercially (Genescript, Pistacaway, NJ). For experimentation, they were conjugated to NeutrAvidin™ Agarose (Thermo Fisher Scientific, Waltham, MA), followed by incubation with lysates of WT BMDMs in 1× TNE buffer (50 mM Tris, pH 8.0, 2 mM EDTA, 1% NP-40), supplemented with protease and phosphatase inhibitors. After incubation for 2 h at 4 °C, agarose beads were washed with 1× TNE buffer, then with PBS. Bound proteins were eluted using pH 2.8, 0.1 M glycine buffer for mass spectrometry. Two or three technical replicates were performed for each sample, with proteins pulled down by unphosphorylated peptides serving as negative control.
Mass spectrometry was performed by the IRCM Proteomics Core Facility, as outlined elsewhere12,31. In brief, samples were trypsin-digested and subjected to liquid chromatography-tandem mass spectrometry on an LTQ Orbitrap Velos (Thermo Fisher Scientific) equipped with a Proxeon nanoelectrospray ion source. A gradient of 100 min was used for liquid chromatography separation, and standard proteomics parameters were used for the mass spectrometers. Protein database searching was performed with Mascot (Matrix Science), and data analysis was conducted using the Scaffold software. Immunoprecipitates were analyzed by criteria for identifying potential CD11b interactors, included: (1) the candidates were 10-fold or greater more abundant in WT BMDMs, compared to Itgb2−/− BMDMs; (2) normalized Total Spectra of the protein candidates exceeded 1; (3) the candidates were identified in all of three independent experiments. Criteria for identifying potential CD200R1 peptide interactors included: (1) the candidates were 10-fold or greater more abundant with CD200R1 pY286 or pY297 peptides, compared to non-phosphorylated peptides; (2) normalized Total Spectra of the protein candidates exceeded 5; (3) the candidates were identified in all of two or three independent experiments.
To study protein phosphorylation, macrophages were stimulated or not with biotinylated CD200R1 mAb OX-110 at RT for 5 min, followed by streptavidin at 37 °C for additional 1 min. After stimulation, cells were lysed using 2× TNE buffer and analyzed by protein electrophoresis. Protein tyrosine phosphorylation was detected by immunoblotting of total cell lysates with phosphotyrosine (pTyr) mAb 4G10 (Sigma-Aldrich). Immunoprecipitation of Dok-1, Dok-2 or Csk was achieved using pre-cleared cell lysates and previously reported antibodies, which were produced in the Veillette laboratory, as reported elsewhere38. Normal rabbit serum (NRS) was used as control.
Expression of specific proteins was revealed by immunoblotting with antibodies directed against Dok-1, Dok-2, Shc, Csk, SHP-1, SHP-2, or SHIP-1 Ab, which were generated in the Veillette laboratory38,54; Ras-GAP (B4F8; Santa Cruz Biotechnologies, Dallas, TX); or β-actin (C4, Santa Cruz Technologies).
Published sequence data
Bulk RNA sequencing data of human blood monocyte-derived macrophages were normalized using the median of ratios method, followed by downstream analysis of differential expression conducted using PyDESeq2. For the study of CD200R1 or SIRPA RNA expression in human TAMs, single-cell RNA sequencing (scRNA-seq) data of TAM subtypes in multiple human cancer types were obtained from Cheng et al.36 and Zhang et al.37 For Cheng et al.36, only data for TAM subtypes identified in at least three human cancer types were used. CD200R1 and SIRPA RNA expression were depicted using violin plots. Differentially expressed genes (DEGs) in CD200R1-positive versus CD200R1-negative C1QC+ TAMs were analyzed using the Scanpy package (v1.9.6), applying a threshold of adjusted p-value of <0.05 and a fold-change ≥ 1.41. Gene Ontology (GO) enrichment analysis was performed using the gseapy package (v1.1.1).
For the study of CD200 and CD47 RNA expression in hematopoietic malignancies and normal cells, dataset annotation and log2-transformed gene expression data were extracted from Dufva et al.56 UMAP coordinates were utilized, where the color gradients for CD200 and CD47 represent the log2-transformed gene expression levels for each dataset. The datasets were grouped based on tumor or cell subtypes, and the mean log2 gene expression values for CD200 and CD47 were computed and depicted. In the study of human melanoma, dataset annotation and log2-transformed transcript-per-million (TPM) normalized scRNA-seq data were from Jerby-Arnon et al.57 The t-SNE coordinates were employed to visualize the data, with color gradients representing the log2-transformed expression levels of CD200 and CD47 in malignant cells, as well as CD200R1 and SIRPA in immune cell populations.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All the data are available within the article and its Supplementary Information. Protein mass spectrometry raw data in this study were deposited in ProteomeXchange repository PXD063538 and PXD063540. Bulk RNA sequencing data of human blood monocyte-derived macrophages were from the Gene Expression Omnibus (GEO) database GSE195440 and GSE227737. Single-cell RNA sequencing (scRNA-seq) data of TAM subtypes in multiple human cancer types were obtained from Cheng et al.36 (GSE154763) and Zhang et al.37 (GSE146771). Single-cell RNA sequencing of human melanoma dataset were from Jerby-Arnon et al. 57 (GSE115978). All reasonable requests for data will be fulfilled by [email protected]. Source data are provided with this paper.
References
Sharma, P. & Allison, J. P. Dissecting the mechanisms of immune checkpoint therapy. Nat. Rev. Immunol. 20, 75–76 (2020).
Baker, D. J., Arany, Z., Baur, J. A., Epstein, J. A. & June, C. H. CAR T therapy beyond cancer: the evolution of a living drug. Nature 619, 707–715 (2023).
Feng, M. et al. Phagocytosis checkpoints as new targets for cancer immunotherapy. Nat. Rev. Cancer 19, 568–586 (2019).
Veillette, A. & Chen, J. SIRPalpha-CD47 immune checkpoint blockade in anticancer therapy. Trends Immunol. 39, 173–184 (2018).
Advani, R. et al. CD47 blockade by Hu5F9-G4 and rituximab in non-Hodgkin’s lymphoma. N. Engl. J. Med. 379, 1711–1721 (2018).
Johnson, L. D. S. et al. Targeting CD47 in Sezary syndrome with SIRPalphaFc. Blood Adv. 3, 1145–1153 (2019).
Gracia-Hernandez, M., Suresh, M. & Villagra, A. The advances in targeting CD47/SIRPalpha “do not eat me” axis and their ongoing challenges as an anticancer therapy. Oncotarget 15, 462–465 (2024).
Lakhani, N. J. et al. Evorpacept alone and in combination with pembrolizumab or trastuzumab in patients with advanced solid tumours (ASPEN-01): a first-in-human, open-label, multicentre, phase 1 dose-escalation and dose-expansion study. Lancet Oncol. 22, 1740–1751 (2021).
Son, J. et al. Inhibition of the CD47-SIRPalpha axis for cancer therapy: a systematic review and meta-analysis of emerging clinical data. Front. Immunol. 13, 1027235 (2022).
Chen, Y. C., Shi, W., Shi, J. J. & Lu, J. J. Progress of CD47 immune checkpoint blockade agents in anticancer therapy: a hematotoxic perspective. J. Cancer Res. Clin. Oncol. 148, 1–14 (2022).
Freeman, S. A. & Grinstein, S. Phagocytosis: receptors, signal integration, and the cytoskeleton. Immunol. Rev. 262, 193–215 (2014).
Tang, Z. et al. CD47 masks pro-phagocytic ligands in cis on tumor cells to suppress antitumor immunity. Nat. Immunol. 24, 2032–2041 (2023).
Lin, G. H. Y. et al. TTI-621 (SIRPalphaFc), a CD47-blocking cancer immunotherapeutic, triggers phagocytosis of lymphoma cells by multiple polarized macrophage subsets. PLoS ONE 12, e0187262 (2017).
Kotwica-Mojzych, K., Jodlowska-Jedrych, B. & Mojzych, M. CD200:CD200R interactions and their importance in immunoregulation. Int. J. Mol. Sci. 22, 1602 (2021).
Choe, D. & Choi, D. Cancel cancer: the immunotherapeutic potential of CD200/CD200R blockade. Front. Oncol. 13, 1088038 (2023).
D’Arena, G. et al. CD200 and chronic lymphocytic leukemia: biological and clinical relevance. Front. Oncol. 10, 584427 (2020).
Alapat, D. et al. Diagnostic usefulness and prognostic impact of CD200 expression in lymphoid malignancies and plasma cell myeloma. Am. J. Clin. Pathol. 137, 93–100 (2012).
Petermann, K. B. et al. CD200 is induced by ERK and is a potential therapeutic target in melanoma. J. Clin. Investig. 117, 3922–3929 (2007).
Yoshimura, K. et al. CD200 and CD200R1 are differentially expressed and have differential prognostic roles in non-small cell lung cancer. Oncoimmunology 9, 1746554 (2020).
Zhang, S. & Phillips, J. H. Identification of tyrosine residues crucial for CD200R-mediated inhibition of mast cell activation. J. Leukoc. Biol. 79, 363–368 (2006).
Cherwinski, H. M. et al. The CD200 receptor is a novel and potent regulator of murine and human mast cell function. J. Immunol. 174, 1348–1356 (2005).
Zhang, S., Cherwinski, H., Sedgwick, J. D. & Phillips, J. H. Molecular mechanisms of CD200 inhibition of mast cell activation. J. Immunol. 173, 6786–6793 (2004).
Mihrshahi, R., Barclay, A. N. & Brown, M. H. Essential roles for Dok2 and RasGAP in CD200 receptor-mediated regulation of human myeloid cells. J. Immunol. 183, 4879–4886 (2009).
Akkaya, M., Aknin, M. L., Akkaya, B. & Barclay, A. N. Dissection of agonistic and blocking effects of CD200 receptor antibodies. PLoS ONE 8, e63325 (2013).
Fenaux, J. et al. 23ME-00610, a genetically informed, first-in-class antibody targeting CD200R1 to enhance antitumor T cell function. Oncoimmunology 12, 2217737 (2023).
Mahadevan, D. et al. Phase I study of samalizumab in chronic lymphocytic leukemia and multiple myeloma: blockade of the immune checkpoint CD200. J. Immunother. Cancer 7, 227 (2019).
Hayakawa, K. et al. CD200 restrains macrophage attack on oligodendrocyte precursors via toll-like receptor 4 downregulation. J. Cereb. Blood Flow. Metab. 36, 781–793 (2016).
Pfeifer, C. W. et al. Dysregulated CD200-CD200R signaling in early diabetes modulates microglia-mediated retinopathy. Proc. Natl. Acad. Sci. USA 120, e2308214120 (2023).
Hui, E. et al. T cell costimulatory receptor CD28 is a primary target for PD-1-mediated inhibition. Science 355, 1428–1433 (2017).
Morrissey, M. A., Kern, N. & Vale, R. D. CD47 ligation repositions the inhibitory receptor SIRPA to suppress integrin activation and phagocytosis. Immunity 56, 2172 (2023).
Chen, J. et al. SLAMF7 is critical for phagocytosis of haematopoietic tumour cells via Mac-1 integrin. Nature 544, 493–497 (2017).
Li, D. et al. SLAMF3 and SLAMF4 are immune checkpoints that constrain macrophage phagocytosis of hematopoietic tumors. Sci. Immunol. 7, eabj5501 (2022).
D’Agostino, G. et al. Beta-Arrestin1 and beta-Arrestin2 are required to support the activity of the CXCL12/HMGB1 heterocomplex on CXCR4. Front. Immunol. 11, 550824 (2020).
Gunalp, S. et al. TRAIL promotes the polarization of human macrophages toward a proinflammatory M1 phenotype and is associated with increased survival in cancer patients with high tumor macrophage content. Front. Immunol. 14, 1209249 (2023).
Koning, N. et al. Expression of the inhibitory CD200 receptor is associated with alternative macrophage activation. J. Innate Immun. 2, 195–200 (2010).
Cheng, S. et al. A pan-cancer single-cell transcriptional atlas of tumor infiltrating myeloid cells. Cell 184, 792–809.e723 (2021).
Zhang, L. et al. Single-cell analyses inform mechanisms of myeloid-targeted therapies in colon cancer. Cell 181, 442–459.e429 (2020).
Davidson, D. et al. The Csk-associated adaptor PAG inhibits effector T cell activation in cooperation with phosphatase PTPN22 and Dok adaptors. Cell Rep. 17, 2776–2788 (2016).
Yamada-Hunter, S. A. et al. Engineered CD47 protects T cells for enhanced antitumour immunity. Nature 630, 457–465 (2024).
Tang, Z. et al. Inflammatory macrophages exploit unconventional pro-phagocytic integrins for phagocytosis and anti-tumor immunity. Cell Rep. 37, 110111 (2021).
Schmiedel, B. J. et al. Impact of genetic polymorphisms on human immune cell gene expression. Cell 175, 1701–1715.e1716 (2018).
Monaco, G. et al. RNA-Seq signatures normalized by mRNA abundance allow absolute deconvolution of human immune cell types. Cell Rep. 26, 1627–1640.e1627 (2019).
Veillette, A., Latour, S. & Davidson, D. Negative regulation of immunoreceptor signaling. Annu. Rev. Immunol. 20, 669–707 (2002).
Liu, D. et al. A comprehensive review on targeting diverse immune cells for anticancer therapy: beyond immune checkpoint inhibitors. Crit. Rev. Oncol. Hematol. 210, 104702 (2025).
Boudakov, I. et al. Mice lacking CD200R1 show absence of suppression of lipopolysaccharide-induced tumor necrosis factor-alpha and mixed leukocyte culture responses by CD200. Transplantation 84, 251–257 (2007).
Mashima, R. et al. Mice lacking Dok-1, Dok-2, and Dok-3 succumb to aggressive histiocytic sarcoma. Lab Investig. 90, 1357–1364 (2010).
Celis-Gutierrez, J. et al. Dok1 and Dok2 proteins regulate natural killer cell development and function. EMBO J. 33, 1928–1940 (2014).
Tarasenko, T. et al. T cell-specific deletion of the inositol phosphatase SHIP reveals its role in regulating Th1/Th2 and cytotoxic responses. Proc. Natl. Acad. Sci. USA 104, 11382–11387 (2007).
Hardy, W. R. et al. Combinatorial ShcA docking interactions support diversity in tissue morphogenesis. Science 317, 251–256 (2007).
Lapinski, P. E. et al. Generation of mice with a conditional allele of the p120 Ras GTPase-activating protein. Genesis 45, 762–767 (2007).
Pao, L. I. et al. B cell-specific deletion of protein-tyrosine phosphatase Shp1 promotes B-1a cell development and causes systemic autoimmunity. Immunity 27, 35–48 (2007).
Yasuda, T. et al. Role of Dok-1 and Dok-2 in myeloid homeostasis and suppression of leukemia. J. Exp. Med. 200, 1681–1687 (2004).
Li, Y. T., Wu, H. L. & Wu, L. L. Isolation of murine hepatic myeloid cells with high yield and purity using immunomagnetic beads for subset analysis. STAR Protoc. 4, 102592 (2023).
Davidson, D., Shi, X., Zhong, M. C., Rhee, I. & Veillette, A. The phosphatase PTP-PEST promotes secondary T cell responses by dephosphorylating the protein tyrosine kinase Pyk2. Immunity 33, 167–180 (2010).
Wu, N. et al. Critical role of lipid scramblase TMEM16F in phosphatidylserine exposure and repair of plasma membrane after pore formation. Cell Rep. 30, 1129–1140.e1125 (2020).
Dufva, O. et al. Immunogenomic landscape of hematological malignancies. Cancer Cell 38, 424–428 (2020).
Jerby-Arnon, L. et al. A cancer cell program promotes T cell exclusion and resistance to checkpoint blockade. Cell 175, 984–997 e924 (2018).
Acknowledgements
We thank members of the Veillette laboratory for useful discussions. We also thank Gaspard Reulet and Caroline Grou for help with the bioinformatics analyzes. This work was supported by grants from the Canadian Institutes of Health Research (MT-14429, MOP-82906, FDN-143338, PJT-178314 and PJT-183593), the Terry Fox Research Institute (1190-02) and the Ministère de l’économie et de l’innovation (MEI; Québec) to A.V. The mass spectrometry analyzes of CD11b were performed by Jun Chen. J.L. and X.Q. are supported by a studentship from the Chinese Scholarship Council. Z.T. received a Fellowship from the Cole Foundation, and A.V. held the Canada Research Chair on Signaling in the Immune System. J.A.N. was supported by Fondation ARC pour la recherche sur le Cancer (PJA20161204835).
Author information
Authors and Affiliations
Contributions
J.L., Z.W., M.-C.Z., Z.T., J.Q., D.D., and A.V. designed experiments. J.L., Z.W., X.Q., M.-C.Z., J.Q., and J.D. performed experiments or analyzed data. T.H., P.D.K., J.A.N., and Y.Y. provided reagents. All authors interpreted the results. J.L. and A.V. wrote the manuscript. All authors commented on the manuscript. A.V. and J.A.N. provided funding.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Li, J., Wang, Z., Qin, X. et al. CD200R1-CD200 checkpoint inhibits phagocytosis differently from SIRPα-CD47 to suppress tumor growth. Nat Commun 16, 5145 (2025). https://doi.org/10.1038/s41467-025-60456-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-025-60456-3
