
Abstract
The origin of pterosaurs, the first vertebrates to achieve powered flight, is poorly understood, owing to the temporal and morphological gaps that separate them from their closest non-flying relatives, the lagerpetids. Although both groups coexisted during the Late Triassic, their limited sympatry is currently unexplained, indicating that ecological partitioning, potentially linked to palaeoclimate, influenced their early evolution. Here we analysed pterosauromorph (pterosaur + lagerpetid) palaeobiogeography using phylogeny-based probabilistic methods and integrating fossil occurrences with palaeoclimate data. Our results reveal distinct climatic preferences and dispersal histories: lagerpetids tolerated a broader range of conditions, including arid belts, enabling a widespread distribution from the Middle to early Late Triassic. Conversely, pterosaurs preferred wetter environments, resulting in a patchier geographical distribution that expanded only as humidity increased in the Late Triassic, probably following the Carnian Pluvial Event. This major environmental disturbance, potentially driven by changes in CO2-related thermal constraints and/or palaeogeography, appears to have had a key role in shaping early pterosauromorph evolution by promoting spatial segregation and distinct climatic niche occupation.
Similar content being viewed by others


Main
Pterosaurs were the first vertebrates to evolve powered flight, more than 60 million years before the earliest birds. However, our understanding of early pterosaur evolution is hindered by the major temporal and anatomical gaps between these highly modified flying reptiles and their closest terrestrial relatives1,2. The fossil record of pterosaurs and their kin is notoriously incomplete3,4,5,6,7,8,9,10,11 (Supplementary Fig. 1), and fundamental aspects of their early evolution, such as the timing, area, ecological settings of their initial radiation and palaeobiology (for example, growth dynamics and climate preferences1,12,13), are still poorly understood compared with other contemporaneous archosaur groups such as dinosaurs14,15,16.
Recent phylogenetic studies, descriptions of new taxa and detailed reevaluations of historically described specimens have renewed interest in pterosaur origin, ancestry and early evolution. This work overturned previous hypotheses of pterosaur relationships, demonstrating that lagerpetids were the closest relatives of pterosaurs, uniting these two groups in the clade Pterosauromorpha2,11,17,18,19. All known lagerpetids are non-flighted forms, whereas all known pterosaurs are volant. This new phylogenetic hypothesis substantially reduces temporal and anatomical gaps between Pterosauria and other avemetatarsalians (pterosaurs, dinosaurs and their closest relatives)2,11,18,19. Consequently, the origin of Pterosauria is now minimally constrained to an interval of ~10–15 million years between the Ladinian and early Norian intervals of the Triassic Period2,11,18,20. The past decade of research has also provided a wider sample of Triassic pterosauromorph specimens from more localities across the globe and a better understanding of their phylogenetic relationships1,2,11,17,18,19,20 and stratigraphic occurrences. These advances offer the chance to investigate early pterosauromorph macroevolution quantitatively (for example, ref. 11), by uncovering potential abiotic factors that might have influenced their diversification, as hinted at by the stratigraphic occurrence and disjunct palaeogeographical distributions of lagerpetids and pterosaurs through the Triassic18.
Sympatric occurrences of lagerpetids and pterosaurs were limited during their ~20-million-year overlap in the Norian and Rhaetian (Fig. 1). Lagerpetids are found in continental fluviolacustrine deposits on at least four current continents and maintained a wide latitudinal and geographic spread throughout most of their evolutionary history11,17,18,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34 (late Ladinian to late Rhaetian, 237–201 Ma; Fig. 1 and Supplementary Fig. 1). Conversely, the earliest body fossils of unambiguous pterosaurs are middle Norian in age and found in a restricted low-latitude belt, primarily, but not exclusively, in marine formations that were deposited around the margins of Tethys1,12,35,36,37. From the late Norian onwards, pterosaurs spread to higher latitudes and in a much broader array of habitats1,3,8,12,38,39,40,41,42. This could suggest that lagerpetids and pterosaurs occupied fundamentally different ecological, environmental or climatic niches. Investigating pterosauromorph distribution in time and space combined with environmental data allows a new opportunity to answer several long-standing key questions in palaeontology: how did Triassic climate change affect pterosaur and avemetatarsalian evolution? What climatic regimes and environments did vertebrate flight first evolve in? And why are early pterosaur fossils so rare, and where might we find more of them?

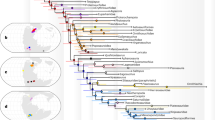
Middle–Late Triassic pterosauromorph occurrences worldwide (left) and close-ups of the southwestern USA (right). An., Anisian; Jr., Jurassic.
We investigated these questions using a holistic approach that integrates novel fossil occurrence data, comprehensively sampled phylogenies, climatic niche modelling and palaeobiogeography. Our aims are (1) to assess and quantify the potential for latitudinal dispersal of Triassic lagerpetids and pterosaurs and to compare it with that of other contemporaneous groups; (2) to investigate climatic preferences of pterosauromorphs; and (3) to quantify and map habitat suitability for pterosaurs and their closest relatives during the Middle to Late Triassic. Furthermore, our results allow us to estimate the ancestral area of origin and hypothetical distribution and dispersal of pterosauromorphs across Pangaea during the Middle Triassic to early Late Triassic—the undersampled time interval in which their origin probably unfolded—to propose testable hypotheses regarding their early biogeography and its potential influence on their evolution and to identify potential target areas for future fieldwork.
Results
Palaeobiogeography
Our palaeobiogeographic analysis suggests that lagerpetids—and pterosauromorphs as a whole—probably originated in southwestern Pangaea (that is, modern South America), whereas the origin of pterosaurs was predicted at low latitudes in the Northern Hemisphere (Extended Data Fig. 1), consistent with previous studies8,11. Note that this and the following results are based on phylogenetic data, which, in light of the biases and incompleteness of the fossil record may simplify what are probably more complex scenarios (Supplementary Fig. 1).
Potential of latitudinal dispersion
The pterosauromorph dispersal inertia curve maintains a consistently high Δ-log likelihood (that is, lower but constant dispersal) across the Ladinian–early Norian interval (Fig. 2a). In other words, the dispersal of lagerpetids (the only pterosauromorphs present at that time) across climatic barriers was constant but more constrained than that of dinosaurs (and avemetatarsalians as a whole). Dinosaurs experienced relatively stronger barriers to dispersal in the Ladinian–early Carnian and late Norian–Rhaetian, interrupted by a ‘release phase’ (that is, increased levels of dispersal) during the middle–late Carnian14 (Fig. 2a). The middle Norian–Rhaetian segment of the pterosauromorph curve shows an increase in Δ-log likelihood, potentially indicating a reduced crossing of barriers through this interval (Fig. 2a). This section of the curve is primarily, if not exclusively, influenced by the addition of pterosaurs (Methods). The pattern of reduced dispersal is surprising given that Triassic pterosaurs were volant and many are found in low-latitude areas (Fig. 1, Extended Data Fig. 1 and Supplementary Fig. 1). A similar pattern (that is, increase in species richness and drop in dispersal) could be achieved by few intermittent crossings of the barriers, which are unlikely to alter the dispersal pattern, followed by cladogenesis within the same geographical area, which will not affect the dispersal through barriers. Nevertheless, as emphasized in the Methods, the importance of this segment of the curve remains uncertain. As pterosaurs could fly and were presumably less constrained by physical barriers, their likelihood for dispersal may have left a less clear phylogenetic signature compared with their non-flying close relatives.

a, A plot showing the potential of dispersal of avemetatarsalian groups through the Triassic (based on Griffin et al.14). b, Absolute accumulated degrees of latitudinal dispersal of avemetatarsalian groups. c, Event-corrected accumulated degrees of latitudinal dispersal of avemetatarsalian groups. J., Jurassic; E. Tr., Early Triassic; I., Induan; Olenek., Olenekian.
Accumulated latitudinal dispersion
A complementary way to decipher the latitudinal dispersion of different groups is by quantifying dispersal events in each clade through time11 (Fig. 2b,c). This shows that lagerpetids dispersed consistently throughout their evolutionary history (Anisian–Rhaetian). The lagerpetid curve for accumulated latitudinal dispersion is high both in absolute (Fig. 2b) and corrected values (Fig. 2c). These curves maintain a plateau throughout the Carnian and Norian and decrease only in the Rhaetian, when the taxonomic diversity of the group declines. This pattern is similar to that of silesaurids (Discussion), contrary to the results of Müller et al.11, whose lagerpetid curve peaked after the Carnian and was more similar to that of dinosaurs. This inconsistency is caused by (1) the pruning by Müller et al.11 of key lagerpetids (Kongonaphon kely and PVSJ 883) from their dataset, which reduced the number of branches and dispersal events in the Anisian–Carnian interval, thus lowering that part of the curve; and (2) the different strategy of Müller et al.11 of incorporating uncertain biogeographical values at internal nodes, which led them to discard the dispersal events at these particular nodes. The accumulated latitudinal dispersal of pterosaurs peaks immediately after their appearance in the middle Norian and maintains a high profile throughout the Norian and Rhaetian11 (Fig. 2b,c). The abrupt and pronounced Norian peak in the pterosaur curve may underscore the inherent ease of dispersal for these flying reptiles compared with their land-bound counterparts.
Palaeoclimate niche occupation
Statistical comparisons reveal that pterosaurs and lagerpetids occupied different climatic niche spaces during their temporal overlap in the Late Triassic (Fig. 3 and Extended Data Table 1; pairwise permutation multivariate analysis of variance (MANOVA), P = 0.0127). During the Norian and Rhaetian, lagerpetids occupied areas characterized by warmer temperatures and drier conditions and with less pronounced seasonal excursions in temperatures (Fig. 3b–d). Comparatively, pterosaurs occurred in areas that were distinctly cooler and seasonally more variable (Fig. 3b,d and Extended Data Table 1; P < 0.01, P < 0.05). Both lagerpetids and pterosaurs occupied areas with similar values of mean annual precipitation (MAP; Fig. 3c and Extended Data Tables 1 and 2; P > 0.05), although a majority of lagerpetids occupied drier areas (Fig. 3c). Pterosaurs occurred across a wide range of values for annual precipitation and seasonal variation in precipitation (Fig. 3c,e). The palaeoclimate niche occupation of lagerpetids was not constant over time (Fig. 3f). Specifically, Ladinian and Carnian lagerpetids (that is, Kongonaphon, Ixalerpeton, Lagerpeton and PVSJ 883) are found in high-latitude areas in southern Gondwana that were on average colder and withstood higher seasonal fluctuations in temperatures than their Norian and Rhaetian counterparts (Fig. 3 and Extended Data Table 1; pairwise permutation MANOVA, P = 0.012). This pattern is not entirely surprising considering the palaeogeographical occurrence of Ladinian and Carnian lagerpetids compared with their younger counterparts. Most Norian–Rhaetian occurrences are, in fact, found at lower latitudes that were characterized by higher temperatures and smaller seasonal fluctuations in temperature (Fig. 3b,d and Extended Data Table 1; P < 0.001), with the exception of Faxinalipterus and Dromomeron gigas, which were found in the southern part of Pangaea, in geographical areas and palaeoclimatic regimes broadly similar to those of their pre-Norian predecessors (Discussion, Figs. 1 and 3f and Supplementary Fig. 1). As a result, lagerpetids as a whole occupied an overlapping (pairwise permutation MANOVA, P = 0.098; Extended Data Fig. 2 and Extended Data Table 1) but broader palaeoclimate niche than pterosaurs, probably related to their higher tolerance for warmer, more seasonal (temperature-wise) and tendentially drier conditions (Extended Data Fig. 2 and Extended Data Tables 1 and 2).

a, Principal component analysis (PCA) of variables of climatic importance showing pterosaurs (n = 23), pre-Norian lagerpetids (n = 9) and Norian–Rhaetian lagerpetids (n = 17). b–e, Raincloud plots comparing climatic variables between pterosaur and lagerpetids: mean annual temperature (b); mean annual precipitation (c); seasonal variation in temperature (d); seasonal variation in precipitation (e). f, The time-sliced PCA space occupation of lagerpetids and pterosaurs through the Triassic. Results of statistical comparisons are reported in Extended Data Table 1. P, precipitation; T, temperature. The box plots display the median and interquartile range (IQR), the upper whiskers extends from the 75th percentile to the largest value and the lower whiskers extends from the 25th percentile to the lowest value.
Habitat suitability modelling
Climatic suitability offers a route to interpret the palaeobiogeographical distributions of early pterosauromorph body fossils and allows us to identify areas of suitable habitable space in fossil-depleted intervals. It provides a coarse, but useful, representation of their hypothetical fundamental niches (Fig. 3) in geographic space (Fig. 4 and Extended Data Fig. 3). The temporal and spatial biases that control the early pterosaur fossil record epitomize the challenges faced in obtaining a comprehensive understanding of their evolutionary history and geographic distribution (Supplementary Fig. 1). Nevertheless, a probabilistic macroecological approach can provide insights on their climatic preferences and potential habitable areas, predicting potential geographic ranges for pterosaurs and lagerpetids based on climatic suitability. The distribution of areas of high suitability of pterosaurs and lagerperids appear broadly complementary. However, pterosaur potential distribution is patchier overall with lower habitability in continental interior areas (that is, arid regions) compared with the more widely distributed of lagerpetids. Although pre-Norian pterosaur body fossils have yet to be recovered, ecological niche modelling indicates the presence of potential suitable habitable space for hypothetical early pterosaurs that was unevenly spread at low tropical latitudes during the Ladinian (Fig. 4 and Extended Data Fig. 3), compared with the occupation of higher, more temperate latitudes in the Northern Hemisphere and broader and more continuous suitable space occupation in the southwestern tropics, for lagerpetids (Fig. 4). Habitat suitability modelling also indicates a marked extension of habitable pterosaur space towards the coastlines of Northern Tethys in the Carnian–early Norian (Fig. 4), alongside a reduction of suitable continental zones, emphasizing their climatic preference for the more temperate and wetter environments (Extended Data Figs. 3 and 4). By contrast, lagerpetids display a distinct Carnian pattern, maintaining a more homogeneous latitudinal occupation but with decreasing habitat suitability along the southern margin of the northern Tethys gulf, while simultaneously exhibiting an increasingly higher preference towards the southwestern tropics and reaching slightly higher latitudes in the Southern Hemisphere (Fig. 4). During the Norian (our ‘training stage’; Fig. 4 and Extended Data Fig. 3), pterosaurs (now represented by body fossils) were widely distributed in coastal, wet, tropical and low-latitude areas (Figs. 1 and 4). This is accompanied by the continuing patterns of reduction in suitable continental interior areas, except alongside the palaeoequatorial belt (broadly corresponding to the Chinle and Dockum basins) where favourable conditions for the group are maintained. Overall, in the Norian and Rhaetian, areas of pterosaur high climatic suitability occur at broad latitudes, extending from the northern margin of the Tethys to northern Australia and central South America (that is, southern Brazil and northern Argentina). Low suitability is recorded at high latitudes outside the tropics and in what is now equatorial Africa, Antarctica, Oceania and northeastern Eurasia and continental interiors (for example, modern Brazil and Canada–USA border). This pattern is broadly maintained through the late Norian and Rhaetian with an expansion of high suitable zones along the eastern margin of Pangaea (Fig. 4 and Extended Data Figs. 3 and 4). By contrast, lagerpetid suitability habitats are maintained throughout the Norian–Rhaetian interval with the addition of the western palaeoequatorial belt (that is, Chinle and Dockum basins) and northwestern Africa.

Note that the pre-Rhaetian suitability maps are modelled predictions because, so far, there is no Ladinian–Carnian fossil record of pterosaurs.
Discussion
Palaeobiogeography, palaeoclimate and ecological niches
Triassic pterosauromorph distribution suggests that pterosaurs and lagerpetids had distinct and divergent early evolutionary histories, with regard to biogeography and dispersal. Lagerpetids achieved a wide latitudinal distribution soon after their first appearance (predominantly in southern Pangaea) and maintained a presence at high and low latitudes, globally, throughout their evolutionary history10,11,18 (Fig. 1 and Supplementary Fig. 1). The clade’s dispersal across climatic barriers remained low and constant through time (Fig. 2), with our results suggesting that lagerpetids had a broad tolerance for a range of different climatic conditions, for example, higher thermal tolerances and/or other behavioural strategies to deal with high temperatures. By contrast, the earliest pterosaurs are found in low-latitude lacustrine and fluvial beds of the Chinle and Dockum basins during the early and middle Norian and more predominantly (but not exclusively) in coastal habitats of the northern Tethyan region3 (Fig. 1 and Supplementary Fig. 1). However, within a few million years, pterosaurs are then found worldwide in remarkable variety of environments including many beyond low-latitudinal coastal regions, as shown by Arcticodactylus from fluvial deposits in Greenland (Fleming Fjord Formation, Norian–Rhaetian)38, Caelestiventus from the deserts of southwestern USA (Nugget Sandstone, upper Norian–Hettangian)41 and Pachagnathus and Yelaphomte from Argentina (Quebrada del Barro Formation, upper Norian–lower Rhaetian)20. In line with this clade’s dramatic peak of dispersal in the middle Norian (Fig. 2), it is possible that these younger pterosaur occurrences are evidence of the clade’s first dispersals outside of its hypothesized low-latitude ancestral area. As such, these occurrences might represent successive invasions of diverse habitats following a climatically driven ecological release (see below), rather than evidence of ancestral preference. The hypothesis of a rapid global spread8 from low latitudes finds support in (1) the phylogenetic predominance of low-latitude taxa closer to the root of the clade (2) and the ~6-million-year gap between the earliest identifiable pterosaurs in the fossil record (that is, SMU 69125, and PEFO 45782 from the early Norian of the southwestern USA) and the first occurrences of pterosaurs outside the palaeotropics (that is, Pachagnathus, Yelaphomte and Arcticodactylus from Greenland and South America).
The stratigraphic and biogeographical distribution of lagerpetids and pterosaurs, and any other group, is the result of prominent interaction of biotic and abiotic factors. The palaeoclimatically informed phylogeographic model and spatially explicit ecological niche models we generated offer an explanation for the above-mentioned biogeographical patterns. First, our climate-based principal component analysis (PCA) shows that, in the Norian–Rhaetian, lagerpetids and pterosaurs had statistically distinct climate preferences (Fig. 3a and Extended Data Table 1). Lagerpetids occupied a broad climatic niche and are found to inhabit pronoucedly warmer (Fig. 3b and Extended Data Table 1) and drier (Fig. 3c and Extended Data Table 1) localities than those yielding Triassic pterosaurs. The occurrence of lagerpetids under varied climatic conditions suggests a remarkable ecological versatility, which may have enabled their more continuous distribution across different latitudes (Fig. 4). By contrast, early pterosaurs exhibit a patchier distribution, particularly in terms of mean annual and seasonal temperatures (Fig. 3d,e). Pterosaur distribution may indicate a narrower, more specialized climatic preference and/or that these suitable conditions were initially limited to equatorial areas. Nevertheless, suitable conditions for pterosaurs are found globally, although in discontinuous patches, and less frequently in continental interiors, in the late Norian–Rhaetian (Fig. 4b,d), in accordance with the pattern of quantified latitudinal dispersal and increased geographical spread in the fossil record (Fig. 1). Another possibility is that, during this early stage of pterosaur evolutionary history, perhaps owing to initially constrained flight capabilities and/or partial confinement to arboreal environments1 (for example, to protect from predation and thermal stress or abundant food supply), their biogeography was constrained by the distribution of the canopy habitat they inhabited. As more suitable areas became available by the Norian (Fig. 4), pterosaurs may have dispersed latitudinally, benefiting from potentially increased suitable habitats. However, their climatic preferences might have resulted in a sparse and locally restricted distribution, a possibility confirmed by their modelled, patchily distributed suitable areas. This aspect could potentially be attributed to their limited capabilities for long-distance aerial dispersal and/or a stage that was still partially confined to arboreal environments. This unique ecology could account for the relatively high dispersal inertia observed in pterosaur lineages (Fig. 2a) and more pronounced latitudinal dispersion towards the latest Triassic (Fig. 2b,c) (that is, endemic dispersion within climatic blocks). Our combined analyses suggest that changes in the distribution and extension of climatically suitable areas throughout the Triassic are congruent with latitudinal dispersal patterns, demonstrating limited geographical overlap between lagerpetids and pterosaurs. This may exemplify the intricate interplay between climatic conditions and habitat, as well as the development of diverse pterosauromorph ecological niches, that probably acted on the spatiotemporal distribution of this clade. Paradoxically, pterosaurs survived the end-Triassic mass extinction but lagerpetids did not, despite pterosaurs occupying a patchier, narrower climatic niche and having a more restricted geographic distribution than lagerpetids before this event. This suggests that other factors such as the ability to fly might have played important roles in the differential survivorship of these two clades during this extinction.
On the origin of pterosaurs
Historically, the evolution of powered flight in vertebrates (that is, pterosaurs and bats43,44) has resulted in a geologically instantaneous and geographically widespread distribution; however, until recently, a lack of early pterosaur fossils prevented any assessment into whether the evolution of a volant vertebrate leads to a predictable biogeographic outcome. Now, efforts to understand the timing and place of pterosaur origin have increased in concert with recent discoveries. A literal reading of the Triassic pterosaurs record suggests that the group originated at low latitudes in northern Pangaea. This hypothesis is supported both by the early-diverging phylogenetic positions of European taxa8 and by the oldest occurrence of indeterminate specimens of this group in the early–middle part of the Norian in the southwestern USA. However, it is worth noting that, while there is probably a genuine biogeographical signal in these observations, they could also reflect severe sampling biases4,5,6. We can now explore this issue by combining the results of our analyses to provide a potential model that incorporates the biogeographical and climatic circumstances around this event. However, we must first address the absence of Middle Triassic–early Late Triassic pterosaurs from the fossil record.
The most recent divergence time estimates based on pterosauromorph phylogenetic relationships constrain pterosaur origin to a 10–15-million-year interval that extends across the Ladinian–early Norian2. However, while an Early Triassic or Middle Triassic origin for pterosaurs cannot be excluded, no body or trace fossils are known before the Norian1,8,12. Most of the earliest pterosaurs have been found in black shale lithologies in mid-latitudes from the middle Norian Alpine arch (that is, Italy, Austria and Germany). Similar facies are found in the same geographical area throughout the Anisian–Carnian interval, but despite the high potential for exceptional preservation at these localities, pre-Norian pterosaurs have yet to be found (or are currently unrecognized) from well-sampled Middle Triassic and early Late Triassic Lagerstätten not only in this region (for example, Monte San Giorgio, late Anisian–late Ladinian, Italy and Switzerland; upper Buntsandstein, Muschelkalk and lower Keuper, Anisian–Carnian, in Germany and equivalent beds in central Europe; Lunz, early Carnian, Austria) but also worldwide (Yangjuan, Anisian, China; Digxiao, Ladinian, China; Xinpu, Carnian, China; Raibl, early Carnian, Italy; Madygan, Ladinian–Carnian, Kyrgyzstan)9,12, even after decades of extensive sampling. Similarly, the absence of pterosaurs remains unexplained in the lower part of the Newark Supergroup in North America (that is, Lockatong Formation and Cow Branch Formation, lower Norian), which has yielded the fragile remains of gliding reptiles such as Icarosaurus and Mecistotrachelos, among others45,46.
Ecological niche modelling offers a potential explanation for pterosaur absences in these localities. Our analyses suggest that many of these areas were climatically unsuitable for pterosaurs before the Norian (Fig. 4). For instance, the northern margin of the Tethys Ocean was climatically poorly suitable for pterosaurs until the late Carnian (Fig. 4). This could support the idea that pterosaurs originated shortly before their first appearance in the fossil record and/or that pterosaurs originated earlier, and elsewhere, but did not have a global distribution until later in the Norian1,12. However, while our model does not necessarily support a pre-Norian origin of the group in the Tethyan region, it provides an additional explanation for their earlier absence from this area and time interval. Specifically, our results suggest that there is potential for pre-Norian and early-Norian pterosaurs to be found elsewhere, in areas identified as habitable in our palaeoclimatically informed suitability models (Fig. 4 and Extended Data Fig. 3). Our niche models suggest that it might have been possible for pterosaurs to occur in the late Middle Triassic (Anisian–Ladinian) deposits of the southwest USA (upper Moenkopi Formation of Arizona and New Mexico), Morocco (members T3–T4 of the Timezgadiouine Formation, Argana Basin), India (Yerrapalli Formation), China (Xingyi fauna), Tanzania (Lifua Member of the Manda Beds), Brazil (lower Santa Maria Supersequence) and potentially southern and central Europe (for example, Dont Formation, Italy; Vellberg Formation, Germany). Suitable areas during the Carnian include the southwestern USA (for example, lower Otischalkian beds in the Dockum Group of Texas and New Mexico), Brazil (middle Santa Maria Formation), Argentina (Chañares Formation and lower Ischigualasto Formation), India (Tiki Formation and Maleri Formation), Madagascar (Isalo II/Makay Formation), continental Europe (Krasiejów locality, Poland), the lower Norian beds of the southwestern USA (upper Otischalkian and lowermost Adamanian beds of the Chinle Formation and Dockum Group, Arizona, Texas and New Mexico) and the basins of the Newark Supergroup, among others. In addition to these areas, the South China block shows high climatic suitability throughout the Anisian–early Norian. The upper portion of the Anisian–Carnian Guanling Formation (Panxian Fauna, Guanling Biota) also has high climatic suitability to yield pterosaur fossils, although the fossil record of its marine fauna shows limited terrestrial influence.
These results carry the caveat of known biases in the pterosaur fossil record4,5,6,9 (Supplementary Information) but provide testable hypotheses for future targeted fieldwork. While these formations were deposited in areas of high climatic suitability for pterosaurs, their geological characteristics might not have been suitable for preserving their fossils. As noted previously, many Triassic pterosaurs are preferentially preserved in black shale units1,12, although partial skeletons and individual bones have been more commonly recovered from fluvial, alluvial and desert depositional environments more recently20,38,41. Accordingly, among the above-mentioned formations, we particularly emphasize target lithologies representative of palaeoenvironments with known high preservation potential, particularly microvertebrate bonebeds, as these facies have the highest chance to preserve the small, fragile body fossils of Triassic pterosaurs. Articulated or associated skeletons comprise nearly all Triassic pterosaur occurrences, suggesting that their record in deposits where bones are recovered as disassociated and isolated elements (for example, microvertebrate bonebeds) may be underrepresented due to a lack of recognizable diagnostic features in collected skeletal elements from these settings.
Conclusions
Our study examines the historical palaeobiogeography of pterosaurs and their close relatives, lagerpetids, which together comprise pterosauromorphs. We identify climatic limits shaping the distribution of Triassic pterosauromorphs. Although multiple factors probably contributed to the distribution of these groups, the broad climatic adaptability and wide latitudinal spread of lagerpetids suggest ecological versatility, which allowed members of this group to thrive in localities with considerably warmer conditions than those yielding Triassic pterosaur remains. By contrast, early pterosaurs had a more constrained distribution in terms of mean annual and seasonal temperatures, favouring wetter areas that became more common after the Carnian, enabling their spread during the latest Triassic.
Our study independently aligns with the hypothesis suggesting that pterosaurs became widespread soon after their likely origin in the Middle Triassic–Late Triassic2. Furthermore, the modelled Carnian–early Norian pterosaur climatic niche would probably be found in temperate and wet conditions typical of tropical, near-coastal low-to-middle-latitude depositional environments. In emphasizing the complex interplay between climatic conditions, habitat and the development of diverse ecological niches, our study contributes valuable insights into the evolution and distribution of pterosaurs and their close relatives.
Methods
Palaeobiogeography
To explore the phylo-palaeobiogeography of Triassic pterosauromorphs, we implemented complementary analyses based on two recently published studies to (1) assess the degree to which pterosauromorphs dispersed across climatic barriers compared with other avemetatarsalian clades (following ref. 14) and (2) quantify the latitudinal dispersion of lagerpetids and pterosaurs alongside that of other avemetatarsalians (following ref. 11). To better compare the results of these analyses, which drew on different datasets, we assembled and tip-dated an avemetatarsalian supertree (Extended Data Fig. 1). This grafts the dinosaur tree topology recovered by ref. 14 into a randomly selected most parsimonious tree of Archosauromorpha from ref. 11. We then manually incorporated the dinosaurian and silesaurid taxa present in the latter dataset but not the former. This strategy allowed us to retain the entire pterosauromorph sample included in ref. 11, which is the most comprehensive available for this group. Each taxon in this newly assembled dataset was then scored for two different sets of geographical areas (see below). This was deemed necessary because the two above-mentioned studies were designed using distinct geographical frameworks (see details below). Overall, these operations improved the sampling of the selected avemetatarsalian groups and allowed more meaningful comparisons between the two sets of biogeographical analyses. Our analyses cover the Ladinian–Rhaetian interval, but the taxonomic sample includes Jurassic species, enabling consideration of all potential dispersal events nested in the Triassic.
Griffin et al.14 demonstrated the limited impact of changing archosaur topology in their results. However, their tree did not include a broad sample of lagerpetids and pterosaurs, so here we focus on the impact that different pterosauromorph tree topologies could have on our results. In Pterosauromorpha, the relationships within Pterosauria are relatively well established, so the topology here adopted corresponds to the strict consensus from ref. 11 and derivative analyses. Conversely, the relationships within Lagerpetidae are not well resolved, and it is not possible to produce a fully resolved strict consensus topology. However, for the purpose of our biogeographical analyses, this does not matter because the alternance of ‘Laurasian’ and ‘Gondwanan’ areas of origin at the tips is broadly maintained: a South American taxon (that is, Faxinalipterus) is consistently found at the earliest diverging branch, followed by Scleromochlus, from Europe. Further up the tree, a minority of North America taxa (that is, Dromomeron romeri and Dromomeron gregorii) are found nested within a majority of South American species. Our choice of using a randomly selected tree following ref. 14 is thus justified because the patterns of geographical areas at the tips of the tree are broadly stable and the patterns of potential of latitudinal dispersion and accumulated latitudinal dispersion are not affected by the taxon order of the tips (Supplementary Information).
Potential of latitudinal dispersion
First, we implemented an event-based quantitative analysis that aimed to assess the potential for pterosauromorph latitudinal dispersal during the waxing and waning of climatic barriers, following the methodology of ref. 14. Griffin et al.14 originally developed this method to test “whether the early phylogenetic history of dinosaurs retains a signal for restricted dispersal” (p. 317). Here, we applied the protocol to pterosauromorphs, which were sparsely represented in the latter study. We limit our interpretation of the pterosauromorph curve to the Ladinian–Carnian portion of the whole interval, where the signal is driven primarily by lagerpetids, because we cannot be sure to what extent our model is applicable to volant taxa, as pterosaurs affect the Carnian–Rhaetian portion of the results.
We implemented this analysis following the methodology of Griffin et al.14. First, we scored each taxon in the newly assembled dataset for one of five geographical regions: eastern Laurasia, western Laurasia, equatorial belt, northern Gondwana or southern Gondwana. These regions were selected because of the biogeographical importance of their boundaries throughout the Ladinian–Rhaetian (our interval of interest), namely, an arid belt in low-latitude southern Pangaea47 and, less relevant to the time interval of our study, the Hispanic Corridor and Viking Strait48,49. Second, we extended the lower boundary of the analyses to include the Ladinian stage to include the complete stratigraphic range of lagerpetids. We then implemented a dispersal–extinction–cladogenesis (DEC) model on the time-calibrated phylogeny under maximum likelihood. To simulate the waxing and waning of arid climatic barriers to dispersal, we adopted distinct rate matrices of dispersal across selected geographical areas and tested their occurrence across the whole interval. The Δ-likelihood is a measure of how strongly the dispersal pattern is supported at any time: in other words, a more negative Δ indicates dispersal/waning of climatic barriers. The Δ measures how much dispersal is occurring by imposing a penalty when the climate barrier is crossed. Thus, the Δ will be more negative when more organisms (lineages) are dispersing when barriers are present. Conversely, a Δ closer to zero indicates strong barriers to dispersal, or that the crossing of barriers was sporadic and largely followed by cladogenesis. Overall, this model uses the phylogeny of different avemetatarsalian groups to test whether, and when, climatic barriers affected the dispersal of each group, particularly lagerpetids, during the Ladinian–Rhaetian.
Accumulated latitudinal dispersion
We also quantified the latitudinal dispersal of each avemetatarsalian group, following the methodology of Müller et al.11. Differing from that study, we decided against pruning the lagerpetids Kongonaphon kely and PVSJ 883 to enhance completeness. The stratigraphic and palaeobiogeographical occurrences of these taxa are of paramount importance for our analyses, and we believe that the benefits of their inclusion outweigh the uncertainties in their phylogenetic positions. Specifically, Kongonaphon kely is not only the stratigraphically oldest lagerpetid but also the only member of this clade from this region of Gondwana (Madagascar, representing the Indian subcontinent + Madagascar continental area). Similarly, we retained the unnamed taxon PVSJ 883, from the late Carnian of Argentina10,16, because its occurrence suggests a dispersal event that would otherwise be ignored.
To run the analyses, we used the same tip-dated supertree of Avemetatarsalia as discussed above. This time, each taxon in the dataset was assigned to one of eight discrete geographic areas (that is, western North America, eastern North America, Brazil, Argentina, Europe + Russia, South Africa, and Indian subcontinent + Madagascar) among the ten determined by Button et al.50. The quantification of dispersal events relies on reconstructing ancestral states at internal nodes. This was achieved using the R package BioGeoBEARS51 v.1.1.2 in R v.4.3.1 (ref. 52), using different likelihood-based palaeobiogeographic models: DEC, dispersal vicariance analysis likelihood (LIKEDIVA) and BAYAREALIKE. Only the simple DEC model was used for subsequent calculations to avoid long-distance palaeolatitudinal jumps in the latitudinal dispersion calculations14 following Müller et al.11, and for meaningful comparison with the first analysis that is also based on DEC14 for the same reason. However, we notice that, in our comparisons of biogeographical models, ‘+j’ models are better supported (Supplementary Table 1). This may suggest a more complex biogeographic history in which (aerial?) dispersal rather than vicariance may have been the dominating phenomenon (expected in flying animals) in early pterosaur macroevolution.
Because the main barriers preventing dispersion in the Middle to Late Triassic were arid latitudinal belts at mid-to-low latitudes47, we quantified the accumulated amount of latitudinal dispersal of lagerpetids and pterosaurs separately and compared them with those of other avemetatarsalian clades (silesaurids and dinosaurs).
First, we counted the number of dispersal events across the tree by identifying those branches with different geographical values at their nodes (or tips). The latitudes of well-known avemetatarsalian-bearing localities were adopted as proxies for the broader geographic area and used to calculate the latitudinal extent of dispersion events. The localities we used are the same as those in Müller et al.11: Otis Chalk Quarries (western North America, palaeolatitude 6.9° N); New Haven County (eastern North America, 16.2° N); Buriol site (Brazil, palaeolatitude 39.7° S); Quebrada del Puma site (Argentina, palaeolatitude 40.6° S); Lombardia (Europe + Russia, palaeolatitude 27.1° N); Free State (South Africa, 42.8° S); and Manda (Indian subcontinent, palaeolatitude 53.7° S). Unlike ref. 11, we did not discard uncertain nodes but instead averaged the latitudinal values of different areas when an overwhelming signal was not computed. Only events that started (but did not necessarily end) in the Triassic were included in the final analyses.
The total amount of latitudinal dispersion (measured in degrees) for each clade was then calculated and plotted across the Anisian–Rhaetian through 1-million-year-long bins (Fig. 2b) and subsequently averaged by the number of dispersal events in each interval (Fig. 2c). These calculations were done using a slight modification of the R codes provided by Müller et al.11 (see ‘Code availability’ section).
Palaeoclimate niche occupation
To explore the palaeoclimatic niches occupied by pterosauromorph lineages, we compiled a locality-based dataset of all Triassic lagerpetid and pterosaur occurrences. This integrated a literature search with unpublished fieldwork data (20+ years in the southwestern USA in Arizona, Texas and New Mexico)3,11,17,20,28,29,30,31,32,33,34,35,36,37,38,40,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74 (see ‘Dataset_pterosauromorphs_R1.xls’ in Supplementary Data 1). Unique occurrence data were recorded for each specimen in our dataset. Unnamed and/or undescribed specimens were examined in person and included to increase the data available for our palaeoclimate niche analyses. This sampling strategy means that the datasets for the palaeobiogeographic and climate analyses are independent (that is, the first relies on phylogeny and the second on occurrence data), but all taxa present in the former are also included in the latter. This strategy recognizes the value of including undescribed or indeterminate lagerpetid and pterosaur specimens, which provide fundamental information on the distributions of their respective clades, even if they cannot be included in phylogenetic analyses. The lagerpetids included in the palaeoclimate niche analyses but excluded from the phylogenetic framework (and, hence, the palaeobiogeographic analyses) are: the indeterminate lagerpetids (NMMMNH P-80469, PEFO 44476 and PEFO 50545) from, respectively, NMMNH L-149 in the Los Esteros Member of the Santa Rosa Formation of New Mexico (Otischalkian holochronozone, lower Norian)64, PFV 456 (Thunderstorm Ridge) in the Blue Mesa Member (Adamanian holochronozone, middle Norian)34 and PFV 215 (Zuni Well Mound) in the Petrified Forest Member (Revueltian holochronozone, upper Norian)34 of the Chinle Formation of Petrified Forest National Park and multiple representatives of the genus Dromomeron from 13 distinct localities in the southwestern USA30,31,34,59,60,61,62,67 (Fig. 1 and Supplementary Data 1). Similarly, the pterosaurs that are present in the palaeoclimate analysis but could not be included in the phylogenetic dataset are Arcticodactylus from Greenland38,40, which is currently under investigation68; three unnamed pterosaurs from the southwestern USA, including an undescribed pterosaur (tentatively referred to as Eudimorphodon sp.) from the ‘Kalgary localities’ (approximately upper Carnian–lower Norian) of the Tecovas Formation of the Dockum Group3,69 and fig. 9.5 in ref. 70, two undescribed pterosaurs from the Chinle Formation of Petrified Forest National Park, one (PEFO 45782) from PFV 456 (Thunderstorm Ridge) in the Blue Mesa Member (Adamanian holochronozone, middle Norian, ~220 Mya (refs. 73,74)); and an undescribed pterosaur (PEFO 53384) from the Owl Rock Member (Apachean holochronozone, late Norian)71. Finally, MCSNB 8950 is an unnamed taxon from the Argilliti di Riva di Solto Formation of northern Italy (upper Norian), which was previously referred to Eudimorphodon ranzi72 but is now thought to be an unnamed new genus12.
However, because the resolution of the general circulation (palaeoclimatic) models is 1° × 1° (~111 km2) (see below), we consider only one locality if more than one specimen was found in a radius smaller than 111 km, unless they yielded different taxa (for example, the pterosaur localities in northern Italy). The occurrence of taxa with uncertain stratigraphic ranges covering multiple time bins was considered present in each of the relevant bins. This resulted in a summarized dataset of 54 occurrences (24 pterosaur and 30 lagerpetid entries; Supplementary Data 1).
Palaeoclimate reconstructions
To explore whether palaeoclimatic conditions influenced the biogeographic distributions of pterosaurs and lagerpetids, we integrated occurrence data for these groups with outputs from a general circulation (palaeoclimate) model. The values extracted from these models show a good match with the estimates from geochemical, sedimentological and biological proxies at the basin level and broader scales (for example, see refs. 75,76 for a detailed study of the palaeoclimate of the Chañares–Los Rastros–Ischigualasto Triassic succession). The accuracy and evaluation of general circulation (palaeoclimate) model outputs are further discussed in the Supplementary Information. Palaeoclimate model simulations used a recent version of the coupled Atmosphere–Ocean General Circulation Model and HadCM3L77 (specifically HadCM3L-M2.1aD), following the nomenclature in the work of Malanoski et al.78), including the modifications described by Judd et al.79. The model has a resolution of 3.75° longitude × 2.5° latitude in the atmosphere and ocean, with 19 hybrid levels in the atmosphere and 20 vertical levels in the ocean with equations solved on the Arakawa B-grid. Atmospheric subgrid scale processes such as convection are parameterized as they cannot be resolved at the resolution of the model. The ocean model is based on the model of Valdes et al.80 and is a full primitive equation, three-dimensional model of the ocean. A second-order numerical scheme is used along with centred advection to remove nonlinear instabilities. Flux adjustments (such as artificial heat and salinity adjustments in the ocean model to prevent the model drifting to unrealistic values) are not required in this model81, which is crucial for long palaeoclimate simulations. Sea ice is calculated on a zero-layer model with partial sea ice coverage possible, with a consistent salinity assumed for ice. Because geological data recording land surface vegetation for Triassic stages are uncertain and globally sparse, we use a version of the model that includes the dynamical vegetation model TRIFFID (Top-Down Representation of Interactive Foliage and Flora Including Dynamics) and land surface scheme MOSES 2.1a (ref. 82). TRIFFID predicts the distribution and properties of global vegetation based on plant functional types (PFTs), in the form of fractional coverage (and, thus, PFT coexistence) within a grid cell, and is, in turn, based on competition equations based on the climate tolerance of five PFTs. The HadCM3L model demonstrates high performance at reproducing the modern-day climate79 and has been used for an array of pre-Quaternary palaeoclimate experiments79,83,84. Palaeoclimate experiments typically require on the order of hundreds of years to reach a near-surface quasi-equilibrium state (but many thousands of years for the deep ocean79,80) as well as true climate equilibrium. Relatively low-resolution global climate models such as HadCM3L are relatively computationally expensive, allowing near-fully equilibrated simulations of climate to be undertaken that would not be possible with higher-resolution, more complex models85.
Seven model simulations with Ladinian (239.54 Ma), ‘early’ Carnian (233.6 Ma), ‘late’ Carnian (232 Ma), ‘early’ Norian (227 Ma), ‘middle’ Norian (222.4 Ma), ‘late’ Norian (217.8 Ma) and Rhaetian (204.9 Ma) were carried out using stage-specific boundary conditions (topography, bathymetry, solar luminosity, continental ice and partial pressure of CO2 (pCO2)). Stage-specific realistic carbon dioxide concentrations were chosen on the basis of proxy-CO2 (Ladinian, 1,034 ppm; ‘early’ Carnian, 1,492 ppm; ‘late’ Carnian, 1,614 ppm; ‘early’ Norian, 2,059 ppm; ‘middle’ Norian, 1,810 ppm; ‘late’ Norian, 1,481 ppm; and Rhaetian, 1,503 ppm) reconstructions from ref. 86, and the solar constant was based on ref. 87. The orography and bathymetry were derived from palaeogeographic digital elevation models, produced by ref. 88 as part of the PALEOMAP project (see ref. 79 for more details). Each stage-specific digital elevation model is interpolated from a 1° × 1° grid onto the model 3.75° × 2.5° grid. Similarly, land ice is also transformed onto the model grid assuming a simple parabolic shape to estimate the ice sheet height (m). Surface soil conditions were set at a uniform medium loam everywhere because stage-specific soil parameters during the Triassic are not globally known. All other boundary conditions (such as orbital parameters, aerosol concentrations and so on) are held constant at preindustrial values. The simulations were carried out for a total of over 10,000 years. By the end of the simulations, (1) the globally and volume-integrated annual mean ocean temperature trend is less than 1 °C per 1,000 years; (2) trends in surface air temperature are less than 0.3 °C per 1,000 years; and (3) net energy balance at the top of the atmosphere, averaged over a 100-year period at the end of the simulation, is less than 0.25 W m−2. Climatological means were produced from the last 100 years of each simulation. All these simulations are identical to the ‘Scotese07’ simulations described in ref. 78 and the ‘model 2’ simulations of ref. 79. The climate predicted by the model is compared with proxy indicators of climate in the Supplementary Information. The outputs from the simulations are available at https://www.paleo.bristol.ac.uk/ummodel/scripts/html_bridge/scotese_07.html.
Palaeoclimatic niche space
To explore the climatic conditions occupied by pterosauromorphs during the Late Triassic, multivariate statistical tests were used and summary statistic plots were constructed. This approach is commonly used in modern ecology89,90,91,92,93 and, in the past decade, has become more common in studies pertaining to the fossil record of both invertebrate94,95,96,97,98,99 and vertebrate groups15,16,73,100. Accordingly, each taxon was assigned climate variables based on the mean values for their stratigraphic age and geographic locations (namely, mean annual temperature (MAT), MAP, seasonal variation in temperature and seasonal variation in precipitation; Fig. 3 and Supplementary Table 2). Taxa spanning two geological stages (n = 8) were assigned the mean of the variables for both stages and assigned single, averaged, stratigraphic occurrence. To quantify ‘palaeoclimatic niche space’ for each taxon, we followed the procedure outlined in ref. 16. Obtaining information on species’ fundamental niches from the fossil record is challenging: therefore, the term ‘palaeoclimatic niche’ as used here refers to an approximation of the realized climatic niche of the fossil taxa (that is, the set of climatic conditions occupied by a taxon16). A PCA was performed using the prcomp() function in R v.4.4.2 (ref. 52), which included the scaling argument so that variables were scaled to have unit variance before the analysis took place. A non-parametric MANOVA was performed to statistically compare the distribution of the two groups using the R package RVAideMemoire89. Raincloud plots displaying both box plots (depicting the distributions of the palaeoclimate data within the taxonomic groups) and frequency distributions were constructed to examine the range of individual palaeoclimatic conditions occupied by both groups. To statistically compare the distributions between both groups, pairwise comparisons were performed in R using Wilcoxon rank-sum (Mann–Whitney) tests. These specific statistical tests were chosen because the palaeoclimate variable data do not conform to the assumptions of a normal distribution, which was determined through probability plots (for example, quartile–quartile plots) and Shapiro–Wilk tests.
Habitat suitability modelling
To assess the climatic suitability of these taxa, we implemented the DOMAIN algorithm100 through the R package ‘dismo’101, which is an ecological niche modelling tool using the Gower distance metric to assess climatic suitability. The DOMAIN algorithm quantifies the disparity between the climatic conditions of map pixels and the nearest species observation within the n-dimensional environmental space, diverging from geographic proximity. Although this model is generally classified as a coarse niche modelling technique102,103,104,105, it offers the advantage of straightforward implementation and minimal assumptions, which are necessary due to the paucity of the pterosauromorph fossil record. Given the coarse spatial and temporal resolution of our dataset, coupled with our exclusive focus on producing suitability maps based solely on climate, and the low number of occurrences in our datasets, we favoured this simplistic modelling approach over more recently introduced ecological niche and habitat suitability modelling techniques, including those previously used by our team in prior works101,102,103,104,105, where the associated limitations and assumptions are also addressed. We furthermore calibrated the model on a compound occurrence dataset that maximizes the geographic spread for each of the subclades, centring on the best sampled interval (the middle–late Norian, 218 Ma) and including palaeobiogeographic outliers from the Carnian (Lagerpetidae) and throughout the full duration of the Norian for pterosaurs, projecting these models into more refined climatic simulations (eight time slices from the Ladinian to the Rhaetian). MAT (°C) and MAP (mm per year) are the variables used for the habitat suitability modelling.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All datasets can be found alongside their respective R codes via Figshare at https://figshare.com/s/742a30f30deb4aefb60a (ref. 106), Supplementary Data 1; https://figshare.com/s/7ac05f57d35b3810a2c7 (ref. 107), potential for latitudinal dispersion; https://figshare.com/s/8cabc46601eb64f9b9f0 (ref. 108), accumulated latitudinal dispersion; https://figshare.com/s/6254a692186e79fa9060 (ref. 109), palaeoclimate niche occupation; and https://figshare.com/s/6600233d24db8f1d986e (ref. 110), habitat suitability modelling., climate modelling data repository (https://www.paleo.bristol.ac.uk/ummodel/scripts/papers/Foffa_etal_2025.html). Source data are provided with this paper.
Code availability
All datasets can be found alongside their respective R codes via Figshare at https://figshare.com/s/7ac05f57d35b3810a2c7 (ref. 107), potential for latitudinal dispersion; https://figshare.com/s/8cabc46601eb64f9b9f0 (ref. 108), accumulated latitudinal dispersion; https://figshare.com/s/6254a692186e79fa9060 (ref. 109), palaeoclimate niche occupation; and https://figshare.com/s/6600233d24db8f1d986e (ref. 110), habitat suitability modelling.
References
Baron, M. G. The origin of pterosaurs. Earth-Sci. Rev. 221, 103777 (2021).
Ezcurra, M. D. et al. Enigmatic dinosaur precursors bridge the gap to the origin of Pterosauria. Nature 588, 445–449 (2020).
Barrett, P. M. et al. Pterosaur distribution in time and space: an atlas. Zitteliana 28, 61–107 (2008).
Butler, R. J. et al. Estimating the effects of sampling biases on pterosaur diversity patterns: implications for hypotheses of bird/pterosaur competitive replacement. Paleobiology 35, 432–446 (2009).
Butler, R. J., Benson, R. B. J. & Barrett, P. M. Pterosaur diversity: untangling the influence of sampling biases, Lagerstätten, and genuine biodiversity signals. Palaeogeogr. Palaeoclimatol. Palaeoecol. 372, 78–87 (2013).
Butler, R. J. et al. How do geological sampling biases affect studies of morphological evolution in deep time? A case study of pterosaur (Reptilia: Archosauria) disparity. Evolution 66, 147–162 (2012).
Langer, M. C. et al. in Anatomy, Phylogeny and Palaeobiology of Early Archosaurs and their Kin (eds Nesbitt, S. J. et al.) 156–186 (Geological Society London, 2013).
Upchurch, P. et al. An analysis of pterosaurian biogeography: implications for the evolutionary history and fossil record quality of the first flying vertebrates. Hist. Biol. 27, 697–717 (2015).
Dean, C. D., Mannion, P. D. & Butler, R. J. Preservational bias controls the fossil record of pterosaurs. Palaeontology 59, 225–247 (2016).
Müller, R. T., Langer, M. C. & Dias-Da-Silva, S. Ingroup relationships of Lagerpetidae (Avemetatarsalia: Dinosauromorpha): a further phylogenetic investigation on the understanding of dinosaur relatives. Zootaxa 4392, 149–158 (2018).
Müller, R. T. et al. New reptile shows dinosaurs and pterosaurs evolved among diverse precursors. Nature 620, 589–594 (2023).
Dalla Vecchia, F. M. in Anatomy, Phylogeny and Palaeobiology of Early Archosaurs and their Kin (eds Nesbitt, S. J. et al.) 119–155 (Geological Society London, 2013).
Baron, M. G. Testing pterosaur ingroup relationships through broader sampling of avemetatarsalian taxa and characters and a range of phylogenetic analysis techniques. PeerJ 8, e9604 (2020).
Griffin, C. T. et al. Africa’s oldest dinosaurs reveal early suppression of dinosaur distribution. Nature 609, 313–319 (2022).
Dunne, E. M. et al. Climatic drivers of latitudinal variation in Late Triassic tetrapod diversity. Palaeontology 64, 101–117 (2021).
Dunne, E. M. et al. Climatic controls on the ecological ascendancy of dinosaurs. Curr. Biol. 33, 206–214 (2023).
Kammerer, C. F. et al. A tiny ornithodiran archosaur from the Triassic of Madagascar and the role of miniaturization in dinosaur and pterosaur ancestry. Proc. Natl Acad. Sci. USA 117, 17932–17936 (2020).
Foffa, D. et al. Scleromochlus and the early evolution of Pterosauromorpha. Nature 610, 313–318 (2022).
Kellner, A. W. A. et al. Reassessment of Faxinalipterus minimus, a purported Triassic pterosaur from southern Brazil with the description of a new taxon. PeerJ 10, e13276 (2022).
Martínez, R. N. et al. The dawn of the flying reptiles: first Triassic record in the southern hemisphere. Pap. Palaeontol. 8, e1424 (2022).
Woodward, A. S. On a new dinosaurian reptile (Scleromochlus taylori, gen. et sp. nov.) from the Trias of Lossiemouth, Elgin. Quart. J. Geol. Soc. Lond. 63, 1–4 (1907).
Benton, M. J. Scleromochlus taylori and the origin of dinosaurs and pterosaurs. Philos. Trans. R. Soc. B 354, 1423–1446 (1999).
Bennett, C. S. Reassessment of the Triassic archosauriform Scleromochlus taylori neither runner nor biped, but hopper. PeerJ 8, e8418 (2020).
Foffa, D. et al. The osteology of the Late Triassic reptile Scleromochlus taylori from μCT data. Anat. Rec. https://doi.org/10.1002/ar.25335 (2023).
Romer, A. S. The Chanares (Argentina) Triassic reptile fauna X. Two new but incompletely known long-limbed pseudosuchians. Brevoria 378, 1–10 (1971).
Sereno, P. C. & Arcucci, A. B. Dinosaurian precursors from the Middle Triassic of Argentina: Lagerpeton chanarensis. J. Vertebr. Paleontol. 13, 385–399 (1994).
Langer, M. C., Ramezani, J. & Da Rosa, Á. A. S. U–Pb age constraints on dinosaur rise from south Brazil. Gondwana Res. 57, 133–140 (2018).
Garcia, M. S. et al. The oldest known co-occurrence of dinosaurs and their closest relatives: a new lagerpetid from a Carnian (Upper Triassic) bed of Brazil with implications for dinosauromorph biostratigraphy, early diversification and biogeography. J. S. Am. Earth. Sci. 91, 302–319 (2019).
Cabreira, S. F. et al. A unique Late Triassic dinosauromorph assemblage reveals dinosaur ancestral anatomy and diet. Curr. Biol. 26, 3090–3095 (2016).
Irmis, R. B. et al. A Late Triassic dinosauromorph assemblage from New Mexico and the rise of dinosaurs. Science 317, 358–361 (2009).
Nesbitt, S. J. et al. Hindlimb osteology and distribution of basal dinosauromorphs from the Late Triassic of North America. J. Vertebr. Paleontol. 29, 498–516 (2009).
Martínez, R. N. et al. Vertebrate succession in the Ischigualasto Formation. J. Vertebr. Paleontol. Mem. 32, 10–30 (2012).
Martínez, R. N. et al. A Norian lagerpetid dinosauromorph from the Quebrada del Barro Formation, northwestern Argentina. Ameghiniana 53, 1–13 (2016).
Marsh, A. D. & Parker, W. G. New dinosauromorph specimens from Petrified Forest National Park and a global biostratigraphic review of Triassic dinosauromorph body fossils. PaleoBios 37, 1–56 (2020).
Zambelli, R. Eudimorphodon ranzii gen. nov., sp. nov., uno pterosauro triassico. Rend. Sci. Inst. Lombardo B 107, 27–32 (1973).
Wild, R. Die Flugsaurier (Reptilia, Pterosauria) aus der Oberen Trias von Cene bei Bergamo, Italien. Boll. Soc. Paleontol. Ital. 17, 176–256 (1978).
Dalla Vecchia, F. M. Seazzadactylus venieri gen. et sp. nov., a new pterosaur (Diapsida: Pterosauria) from the Upper Triassic (Norian) of northeastern Italy. PeerJ 7, e7363 (2019).
Jenkins, F. A. Jr et al. A diminutive pterosaur (Pterosauria: Eudimorphodontidae) from the Greenlandic Triassic. Bull. Mus. Comp. Zool. 156, 151–170 (2001).
Andres, B., Clark, J. & Xu, X. The earliest pterodactyloid and the origin of the group. Curr. Biol. 24, 1011–1016 (2014).
Kellner, A. W. A. Comments on Triassic pterosaurs with discussion about ontogeny and description of new taxa. Acad. Bras. Cienc. 87, 669–689 (2015).
Britt, B. B. et al. Caelestiventus hanseni gen. et sp. nov. extends the desert-dwelling pterosaur record back 65 million years. Nat. Ecol. Evol. 2, 1386–1392 (2018).
Yu, Y., Zhang, C. & Xu, X. Complex macroevolution of pterosaurs. Curr. Biol. 33, 770–779 (2023).
Brown, E. E. et al. Quantifying the completeness of the bat fossil record. Palaeontology 62, 757–776 (2019).
Gunnell, G. F. & Simmons, N. B. Fossil evi-dence and the origin of bats. J. Mamm. Evol. 12, 209–246 (2005).
Colbert, E. H. A gliding reptile from the Triassic of New Jersey. Am. Mus. Novit. 2246, 1–23 (1966).
Fraser, N. C. et al. A new gliding tetrapod (Diapsida:? Archosauromorpha) from the Upper Triassic (Carnian) of Virginia. J. Vertebr. Paleontol. 27, 261–265 (2007).
Whiteside, J. H. et al. Extreme ecosystem instability suppressed tropical dinosaur dominance for 30 million years. Proc. Natl Acad. Sci. USA 112, 7909–7913 (2015).
Aberhan, M. Bivalve palaeobiogeography and the Hispanic Corridor: time of opening and effectiveness of a proto-Atlantic seaway. Palaeogeogr. Palaeoclimatol. Palaeoecol. 165, 375–394 (2001).
Schöllhorn, I. et al. Climate and environmental response to the break-up of Pangea during the Early Jurassic (Hettangian-Pliensbachian); the Dorset coast (UK) revisited. Glob. Planet. Change 185, 103096 (2020).
Button, D. J. et al. Mass extinctions drove increased global faunal cosmopolitanism on the supercontinent Pangaea. Nat. Commun. 8, 733 (2017).
Matzke, N. J. BioGeoBEARS: BioGeography with Bayesian (and likelihood) Evolutionary Analysis with R Scripts. version 1.1.1, published on GitHub on November 6, 2018. Zenodo https://doi.org/10.5281/zenodo.1478250 (2018).
R Core Team. R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, Vienna, Austria, 2021); https://www.R-project.org/
Dalla Vecchia, F. M. et al. A crested rhamphorhynchoid pterosaur from the Late Triassic of Austria. J. Vert. Paleontol. 22, 196–199 (2002).
Dalla Vecchia, F. M. The first Italian specimen of Austriadactylus cristatus (Diapsida, Pterosauria) from the Norian (Upper Triassic) of the Carnic Prealps. Riv. Ital. Paleontol. Stratigr. 115, 291–304 (2009).
Stecher, R. A new Triassic pterosaur from Switzerland (Central Austroalpine, Grisons), Raeticodactylus filisurensis gen. et sp. nov. Swiss J. Geosci. 101, 185–201 (2008).
Dalla Vecchia, F. M. Anatomy and systematics of the pterosaur Carniadactylus (gen. n.) rosenfeldi (Dalla Vecchia, 1995). Riv. Ital. Paleontol. Stratigr. 115, 159–188 (2009).
Arcucci, A. New materials and reinterpretation of Lagerpeton chanarensis Romer (Thecodontia, Lagerpetonidae nov.) from the Middle Triassic of La Rioja, Argentina. Ameghiniana 23, 233–242 (1986).
McCabe, M. B. & Nesbitt, S. J. The first pectoral and forelimb material assigned to the lagerpetid Lagerpeton chanarensis (Archosauria: Dinosauromorpha) from the upper portion of the Chañares Formation, Late Triassic. Palaeodiversity 14, 121–131 (2021).
Martz, J. W. et al. A taxonomic and biostratigraphic re-evaluation of the Post Quarry vertebrate assemblage from the Cooper Canyon Formation (Dockum Group, Upper Triassic) of southern Garza County, western Texas. Proc. R. Soc. Edinb. 103, 339–364 (2013).
Sarıgül, V. New basal dinosauromorph records from the Dockum Group of Texas, USA. Palaeontol. Electron 19.2.21A, 1–16 (2016).
Lessner, E. J. et al. New insights into Late Triassic dinosauromorph bearing assemblages from Texas using apomorphy-based identifications. PaleoBios 35, 1–41 (2018).
Martz, J. W. & Small, B. J. Non-dinosaurian dinosauromorphs from the Chinle Formation (Upper Triassic) of the Eagle Basin, northern Colorado: Dromomeron romeri (Lagerpetidae) and a new taxon, Kwanasaurus williamparkeri (Silesauridae). PeerJ 7, e7551 (2019).
Fröbisch, N. B. & Fröbisch, J. A new basal pterosaur genus from the upper Triassic of the Northern Calcareous Alps of Switzerland. Palaeontology 49, 1081–1090 (2006).
Beyl, A. et al. An Otischalkian dinosauromorph assemblage from the Los Esteros Member (Santa Rosa Formation) of New Mexico and its implications for biochronology and lagerpetid body size. J. Vert. Paleontol. 40, e1765788 (2020).
Garcia, M. S. et al. A new sympatric occurrence of lagerpetids (Pan-Aves, Pterosauromorpha) in the upper Triassic of southern Brazil. J. S. Am. Earth Sci. 140, 104897 (2024).
Dalla Vecchia, F. M. Comments on Triassic pterosaurs with a commentary on the “ontogenetic stages” of Kellner (2015) and the validity of Bergamodactylus wildi. Riv. It. Paleontol. Strat. 124, 317–341 (2018).
Marsh, A. D. A new record of Dromomeron romeri Irmis et al., 2007 (Lagerpetidae) from the Chinle Formation of Arizona, USA. PaleoBios 35, 1–8 (2018).
Fitch, A. J. et al. Origins of Pterosauria: hidden fossil fossil record and near extinction of the first and longest-lasting flying vertebrate lineage revealed through a new phylogenetic hypothesis. J. Vertebr. Paleontol. Program and Abstracts 144 (2022).
Andres, B. & Myers, T. S. Lone star pterosaurs. Earth Environ. Sci. Trans. R. Soc. Edinb. 103, 383–398 (2012).
Murry, P. A. in Vertebrate Paleontology of the Dockum Group, Western Texas and Eastern New Mexico. The Beginning of the Age of Dinosaurs (ed. Padian, K.) 109–137 (Cambridge Univ. Press, 1986).
Behrensmeyer, A. K. et al. Taphonomy and paleocommunity reconstruction of a pterosaur-bearing fossil assemblage in the Upper Triassic of Arizona: J. Vertebr. Paleontol., Program and Abstracts 60 (2019).
Wild, R. A new pterosaur (Reptilia, Pterosauria) from the Upper Triassic (Norian) of Friuli, Italy. Gortania 5, 45–62 (1984).
Ramezani, J. et al. High-precision U–Pb zircon geochronology of the Late Triassic Chinle Formation, Petrified Forest National Park (Arizona, USA): temporal constraints on the early evolution of dinosaurs. Bulletin 123, 2142–2159 (2011).
Ramezani, J., Fastovsky, D. E. & Bowring, S. A. Revised chronostratigraphy of the lower Chinle Formation strata in Arizona and New Mexico (USA): high-precision U–Pb geochronological constraints on the Late Triassic evolution of dinosaurs. Am. J. Sci. 314, 981–1008 (2014).
Mancuso, A. C., Horn, B. L. D., Benavente, C. A., Schultz, C. L. & Irmis, R. B. The paleoclimatic context for South American Triassic vertebrate evolution. J. S. Am. Earth. Sci. 110, 103321 (2021).
Mancuso, A. C. et al. Paleoenvironmental and biotic changes in the Late Triassic of Argentina: testing hypotheses of abiotic forcing at the basin scale. Front. Earth Sci. 10, 883788 (2022).
Valdes, P. J. et al. The BRIDGE HadCM3 family of climate models: HadCM3@Bristol v1.0. Geosci. Model Dev. 10, 3715–3743 (2017).
Judd, E. J. et al. A 485-million-year history of Earth’s surface temperature. Science 385, eadk3705 (2024).
Malanoski, C. M., Farnsworth, A., Lunt, D. J., Valdes, P. J. & Saupe, E. E. Climate change is an important predictor of extinction risk on macroevolutionary timescales. Science 383, 1130–1134 (2024).
Valdes, P. J., Scotese, C. R. & Lunt, D. J. Deep ocean temperatures through time. Clim 17, 1483–1506 (2021).
Cox, P. A primitive equation, 3-dimensional model of the ocean, GFDL Ocean Group Technical Report No. 1. Available from Geophysical Fluid Dynamics Laboratory. (1984).
Collins, M., Tett, S. F. B. & Cooper, C. The internal climate variability of HadCM3, a version of the Hadley Centre coupled model without flux adjustments. Clim. Dyn. 17, 61–81 (2001).
Cox, P. et al. The impact of new land surface physics on the GCM simulation of climate and climate sensitivity. Clim. Dyn. 15, 183–203 (1999).
Farnsworth, A. et al. Climate sensitivity on geological timescales controlled by nonlinear feedbacks and ocean circulation. Geophy. Res. Lett. 46, 9880–9889 (2019). (2019a).
Farnsworth, A. et al. Pancost, Past East Asian monsoon evolution controlled by paleogeography, not CO2. Sci. Adv. 5, eaax1697 (2019b).
Foster, G. L., Royer, D. L. & Lunt, D. J. Future climate forcing potentially without precedent in the last 420 million years. Nat. Commun. 8, 14845 (2017).
Gough, D. O. Solar interior structure and luminosity variations. Sol. Phys. 74, 21–34 (1981).
Scotese, C. R. & Wright, N. PALEOMAP Paleodigital Elevation MOdels (PaleoDEMS) for the Phanerozoic PALEOMAP Project, EarthByte. EarthByte https://www.earthbyte.org/paleodem-resource-scotese-and-wright-2018 (2018).
Hervé, M. RVAideMemoire: testing and plotting procedures for biostatistics. R package version 0.9-80. R Project https://CRAN.R-project.org/package=RVAideMemoire (2021).
Williams, J. W. & Jackson, S. T. Novel climates, no‐analog communities, and ecological surprises. Front. Ecol. Environ. 5, 475–482 (2007).
Warren, D. L. et al. Environmental niche equivalency versus conservatism: quantitative approaches to niche evolution. Evolution 62, 2868–2883 (2008).
Kamilar, J. M. & Muldoon, K. M. The climatic niche diversity of malagasy primates: a phylogenetic perspective. PLoS ONE 5, e11073 (2010).
Broennimann, O. et al. Measuring ecological niche overlap from occurrence and spatial environmental data. Glob. Ecol. Biogeogr. 21, 481–497 (2012).
Sofaer, H. R. et al. isleading prioritizations from modelling range shifts under climate change. Glob. Ecol. Biogeogr. 27, 658–666 (2018).
Saupe, E. E. et al. Macroevolutionary consequences of profound climate change on niche evolution in marine molluscs over the past three million years. Proc. R. Soc. B https://doi.org/10.1098/rspb.2014.1995 (2014).
Saupe, E. E. et al. Niche breadth and geographic range size as determinants of species survival on geological time scales. Glob. Ecol. Biogeogr. 24, 1159–1169 (2015).
Strotz, L. C. et al. Metabolic rates, climate and macroevolution: a case study using Neogene molluscs. Proc. R. Soc. B 285, 20181292 (2018).
Jones, L. A. et al. Coupling of palaeontological and neontological reef coral data improves forecasts of biodiversity responses under global climatic change. R. Soc. Open Sci. https://doi.org/10.1098/rsos.182111 (2019).
Antell, G. T. et al. Thermal niches of planktonic foraminifera are static throughout glacial–interglacial climate change. Proc. Natl Acad. Sci. USA 118, e2017105118 (2021).
Chiarenza, A. A. et al. Early Jurassic origin of avian endothermy and thermophysiological diversity in dinosaurs. Curr. Biol. 34, 2517–2527 (2024).
Carpenter, G., Gillison, A. N. & Winter, J. Domain: a flexible modelling procedure for mapping potential distributions of plants and animals. Biodivers. Conserv. 2, 667–680 (1993).
Hijmans, R. J. et al. Package ‘dismo’. R Project http://cran.r-project.org/web/packages/dismo/index.html (2011).
Chiarenza, A. A. et al. Climatic constraints on the biogeographic history of Mesozoic dinosaurs. Curr. Biol. 32, 570–585 (2022).
Chiarenza, A. A. et al. Ecological niche modelling does not support climatically-driven dinosaur diversity decline before the Cretaceous/Paleogene mass extinction. Nat. Commun. 10, 1091 (2019).
Chiarenza, A. A. et al. 100 million years of turtle paleoniche dynamics enable the prediction of latitudinal range shifts in a warming world. Curr. Biol. 33, 109–121 (2023).
Foffa, D. et al. Triassic pterosauromorph specimen occurrences and climate variables. Figshare https://figshare.com/s/742a30f30deb4aefb60a (2025).
Foffa, D. et al. Potential of latitudinal dispersion. Figshare https://figshare.com/s/7ac05f57d35b3810a2c7 (2025).
Foffa, D. et al. Accumulated latitudinal dispersion. Figshare https://figshare.com/s/8cabc46601eb64f9b9f0 (2025).
Foffa, D. et al. Palaeoclimate niche occupation. Figshare https://figshare.com/s/6254a692186e79fa9060 (2025).
Foffa, D. et al. Habitat suitability analysis. Figshare https://figshare.com/s/6600233d24db8f1d986e (2025).
Acknowledgements
D.F. is funded by Marie Skłodowska-Curie Actions: Individual (Global) Fellowship (H2020-MSCA-IF-2020; grant 101022550) and by the Royal Commission for the Exhibition of 1851–Science Fellowship in the first stages of this work. E.M.D. acknowledges the support of the Emerging Talents Initiative (FAU). A.A.C. was funded by a European Research Council (ERC) Starting Grant under the European Union’s Horizon 2020 Research and Innovation Programme (grant 947921 "Mapas" to Sara Varela) and by a Juan de la Cierva Formación 2020 Fellowship (grant FJC2020-044836-I) from the Ministry of Science and Innovation and the European Union Next Generation EU/PRTR and a Royal Society Newton International Fellowship (grant NIF\R1\231802). R.J.B., E.M.D., A.F. and D.J.F. acknowledge a Leverhulme Research Project Grant (RPG-2019-365). A.F. and P.J.V. acknowledge NERC grants NE/X015505/1 and NE/X018253/1. A.F. and D.J.F. acknowledge the Leverhulme grant (grant no. RPG-2019-365). A.F. acknowledges NERC funding (grant NE/X013111/1) and the Chinese Academy of Sciences Visiting Professorship for Senior International Scientists (grant 2021FSE0001). S.J.N. was supported by NSF EAR 1943286. This is Petrified Forest National Park Paleontological Contribution No. 97 and a contribution to the Natural History Museum’s Evolution of Life Theme. The silhouettes in the figures are from PhyloPic (http://phylopic.org/) under Creative Commons licenses CC BY-NC-SA 3.0 and CC BY 3.0.
Author information
Authors and Affiliations
Contributions
D.F., E.M.D. and A.A.C. equally contributed to the design and execution of the project. D.F., E.M.D. and A.A.C. performed the analyses, created plots and figures and wrote the first draft of the manuscript. D.F. coordinated activities among all authors and led the reviews. D.J.L., P.J.V. and A.F. contributed to running the palaeoclimate simulations and climate analysis. A.F., D.J.F. and P.J.V. performed modelling experiments, evaluation and analysis. A.F. contributed to the first draft of the paper. B.M.W. assisted D.F. with the implementation of the biogeography analyses. B.T.K., A.D.M., W.G.P. and S.J.N. contributed to assembling the dataset with fieldwork data from the Chinle Formation and Dockum Group. P.M.B., R.J.B., N.C.F. and S.L.B. aided with the original design of the project and experiments and jointly supervised the work. All authors contributed to the writing and approval of the final paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Ecology & Evolution thanks Fabiana Costa, Lawrence Tanner and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Time calibrated informal tree of early avemetatarsalian relationships and the results of DEC biogeography model.
Abbreviations: AR, Argentina; BR, Brazil, Uruguay, Namibia; CH, China, Thailand, Kyrgyzstan; EU, Europe, Russia and Greenland; INT, India, Tanzania, Zambia, Madagascar; wNA, western USA, British Columbia, Mexico and Venezuela; sAF, South Africa, Lesotho, Zimbabwe.
Extended Data Fig. 2 Climatic preferences of Middle–Late Triassic pterosauromorph lineages.
a, Principal Component Analysis (PCA) of variables of climatic importance showing early pterosaurs (n = 23) and all lagerpetids (n = 26). b–e, raincloud plots comparing climatic variables between early pterosaur and all lagerpetids. Results of statistical comparisons are reported in Supplementary Table 2. Abbreviations: MAP, Mean Annual Precipitation; MAT, Mean Annual Temperature; P, precipitation; T, temperature. The box plots display the median and IQR, the upper whiskers extends from the 75th percentile to the largest value and the lower whiskers extends from the 25th percentile to the lowest value.
Extended Data Fig. 3 Climatic suitability of early pterosaurs and lagerpetids through the Anisian–Rhaetian interval.
The symbol * indicates the reference stage used to generate models for the others. Note that there is, to date, no Ladinian–Carnian fossil record of pterosaurs and no Anisian body fossil of lagerpetids.
Extended Data Fig. 4 Absolute precipitation (left) and anomaly precipitation (right) relative to Ladinian.
Note the increased precipitation (blue areas on the right column plots) along the Tethyan Gulf starting in the Carnian, and increased aridity of the equatorial Pangaea in the middle Norian–Rhaetian.
Supplementary information
Supplementary Information
Supplementary discussion, Figs. 1–3, Tables 1 and 2 and references.
Supplementary Data 1
Triassic pterosauromorph occurrences: specimen numbers, stratigraphic and geological information, current and palaeogeographical coordinates and associated values for climate variables of interest.
Source data
Source Data Fig. 3
Pterosauromorph occurrences and associated PCA scores, and values for the climate variables of interest.
Source Data Extended Data Fig. 2
Pterosauromorph occurrences and associated PCA scores, and values for the climate variables of interest.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Foffa, D., Dunne, E.M., Chiarenza, A.A. et al. Climate drivers and palaeobiogeography of lagerpetids and early pterosaurs. Nat Ecol Evol (2025). https://doi.org/10.1038/s41559-025-02767-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41559-025-02767-8
