
Abstract
Gustavus, a positive regulator in arthropod reproduction, features a conserved SPRY and a C-terminal SOCS box ___domain and belongs to the SPSB protein family. The SPSB family, encompassing SPSB1 to SPSB4, plays pivotal roles in higher animals, including immune response, apoptosis, growth, and stress responses. In Neocaridina denticulata sinensis, alternative splicing yielded two NdGustavus isoforms, NdGusX1 and NdGusX2, with distinct expression patterns—high in ovaries and muscles, respectively, and across all ovarian germ cells. These isoforms showed similar expression dynamics during embryogenesis and significant upregulation post-copper ion exposure (P < 0.05). The in situ hybridization result elucidated that NdGusX1 and NdGusX2 were expressed across the germ cell spectrum in the ovary, with NdGusX1 showing enhanced expression in oogonia and primary oocytes. In addition, RNA interference revealed functional complementation in ovaries and potential functional differentiation in muscles. Knockdown of NdGusX1 and NdGusX2 potentially disrupted endogenous vitellogenin synthesis, regulating vitellogenesis and reducing mature oocyte volume, affecting follicular cavity occupation. This study provides a theoretical framework for understanding the biological functions of the SPSB family in crustacean ovarian maturation.
Similar content being viewed by others

Introduction
The SPSB (SPRY ___domain-containing SOCS box) family, comprising four deeply conserved proteins (SPSB1, SPSB2, SPSB3, and SPSB4), has been delineated across diverse animal species, including Homo sapiens, Mus musculus, and Danio rerio1,2,3,4. These proteins are distinguished by preserving SPRY and SOCS box domains5. The SPRY ___domain, originally identified as a sequence homology in dual-specificity kinase splA and the ryanodine receptor is instrumental in orchestrating calcium release from the sarcoplasmic reticulum and facilitates protein–protein interactions. Additionally, proteins harboring the SPRY ___domain contribute to RNA metabolism and biological development6. Furthermore, the B30.2 ___domain is formed by the SPRY ___domain and a conserved sequence at its N-terminus, known as the PRY region6,7,8. SOCS box ___domain was discovered in the suppressors of cytokine signaling9,10. This SOCS family exerts inhibitory control over the cytokine-activated signaling cascade, in part by augmenting the polyubiquitination of signaling intermediates, after their recognition and degradation by the proteasome9,11,12. The SOCS family is involved not only in the suppression of cytokine signaling pathways, but also in hypersensitivity reactions, autoimmunity, and tumorigenesis13,14,15.
Gustavus, identified with both SPRY and C-terminal SOCS box domains in Drosophila melanogaster, belongs to the SPSB family and exerts a positive impact on fertility16,17. Moreover, its association with Vasa is instrumental in the conserved and positive regulation of Vasa accumulation during embryogenesis, which is indispensable for germ cell differentiation16. In D. melanogaster, mutations in Gustavus result in female infertility17,18. A similar finding was observed in the peach aphid Myzus persicae, where silencing of MpGus in adults significantly reduced the number of developing embryos and nymphs19. Furthermore, MpGus was implicated in the mechanism by which potato virus Y (PVY)-infected plants impeded the reproductive potential of aphids20. In Macrobrachium nipponense, the expression patterns of MnGus in different developmental stages of embryos suggest its involvement in embryonic development and oogenesis21. Additionally, Gustavus fulfills pivotal roles in cell proliferation, stress responses, and apoptosis22.
Recently, the annual increase in heavy metal ion content in water, caused by industrial and agricultural activities, has led to significant harm to aquatic organisms23. Copper ions, although indispensable as cofactors for enzymatic reactions within organisms, can exert toxicity at elevated levels, potentially resulting in tissue injury24,25. The ovary, being both a reproductive organ and a significant site for heavy metal ion deposition, is particularly vulnerable to pathological harm caused by these ions26. Studies have indicated that moderate concentrations of copper ions elicit a slight contraction of oocytes, whereas higher concentrations may precipitate cellular lysis within the ovaries26. Consequently, it is crucial to delve into the underlying mechanisms by which organisms mitigate cellular stress responses to curtail the adverse impact on ovarian function.
Neocaridina denticulata sinensis, a freshwater shrimp, exhibits a genomic DNA that is devoid of large-scale replication or deletion phenomena27,28. Given its swift growth rate, robust survivability, extensive geographical distribution, and continuous spawning ability, it serves as an ideal model for comprehending the molecular mechanisms of physiological processes in Decapoda29. Moreover, the body size and weight of female adults exceed those of males, allowing for monosex culture to maximize economic benefit30. During ovarian development, the accumulation of yolk protein increases, accompanied by a steady rise in the gonadosomatic index (GSI). Additionally, individuals achieve sexual maturity between 4 and 6 months of age29, and the extent of ovarian maturity greatly influences the success of embryonic and larval development, ultimately determining the effectiveness of artificial breeding31. Hence, further exploration into the functional genes associated with ovarian maturation is imperative for enhancement. Gustavus, a facilitator of fertility that is inadequately investigated in crustaceans21,22, warrants more comprehensive research. This study elucidates two Gustavus transcripts, NdGusX1 and NdGusX2, generated via alternative splicing in N. denticulata sinensis. This study delved into the expression patterns of NdGusX1 and NdGusX2 in different tissues and embryonic development stages, further investigated their potential sites of action through in situ hybridization, and explored their potential regulatory roles in cellular stress response. Moreover, this study employs RNA interference to discern the functional differentiation or complementary mechanisms exhibited by NdGusX1 and NdGusX2, as well as their contributions to reproductive processes. The findings contribute fundamental understanding of the SPSB gene’s functionality, providing theoretical underpinnings for augmenting the reproductive potential of economically significant crustaceans.
Results
Cloning and characterization of NdGusX1 and NdGusX2
Based on the genomic and transcriptomic data of N. denticulata sinensis, the sequence information of NdGus was obtained. Genomic structure analysis revealed that NdGus underwent alternative splicing event (alternative first exon), resulting in the formation of two transcripts and subsequently translating into two Gustavus proteins, named NdGusX1 and NdGusX2, respectively (Fig. 1a). Both NdGusX1 and NdGusX2 contained 6 exons and 5 introns, exhibiting identical exon positions and sizes, except for the first exon. Full-length cDNA sequences of NdGusX1 and NdGusX2 were amplified by polymerase chain reaction (PCR) (Fig. 1b,c, Supplementary Table 1). The open reading frame (ORF) of NdGusX1 comprised 849 bp, encoding 282 amino acids, while the ORF of NdGusX2 was 2019 bp, encoding 672 amino acids. The molecular weight (MW) of NdGusX1 was 31.84 kDa with a theoretical isoelectric point (pI) of 9.03, whereas the MW of NdGusX2 was 72.72 kDa with a theoretical pI of 6.09. Both NdGusX1 and NdGusX2 contained typical SPRY and SOCS box conserved domains, without signal peptides (Fig. 2a). Additionally, both NdGusX1 and NdGusX2 have continuous β-sheet structures, with two short α-helical structures at N-terminus, which was a typical feature of SPRY/B30.2 ___domain (Fig. 2b). Notably, NdGusX2 formed an α-helical structure between 559 aa and 568 aa, distinguishing from NdGusX1.

The basic information of NdGusX1 and NdGusX2. (a) The genomic structures of NdGusX1 and NdGusX2; (b, c) Sequence information of NdGusX1 and NdGusX2. The nucleotide sequence is displayed in the 5′–3′ direction.

Protein ___domain of NdGusX1 and NdGusX2 (a) and the predicted tertiary structure of NdGusX1 and NdGusX2 (b). The tertiary structure models are based on template d2fnjal, and the different structures of the two proteins are boxed.
Comparative and phylogenetic analysis of NdGusX1 and NdGusX2
The BLASTP analysis unveiled that the amino acid sequence of NdGusX1 exhibited the highest similarity (98.23%) with Gustavus-like proteins of Procambarus clarkii and Homarus americanus. Moreover, the similarities with Gustavus-like proteins in Eriocheir sinensis, Portunus trituberculatus, and Penaeus japonicus were 97.52%, 97.06%, and 96.81%, respectively. For NdGusX2, its amino acid sequence showed the maximum identity (98.18%) with those from P. clarkii and H. americanus, while displaying similarities of 97.45%, 97.06%, and 96.72% with those of E. sinensis, P. trituberculatus, and P. japonicus, respectively. Multiple sequence alignment revealed strong conservation of SPSB homologs in SPRY and SOCS box domains and low identity to SPSB3 for both NdGusX1 and NdGusX2 (Fig. 3, Supplementary Table 2). The SOCS box ___domain consisted of two conserved motifs, separated by five non-conserved residues. Combining the tertiary structure characteristics of the proteins with the results of InterProScan predictive analysis, a conserved region preceding the SPRY ___domain was confirmed to form the B30.2 ___domain along with the SPRY ___domain, indicating the presence of the B30.2 ___domain in both NdGusX1 and NdGusX2.

Multiple sequence alignments of NdGus with 13 orthologs from other species. The underlined is the SPRY ___domain. The double underlined is the SOCS box ___domain, and the percentage values indicate the identity of the amino acid sequences of each protein compared to NdGusX1.
The phylogenetic tree constructed using the Neighbor-Joining method with 1000 bootstrap replicates unveiled the relationship between NdGus and other SPSB proteins across different species (Fig. 4, Supplementary Table 2). The results indicated that NdGusX1 and NdGusX2 were initially clustered together, followed by clustering with Gustavus from M. nipponense and D. melanogaster, and subsequently with the vertebrate SPSB1 and SPSB4. Notably, NdGusX1 and NdGusX2 exhibited the furthest evolutionary relationship with SPSB3, consistent with the results of multiple sequence alignment. Additionally, the branch length of NdGusX2 appeared longer than that of NdGusX1, suggesting a greater evolutionary divergence for NdGusX2.

The phylogenetic tree of the SPSB family from different organisms by the neighbor-joining method with 1000 bootstrap replicates. The bold font is used for the objective proteins.
Expression patterns of NdGusX1 and NdGusX2 in different tissues and different embryonic developmental stages
Using the eyestalk as a control, data analysis showed that NdGusX1 exhibited significantly higher expression in the ovaries compared to other tissues (P < 0.05). The expression of NdGusX1 in the ovary was 34.03 folds higher than that in the eyestalk. The expressions of NdGusX1 in the testis and heart were 7.78 folds and 9.31 folds higher than those in the eyestalk, respectively (Fig. 5a). In contrast, the expression pattern of NdGusX2 in different tissues was inconsistent with NdGusX1. The expression of NdGusX2 in the muscle was 42.12 folds higher than that in the eyestalk. The expressions of NdGusX2 in the gill and testis were 18.75× and 20.36× higher than those in the eyestalk, respectively (Fig. 5a). Moreover, the in situ hybridization experiments revealed that both NdGusX1 and NdGusX2 were localized in all germ cells of the ovary, with NdGusX1 exhibiting higher expression, especially in oogonia and primary oocytes (Fig. 5d).

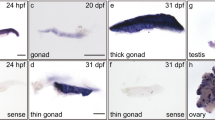
The expression profiles of NdGusX1 and NdGusX2. (a) The expression patterns in mature tissues; (b) The expression patterns in embryos at different developmental stages. 1-cleavage stage, 2-blastula stage, 3-gastrula stage, 4-egg-nauplius stage, 5-egg-metanauplius stage, 6-embryo with eye pigments forming stage, 7-embryo with well-formed eye pigments stage, 8-zoea stage, 9-larval stage; (c) The expression profiles in gonads at different time periods after copper ions stimulation; (d) Localization of NdGusX1 and NdGusX2 in ovary.
To explore the roles of NdGusX1 and NdGusX2 in embryonic development, the expression levels of NdGusX1 and NdGusX2 at different embryonic stages were analyzed (Fig. 5b). It was found that the expression trends of NdGusX1 and NdGusX2 were generally similar across different embryonic developmental stages, showing a trend of down-regulation followed by up-regulation. The lowest expression levels were observed in the egg-nauplius stage and the egg-metanauplius stage and both NdGusX1 and NdGusX2 displayed a significant increase from the embryo with eye pigments forming stage to the larval stage (P < 0.05).
Expression patterns of NdGusX1 and NdGusX2 under Cu2+ stimulation
The SOCS molecule is involved in regulating cytokine signaling and mediating cellular stress responses. Consequently, the expression levels of NdGusX1 and NdGusX2 were examined under Cu2+ stimulation (Fig. 5c). The results demonstrated that following 2.5 μM Cu2+ stimulation, the expression level of NdGusX1 significantly increased at 24 h post-stimulation (P < 0.05), reaching 2.03 times the expression level of the 0 h sample, and returned to normal levels after 72 h. Additionally, the expression level of NdGusX2 significantly increased from 6 to 24 h after Cu2+ stimulation (P < 0.05), reaching its peak at 12 h post-stimulation, being 4.67 times the expression level of the 0 h sample, before gradually returning to normal levels.
The potential functional differentiation and complementary mechanism of NdGusX1 and NdGusX2
Due to the complete sequence identity of NdGusX1 and NdGusX2 in the conservative structural domains, it was speculated that NdGusX1 and NdGusX2 may have a complementary mechanism to ensure the normal progression of specific physiological activities. To explore the potential complementary mechanism between NdGusX1 and NdGusX2, RNA interference technology was employed to knock down NdGusX2, targeted the specific exon of NdGusX2, and the subsequent change in the expression level of NdGusX1 was observed (Fig. 6a). Furthermore, considering the high expression of NdGusX1 and NdGusX2 in the ovaries and muscles, the changes and correlation of their expression levels in these tissues were examined.

Position information for the primers used to prepare double-stranded RNA (a) and the expression level of NdGusX1 and NdGusX2 in the dsNdGusX2-injected experimental group and the dsEGFP-injected control group (b) analyzing by semi-quantitative PCR experiment; (c) Analysis of gray scale based on Image J. E means the dsNdGusX2-injected experimental group and C means the dsEGFP-injected control group.
Following the injection of 5 μL of 4 μg/μL double strand RNA (dsRNA), the expression levels of NdGusX2 in the muscles and ovaries significantly decreased to 48.9% and 73.5% of the control group, respectively (as indicated by the average value) (Fig. 6b,c). The results of semi-quantitative PCR and grayscale analysis revealed that in muscle tissue, the significant decrease in the expression level of NdGusX2 (P < 0.0001) was accompanied by a significant decrease in the expression level of NdGusX1 (P < 0.05) (Fig. 6b,c). However, a divergent phenomenon occurred in the ovaries, wherein the significant decrease in the expression level of NdGusX2 (P < 0.01) was accompanied by a significant increase in the expression level of NdGusX1 (P < 0.05) (Fig. 6b,c).
Effects of NdGusX1 and NdGusX2 on ovarian development
Upon injection of 5 μL of 4 μg/μL dsNdGus, a notable decrease in the expression levels of NdGusX1 and NdGusX2 in the ovaries was observed relative to the control group injected with dsEGFP (P < 0.0001), with expression levels reaching 40.5% and 26.7% of the control group, respectively (as indicated by the average value) (Fig. 7a). The results of semi-quantitative PCR and grayscale analysis indicated that compared to the control group, the expression of Vg slightly increased at 24 h post interference, followed by a significant decrease at 48 h to 96 h post interference (P < 0.001) (Fig. 7b,c). Additionally, the expression of VgR did not exhibit a significant change post-interference (P > 0.05) (Fig. 7b,c). However, at 24 h post-interference, the expression level of Vasa significantly decreased (P < 0.01), and these gene expression levels tended to return to normal with prolonged interference (Fig. 7b,c). Furthermore, a significant decrease in total protein and triglyceride content in the ovaries was observed (P < 0.05), but no significant differences were found in the gonadosomatic index between the experimental and control groups (P > 0.05) (Table 1). Assessment of oocyte development in the ovaries via HE staining showed that at 24 h post interference, the space between the mature oocyte membrane and follicle cell membrane increased, resulting in oocytes no longer filling the follicular cavity and a less dense arrangement of oocytes (Fig. 7d). These observations suggested a relative decrease in the size of oocytes within the follicular cavity and a corresponding reduction in yolk protein accumulation.

The effects of NdGusX1 and NdGusX2 on ovarian development. (a) The detection of interference efficiency of NdGusX1 and NdGusX2; (b) and (c) The expression levels of genes associated in oogenesis after knocking down NdGus (b): Semi-quantitative PCR experiment; (c) Analysis of gray scale based on Image J and the 18S rRNA is used as an internal reference gene); (d) Oocyte development after 24 h of interference.
Discussion
The SPSB family is a class of proteins containing SPRY and SOCS box domains. Although numerous proteins with SPRY or SOCS box ___domain have been discovered, the precise function of SPSB is still vague. In higher animals, the SPSB family has evolved into diversified species, each fulfilling different functions, whereas the evolutionary diversity of the SPSB family in lower animals is comparatively limited, indicating that an SPSB molecule in these animals may perform multiple functions. In H. sapiens and M. musculus, four SPSB molecules have been characterized, namely SPSB1/2/3/43,4. In D. melanogaster, Gustavus was initially discovered and confirmed to be a homologous protein of the SPSB family, exhibiting positive regulation of fertility17. Furthermore, Gustavus was also found in M. nipponense and M. persicae19,21.
In this study, the genomic structure analysis unveils that two different transcripts of NdGus are generated by alternative first exon events, indicating the potential for varied splicing patterns in different tissues and developmental stages. This results in the expression of different proteins and the execution of distinct functions. In M. persicae, MpGus also undergoes alternative splicing events to form three transcript variants19. Despite the differing 5′ terminus of the three cDNA sequences, the SPRY and SOCS box ___domain regions remain identical. Multiple sequence alignment with homologous proteins from diverse species demonstrates that the SOCS box ___domain of NdGusX1 and NdGusX2 is composed of two conserved motifs spacing with 5 amino acids, in line with the reported sequence characteristics described by Hilton5. Interestingly, in D. japonica, there are 23 amino acids in the middle of the two-segment sequence of the SOCS box ___domain22. This difference may be related to the evolutionary process. In addition, the high conservation of homologs within the SPSB family across different species suggests that Gustavus fulfills crucial roles in physiologic activities. The result of phylogenetic analysis implies that NdGusX1 and NdGusX2 are clustered with SPSB1 and SPSB4, indicating similar functions to SPSB1 and SPSB4. The MnGus in M. nipponense is also clustered with SPSB121. Moreover, the longer phylogenetic branch length of NdGusX2 compared to NdGusX1 suggests greater evolutionary divergence and potentially different functions in N. denticulata sinensis.
The expression profiles of genes across different tissues and developmental stages are intricately related to their biological functions. In this study, the expression of NdGusX1 in germ cells of the ovaries suggests its involvement in the regulation of ovarian development. Similarly, analysis of MpGus expression levels in M. persicae shows maximum expression in the abdominal tissue, indicating its potential role in the reproduction of the peach aphid19. Furthermore, SSB-1, SSB-2, and SSB-3 in M. musculus are expressed in the testes, while SSB-1 and SSB-4 are expressed in the ovaries32. Likewise, in D. rerio, SSB-1 and SSB-4 are expressed in the ovaries and testes, with in situ hybridization experiments confirming their presence in germ cells2. Additionally, DjSpsb is expressed in the testes and yolk glands of D. japonica22. Collectively, the SPSB family plays a vital role in the reproductive processes of organisms. Regarding NdGusX2, it is highly expressed in the muscles, consistent with the reported expression pattern of MnGus in M. nipponense21. Previous research has shown that the SPRY ___domain is involved in calcium release from the sarcoplasmic reticulum during muscle contraction33. This suggests that NdGusX2 may have a potential analogous function to MnGus, possibly associated with muscle development. In D. rerio, SSB-1 is also expressed in the muscle2. However, contrasting findings emerge from the study of the SPSB family in M. musculus, where it is observed that SSB-3 exhibits high expression in the muscles, rather than SSB-134.
Similar to Vasa, Gustavus is a component of ribonucleoprotein (RNP) complexes17. RNP complexes are involved in establishing cell polarity and laying a foundation for embryonic development35. The expression patterns of NdGusX1 and NdGusX2 in the different embryonic stages indicate that they are highly expressed in the early stages. This expression pattern aligns with findings in D. rerio, where SSB-1 and SSB-4 also show high expression levels in the early stages of embryonic development (from the zygote stage to the blastula stage), with the highest expression level during the zygote stage2. This could be attributed to the regulation of maternally contributed factors during the early stages of embryonic development, inferring that NdGusX1 and NdGusX2 may be maternal genes36. During the egg-nauplius stage and the egg-metanauplius stage, the expression levels of NdGusX1 and NdGusX2 are significantly lower than in other developmental stages, indicating rapid embryonic development and extensive consumption of maternally sourced nutrients. As the embryos develop, the expression levels of NdGusX1 and NdGusX2 notably increase. This increasing trend is supported by similar reports in M. persicae, suggesting that the increased expression may be closely linked to embryonic development and reproductive processes19. Additionally, the expression level of NdGusX2 increases remarkedly in the larval stage, possibly due to its role in muscle development during the later stages of embryonic development. Similarly, in the later stages of embryonic development in M. nipponense, MnGus also exhibits a comparable expression pattern21.
The copper ion is a vital trace element that acts as a cofactor for enzymes. However, extensive usage of copper sulfate to eliminate algae from ponds leads to a significant increase in copper ion concentrations in the water, surpassing the normal requirements of aquatic organisms and severely disrupting their normal physiological functions. Elevated copper level is detrimental to the organism, leading to tissue damage24,25. Previous research has indicated that low concentrations of copper ions may cause slight shriveling of oocytes, while high concentrations may lead to oocyte rupture26. Furthermore, SOCS molecules are involved in regulating cytokine signaling pathways and mediating cellular stress responses9,11,12. The expression patterns of NdGusX1 and NdGusX2 under copper ion stress are analyzed to evaluate if external copper ion stimulation can induce their expression, thereby reducing the impact of copper ions on gonadal tissue. The results demonstrate that under copper ions stimulation, the expression levels of NdGusX1 and NdGusX2 significantly increased, reaching peak levels at 24 h and 12 h after stimulation, respectively, and then returning to normal levels. These findings align with those observed in D. japonica22. Under 24 °C heat stress, the expression level of DjSpsb is slightly higher than that of the control group, without a significant difference. However, after 3 days of incubation at 30 °C, the DjSpsb expression level increases significantly. When exposed to ethanol-induced stress, the expression level of DjSpsb is significantly up-regulated in a dose-dependent manner, and the expression level reaches a peak at 2% ethanol concentration. These observations suggest that Spsb may modulate the cellular stress response by altering its expression level under various stress stimuli.
RNA interference is a widely utilized approach for investigating gene function37. Several studies have demonstrated that conducting RNA interference experiments in crustaceans is currently the most effective and cost-efficient method38. In this research, RNA interference is employed to knock down NdGusX2, and the expression levels of NdGusX1 in the ovaries and muscles are measured to explore the potential complementary effects between NdGusX1 and NdGusX2 in these tissues. It is found that a significant decrease in the expression level of NdGusX2 leads to a marked increase in the expression level of NdGusX1 in ovaries, while the expression level of NdGusX1 in muscles declines significantly, indicating potential functional complementation between NdGusX1 and NdGusX2 in ovaries, and functional differentiation in muscles. Investigating the roles of NdGusX1 and NdGusX2 in muscles may provide new insights into the mechanism of alternative splicing of NdGus.
The process of ovarian maturation entails the ongoing accumulation of vitellogenin. As a precursor of vitellogenin, Vg plays a crucial role in oogenesis. Previous research has shown that the continuous decrease in Vg expression delays ovarian maturation39. The current research elucidates that a significant reduction in the expression levels of NdGusX1 and NdGusX2 results in a remarked decrease in Vg expression after dsNdGus injection for 48 h, indicating that NdGus may control the ovarian maturation by regulating Vg expression. Similar conclusions have been drawn in research on Gustavus in M. nipponense21. VgR can bind and facilitate the entry of exogenous Vg into oocytes, highlighting its important role in oogenesis39. Following NdGus knockdown, the expression level of VgR decreases, albeit not significantly, and returns to normal levels at 96 h, suggesting that knocking down NdGus affects the accumulation of endogenous Vg but not the synthesis and transport of exogenous Vg. Vasa has been proven to be an interacting protein of Gustavus in previous studies16,17. When the expression level of NdGus decreases, the expression level of Vasa also decreases significantly at 24 h after interference, then returns to normal levels, indicating that Vasa may be regulated by other genes within the organism and plays an important role in ovarian development.
Proteins and lipids serve as the primary nutrients and energy substances in crustacean oocytes, not only participating in the formation of cell membranes but also providing nourishment and energy for early embryonic development31. In addition, vitellogenesis involves the continuous accumulation of nutrients including proteins and lipids. This research demonstrates that silencing NdGus using RNA interference technology leads to a notable reduction in the total protein and triglyceride content in the ovaries. Although the ovarian gonadosomatic index does not display significant differences, the oocytes diminish in size, failing to occupy the entire follicular cavity, resulting in a comparatively limited accumulation of yolk substances in the cytoplasm. Based on the research results, it can be inferred that NdGus influences ovarian development in N. denticulata sinensis by controlling the accumulation of nutrients in oocytes.
In summary, NdGus generates two transcripts (NdGusX1 and NdGusX2) due to alternative first exon events. NdGusX1 and NdGusX2 are highly conserved across different species and show high homology with SPSB1 and SPSB4. NdGusX1 and NdGusX2 mediate the synthesis of endogenous vitellogenin, thereby regulating vitellogenesis and further influencing ovarian maturation. Moreover, NdGusX1 and NdGusX2 may also mediate cellular stress responses by adjusting their expression levels to mitigate the impact of stimuli on gonadal tissues. Additionally, NdGusX2 seems to be involved in muscle development. This research lays a theoretical foundation for the potential application of ripening technology in aquaculture.
Methods
Experimental animals and tissues collection
Healthy shrimps, measuring 20 ± 1.5 mm in length and weighing 0.12 ± 0.05 g (mean ± SEM), were obtained from the aquaculture market in Baiyangdian, Baoding. Then, all shrimps were cultivated in a circulating water system equipped with an aeration pump, maintaining the water temperature at 25 ± 1 °C (light:dark = 1:1). Prior to being utilized for further research, all experimental shrimps underwent a week of acclimatization and were fed a daily commercial diet (Sera shrimps natural, Kunshan). Additionally, different tissues were individually dissected from over three shrimps under sterile conditions. The surveyed samples were promptly frozen with liquid nitrogen and used for RNA extraction afterward or stored at − 80 °C.
Total RNA extraction and cDNA synthesis
Mature shrimp tissues were used to extract total RNA, employing the RNAsimple Total RNA Kit (TIANGEN, China) in accordance with the manufacturer’s protocol. The concentration and quality of the obtained RNA sample were assessed by NanoDrop 2000 spectrophotometry and 1% agarose gel electrophoresis. Subsequently, high-quality samples underwent reverse transcription reactions performed by HiScript II Reverse Transcriptase and gDNA wiper Mix kit (Vazyme, Nanjing) under the manufacturer’s presentation. The gDNA wiper Mix in the kit was used to remove DNA. The reactions were performed at 42 °C for 20 min, terminated by heating at 85 °C for 5 s. Measure the OD260/280 of cDNA using NanoDrop 2000c (Thermo Scientific, USA) to ensure the quality of cDNA. Dilute the cDNA sample 10 times for the template of real-time quantitative polymerase chain reaction (RT-qPCR), and further confirm the quality of cDNA by checking whether the CT value of 18S rRNA falls within the range of 9–18. All cDNA samples were then stored at − 20 °C for RT-qPCR analysis.
Cloning and sequence analysis of full-length NdGus cDNA
Based on the genomic and transcriptomic data of N. denticulata sinensis, the ORF of NdGus was predicted and further amplified with the specific primers (Supplementary Table 1). The PCR products were verified by agarose gel electrophoresis, and the correct bands were cut for DNA recovery. The target DNA fragment was ligated to the pMD19-T vector (Takara, Dalian) and then transformed to E. coli DH5ɑ competent cells (WEIDI, Shanghai). The positive clones were screened and sequenced at GENEWIZ Co., Ltd. (Tianjin, China).
The gene structures of NdGus (NdGusX1 and NdGusX2) were analyzed according to existing genomic and transcriptomic data. The basic physicochemical properties, functional domains, and tertiary structures of NdGusX1 and NdGusX2 proteins were predicted by the ExPASy-ProtParam (http://www.expasy.org/tools/protparam.html), SMART (https://smart.embl.de/), and PHYRE2 (http://www.sbg.bio.ic.ac.uk/phyre2/), respectively40. Additionally, homologous protein sequences of NdGus were obtained from the GenBank database, and multiple sequence alignment was visualized by DNAMAN software. MEGA11 software and the iTOL online website (https://itol.embl.de/) were used to construct and beautify NJ phylogenetic trees for SPSBs41.
Expression pattern of NdGus in different tissues, embryos, and gonads stimulated by copper ions
To investigate the tissue distribution of NdGus, gills, intestines, muscles, hepatopancreas, eyestalk, stomach, and heart were collected from three female and three male shrimps. Additionally, testes and ovaries were collected from six male and six female shrimps, respectively. Embryos at different developmental stages, including cleavage stage, blastula stage, gastrula stage, egg-nauplius stage, egg-metanauplius stage, embryo with eye pigments forming stage, embryo with well-formed eye pigments stage, zoea stage, and larval stage, were obtained for expression pattern analysis. The ovaries and testes were collected after the shrimps were stimulated by copper ions for 0, 6, 12, 24, 36, 48, and 72 h, aiming to explore the roles of NdGusX1 and NdGusX2 in cellular stress response.
The expression patterns of NdGusX1 and NdGusX2 were analyzed in different tissues, embryos at different developmental stages, and gonadal samples stimulated by copper ions, with 18S rRNA serving as the internal reference gene. A negative control group was set up using DEPC water as the template. Specific primers for RT-qPCR were designed by Primer Premier 5.0 (Supplementary Table 1). The reaction system of qRT-PCR was 10 μL volume including 5 μL 2 × Universal SYBR Green Fast qPCR Mix (ABclonal® Technology, Wuhan), 0.2 μL primers (10 mM), and 0.5 μL cDNA. To ensure primer specificity, annealing temperatures were explored using gradient PCR. The reaction procedure of RT-qPCR was set including 95 °C for 3 min, 35 cycles of 95 °C for 10 s, 55 °C for 10 s, and 72 °C for 15 s, 55 °C for 1 min, 95 °C for 15 s. Additionally, to ensure that the amplification product was a single expansion, melting curves were used by the program from 60 to 95 °C at a rate of 0.4 °C/s (Bioer LineGene 9600, Hangzhou). Each sample was performed in quadruplicate, and the relative expression level was calculated by the 2−ΔΔCT method42.
In situ hybridization
Digoxigenin-labeled RNA probes, targeting the common region of NdGusX1 and NdGusX2 and the specific region of NdGusX2, were synthesized using T7 RNA transcriptase and then stored at − 20 °C (Supplementary Table 1). Ovaries harvested from shrimp at the early vitellogenesis stage were fixed overnight at 4 °C in a 4% paraformaldehyde fixative solution. The samples underwent sequentially dehydration in 50% ethanol, 75% ethanol, 95% ethanol, and 100% ethanol, transparent treatment in xylene, and embedding in paraffin. Subsequently, the embedded tissue was sliced to 5 μm for ISH. ISH for ovarian tissues was performed as described in the literature43. The slices were then digested by proteinase K at 37 °C for 30 min, prehybridized at 55 °C for 4 h, and hybridized with a digoxigenin (DIG)-labeled RNA probe (Roche, Mannheim, Germany) against NdGusX1/2 mRNA at 55 °C overnight. Then, anti-digoxigenin-AP Fab fragments (Roche, Mannheim, Germany) were incubated overnight at 4 °C in slices. Finally, signals were developed using the BCIP/NBT Chromogen Kit (Solarbio, Beijing).
RNAi methodology
The linearized DNA template for transcription was obtained by PCR with primers containing the T7 promoter sequence at the 5′ terminus (Supplementary Table 1). The double-stranded RNA (dsRNA) was synthesized from a 1 μg DNA template using the T7 High Yield Transcription Kit (Thermo Fisher Scientific, American). Subsequently, the synthesized dsRNA was then purified and its concentration and quality were tested. Shrimps at the same ovarian developmental stage and with similar body sizes were randomly selected as the subjects of the RNA interference assay. The purified dsRNA was diluted into different concentration gradients with DEPC-treated water (Sangon, Beijing), and the final injection concentration of dsRNA was determined by pre-experiment. The experimental group was injected with 5 μL of dsNdGus/dsNdGusX2 at a concentration of 4 μg/μL per tail, and the control group was injected with 5 μL of dsEGFP at a dose of 4 μg/L per tail. The experimenter held the sides of the shrimp’s abdomen with the thumb and middle finger of the left hand and then operated a micro-syringe with the right hand to administer a sterile needle into the second abdomen of the shrimp thereby delivering dsRNA into the shrimp’s body. Tissues were collected 24 h after injection, and the total RNA was extracted from the samples, and further reverse transcribed into cDNA. The interference efficiency was detected by RT-qPCR to determine the optimized injection concentration, which was used for formal experiments.
Functional differentiation and complementary mechanism of NdGusX1 and NdGusX2
The shrimps, all at the same ovarian developmental stage and with similar body size, were randomly divided into two groups. The experimental group was injected with dsNdGusX2, while the control group received dsEGFP injections. Post-injection 24 h, muscles, and ovaries were accurately collected and placed into liquid nitrogen for RNA extraction and subsequent cDNA synthesis. The efficiency of RNA interference was then determined, and the change of NdGusX1 and NdGusX2 expressions in muscles and ovaries was observed.
Effects of NdGusX1 and NdGusX2 on ovarian development
The ovaries of shrimps were dissected at 24, 48, 72, and 96 h after injection to assess interference efficiency and the expression levels of Vg, VgR, and Vasa related to oogenesis. In addition, the total protein content in the ovaries was measured using the Coomassie Brilliant Blue G-250-based colorimetric method (Solarbio, Beijing), and the triglyceride content was determined by the GPO-PAP method44,45. After 24 h of RNA interference, the weight of the broodstock was measured, followed by the accurate weighing of the broodstock ovaries. Subsequently, the GSI of the broodstock was calculated by the following formula:
Wgonad, the weight of the gonad (g); Wbroodstock, the weight of the broodstock (g).
Hematoxylin–eosin staining
The ovaries were fixed with 4% paraformaldehyde fixative solution overnight, and then soaked in double distilled water for 12 h. The samples sequentially underwent dehydration in gradient ethanol concentrations (50–100%), transparent treatment in xylene, and embedding in paraffin wax. Subsequently, the paraffin-embedded tissue was sliced into a 5 μm section, which sequentially underwent deparaffinization in xylene, and rehydration in 100% ethanol, 95% ethanol, 85% ethanol, 70% ethanol, 50% ethanol, and double distilled water. Finally, the section was stained with hematoxylin–eosin, followed by observation under microscopy46.
Statistical analysis
The relative expression levels of genes were calculated by the 2−ΔΔCT method, and the data were processed and analyzed using GraphPad Prism software (Version 9.0.2, San Diego, USA). All data were expressed as the mean ± standard deviation (N = 3), and statistical analysis was performed using SPSS Statistics 19 software. A one-way analysis of variance (ANOVA) and Duncan’s multiple tests were used for data analysis in gene expression and relative activity. The t-test method was used to analyze the significant differences between the samples, with significant differences between the groups represented by different letters (P < 0.05). In contrast, the non-significant differences were indicated by the same letters (P > 0.05). Additionally, the gray scales of the bands were detected and analyzed using ImageJ software.
Data availability
Data will be made available on request. If someone wishes to request data from this study, please contact Jiquan Zhang by email ([email protected]).
References
Gerhard, D. S. et al. The status, quality, and expansion of the NIH full-length cDNA project: The Mammalian Gene Collection (MGC). Genome Res. 14, 2121–2127. https://doi.org/10.1101/gr.2596504 (2004).
Li, J. Z. et al. Cloning and characterization of the SSB-1 and SSB-4 genes expressed in zebrafish gonads. Biochem. Genet. 47, 179–190. https://doi.org/10.1007/s10528-008-9215-1 (2009).
Ota, T. et al. Complete sequencing and characterization of 21,243 full-length human cDNAs. Nat. Genet. 36, 40–45. https://doi.org/10.1038/ng1285 (2004).
Xing, Y., Gosden, R., Lasko, P. & Clarke, H. Murine homologues of the Drosophila gustavus gene are expressed in ovarian granulosa cells. Reproduction 131, 905–915. https://doi.org/10.1530/rep.1.01046 (2006).
Hilton, D. J. et al. Twenty proteins containing a C-terminal SOCS box form five structural classes. Proc. Natl. Acad. Sci. U. S. A. 95, 114–119. https://doi.org/10.1073/pnas.95.1.114 (1998).
Rhodes, D. A., de Bono, B. & Trowsdale, J. Relationship between SPRY and B30.2 protein domains. Evolution of a component of immune defence?. Immunology 116, 411–417. https://doi.org/10.1111/j.1365-2567.2005.02248.x (2005).
Henry, J., Ribouchon, M.-T., Offer, C. & Pontarotti, P. B30.2-like ___domain proteins: A growing family. Biochem. Biophys. Res. Commun. 235, 162–165. https://doi.org/10.1006/bbrc.1997.6751 (1997).
Woo, J. S. et al. Structural and functional insights into the B30.2/SPRY ___domain. EMBO J. 25, 1353–1363. https://doi.org/10.1038/sj.emboj.7600994 (2006).
Kamura, T. et al. The Elongin BC complex interacts with the conserved SOCS-box motif present in members of the SOCS, ras, WD-40 repeat, and ankyrin repeat families. Genes Dev. 12, 3872–3881. https://doi.org/10.1101/gad.12.24.3872 (1998).
Starr, R. et al. A family of cytokine-inducible inhibitors of signalling. Nature 387, 917–921. https://doi.org/10.1038/43206 (1997).
Kile, B. T. et al. The SOCS box: A tale of destruction and degradation. Trends Biochem. Sci. 27, 235–241. https://doi.org/10.1016/s0968-0004(02)02085-6 (2002).
Zhang, J. G. et al. The conserved SOCS box motif in suppressors of cytokine signaling binds to elongins B and C and may couple bound proteins to proteasomal degradation. Proc. Natl. Acad. Sci. U. S. A. 96, 2071–2076. https://doi.org/10.1073/pnas.96.5.2071 (1999).
Babon, J. J., Sabo, J. K., Zhang, J. G., Nicola, N. A. & Norton, R. S. The SOCS box encodes a hierarchy of affinities for Cullin 5: Implications for ubiquitin ligase formation and cytokine signalling suppression. J. Mol. Biol. 387, 162–174. https://doi.org/10.1016/j.jmb.2009.01.024 (2009).
Kuang, Z. H. et al. The SPRY ___domain-containing SOCS box protein SPSB2 targets iNOS for proteasomal degradation. J. Cell Biol. 190, 129–141. https://doi.org/10.1083/jcb.200912087 (2010).
Lewis, R. S. et al. TLR regulation of SPSB1 controls inducible nitric oxide synthase induction. J. Immunol. 187, 3798–3805. https://doi.org/10.4049/jimmunol.1002993 (2011).
Gustafson, E. A., Yajima, M., Juliano, C. E. & Wessel, G. M. Post-translational regulation by gustavus contributes to selective Vasa protein accumulation in multipotent cells during embryogenesis. Dev. Biol. 349, 440–450. https://doi.org/10.1016/j.ydbio.2010.10.031 (2011).
Styhler, S., Nakamura, A. & Lasko, P. VASA localization requires the SPRY-___domain and SOCS-box containing protein, GUSTAVUS. Dev. Cell 3, 865–876. https://doi.org/10.1016/s1534-5807(02)00361-1 (2002).
Kugler, J. M., Lem, C. & Lasko, P. Reduced cul-5 activity causes aberrant follicular morphogenesis and germ cell loss in Drosophila oogenesis. PloS ONE 5, e9048. https://doi.org/10.1371/journal.pone.0009048 (2010).
Gao, Y. et al. The gustavus gene can regulate the fecundity of the green peach aphid, Myzus persicae (Sulzer). Front. Physiol. 11, 596392. https://doi.org/10.3389/fphys.2020.596392 (2021).
Ziebell, H. et al. Cucumber mosaic virus and its 2b RNA silencing suppressor modify plant-aphid interactions in tobacco. Sci. Rep. 1, 187. https://doi.org/10.1038/srep00187 (2011).
Zhang, F. Y. et al. cDNA cloning and expression analysis of Gustavus gene in the oriental river prawn Macrobrachium nipponense. PloS ONE 6, e17170. https://doi.org/10.1371/journal.pone.0017170 (2011).
Dong, Z. M. et al. Identification and expression analysis of a Spsb gene in planarian Dugesia japonica. Gene 564, 168–175. https://doi.org/10.1016/j.gene.2015.03.032 (2015).
Pan, K. & Wang, W. X. Trace metal contamination in estuarine and coastal environments in China. Sci. Total Environ. 421, 3–16. https://doi.org/10.1016/j.scitotenv.2011.03.013 (2012).
De Boeck, G., Meeus, W., De Coen, W. & Blust, R. Tissue-specific Cu bioaccumulation patterns and differences in sensitivity to waterborne Cu in three freshwater fish: Rainbow trout (Oncorhynchus mykiss), common carp (Cyprinus carpio), and gibel carp (Carassius auratus gibelio). Aquat. Toxicol. 70, 179–188. https://doi.org/10.1016/j.aquatox.2004.07.001 (2004).
Harris, E. D. Copper homeostasis: The role of cellular transporters. Nutr. Rev. 59, 281–285. https://doi.org/10.1111/j.1753-4887.2001.tb07017.x (2001).
Bhat, R. A., Bakhshalizadeh, S., Guerrera, M. C., Kesbic, O. S. & Fazio, F. Toxic effect of heavy metals on ovarian deformities, apoptotic changes, oxidative stress, and steroid hormones in rainbow trout. J. Trace Elem. Med. Biol. 75, 127106. https://doi.org/10.1016/j.jtemb.2022.127106 (2023).
Kenny, N. J. et al. Genomic sequence and experimental tractability of a new decapod shrimp model, Neocaridina denticulata. Mar. Drugs 12, 1419–1437. https://doi.org/10.3390/md12031419 (2014).
Sin, Y. W. et al. Identification of putative ecdysteroid and juvenile hormone pathway genes in the shrimp Neocaridina denticulata. Gen. Comp. Endocrinol. 214, 167–176. https://doi.org/10.1016/j.ygcen.2014.07.018 (2015).
Mykles, D. L. & Hui, J. H. L. Neocaridina denticulata: A decapod crustacean model for functional genomics. Integr. Comp. Biol. 55, 891–897. https://doi.org/10.1093/icb/icv050 (2015).
Toyota, K. et al. Sex determination and differentiation in decapod and cladoceran crustaceans: An overview of endocrine regulation. Genes 12, 305. https://doi.org/10.3390/genes12020305 (2021).
Parma, L. et al. Fatty acid composition of eggs and its relationships to egg and larval viability from domesticated common sole (Solea solea) breeders. Reprod. Domest. Anim. 50, 186–194. https://doi.org/10.1111/rda.12466 (2015).
Masters, S. L. et al. Genetic deletion of murine SPRY ___domain-containing SOCS box protein 2 (SSB-2) results in very mild thrombocytopenia. Mol. Cell Biol. 25, 5639–5647. https://doi.org/10.1128/Mcb.25.13.5639-5647.2005 (2005).
Ponting, C., Schultz, J. & Bork, P. SPRY domains in ryanodine receptors (Ca(2+)-release channels). Trends Biochem. Sci. 22, 193–194. https://doi.org/10.1016/s0968-0004(97)01049-9 (1997).
Kleiber, M. L. & Singh, S. M. Divergence of the vertebrate sp1A/ryanodine receptor ___domain and SOCS box-containing (Spsb) gene family and its expression and regulation within the mouse brain. Genomics 93, 358–366. https://doi.org/10.1016/j.ygeno.2008.11.011 (2009).
Kress, T. L., Yoon, Y. J. & Mowry, K. L. Nuclear RNP complex assembly initiates cytoplasmic RNA localization. J. Cell Biol. 165, 203–211. https://doi.org/10.1083/jcb.200309145 (2004).
Pelegri, F., Knaut, H., Maischein, H. M., Schulte-Merker, S. & Nusslein-Volhard, C. A mutation in the zebrafish maternal-effect gene nebel affects furrow formation and vasa RNA localization. Curr. Biol. 9, 1431–1140. https://doi.org/10.1016/s0960-9822(00)80112-8 (1999).
Zhao, J. H. & Guo, H. S. RNA silencing: From discovery and elucidation to application and perspectives. J. Integr. Plant Biol. 64, 476–498. https://doi.org/10.1111/jipb.13213 (2022).
Hiruta, C. et al. Development of a microinjection system for RNA interference in the water flea Daphnia pulex. BMC Biotechnol. 13, 96. https://doi.org/10.1186/1472-6750-13-96 (2013).
Bai, H. K. et al. Molecular characterization and developmental expression of vitellogenin in the oriental river prawn Macrobrachium nipponense and the effects of RNA interference and eyestalk ablation on ovarian maturation. Gene 562, 22–31. https://doi.org/10.1016/j.gene.2014.12.008 (2015).
Schultz, J., Milpetz, F., Bork, P. & Ponting, C. P. SMART, a simple modular architecture research tool: Identification of signaling domains. Proc. Natl. Acad. Sci. U. S. A. 95, 5857–5864. https://doi.org/10.1073/pnas.95.11.5857 (1998).
Saitou, N. & Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425. https://doi.org/10.1093/oxfordjournals.molbev.a040454 (1987).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402–408. https://doi.org/10.1006/meth.2001.1262 (2001).
Wang, B. et al. Comparative studies on duplicated tdrd7 paralogs in teleosts: Molecular evolution caused neo-functionalization. Comp. Biochem. Physiol. Part D Genomics Proteomics 30, 347–357. https://doi.org/10.1016/j.cbd.2019.04.006 (2019).
Grintzalis, K., Georgiou, C. D. & Schneider, Y. J. An accurate and sensitive Coomassie Brilliant Blue G-250-based assay for protein determination. Anal. Biochem. 480, 28–30. https://doi.org/10.1016/j.ab.2015.03.024 (2015).
Luley, C. et al. Point-of-care testing of triglycerides: Evaluation of the Accutrend triglycerides system. Clin. Chem. 46, 287–291 (2000).
Ren, Y. H. et al. Insight of vitellogenesis patterns: A comparative analysis of the differences between the primary and secondary vitellogenesis period in the ovary, hepatopancreas, and muscle of mud crab, Scylla paramamosain. Front. Genet. 13, 965070. https://doi.org/10.3389/fgene.2022.965070 (2022).
Acknowledgements
This work was supported by the National Natural Science Foundation of China (Grant Nos. 32172954, 32373121), Key Research and Development Project of Hebei Province (22323201D), Science Research Project of Hebei Education Department (ZD2022093), The Natural Science Foundation of Hebei Province of China (D2022201003, D2023201002).
Author information
Authors and Affiliations
Contributions
M.L.: Methodology, Investigation, Writing—original draft, Writing—review and editing. D.F.: Methodology, Writing—review. J.Z.: Conceptualization, Writing—review and editing, Funding acquisition. Y.S.: Conceptualization, Writing—review and editing, Supervision.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
According to the local legislation and institutional requirements, ethical approval is not required for the study involving animals (shrimp), which are not endangered invertebrates. In addition, no genetically modified organism was used in the study. According to the Fisheries Law of the People’s Republic of China, no permission is required to collect the animals for this study. @@All methods involved in this study were conducted under the guidelines and regulations of the ARRIVE guidelines (https://arriveguidelines.org). The experimental protocols involved in this study have been authorized by Hebei University.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Liang, M., Feng, D., Zhang, J. et al. Functional complementation of two splicing variants of Gustavus in Neocaridina denticulata sinensis during ovarian maturation. Sci Rep 14, 20939 (2024). https://doi.org/10.1038/s41598-024-72080-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-72080-0
