
Abstract
Land use changes soil microbial and chemical properties, but the mechanism of biological nitrogen fixation under different land use patterns is rarely reported, so we used four types of soil: Natural forest soil (NS), healthy banana soil (HS), diseased banana soil (DS) and paddy soil (PS). Treatments included the control (CK), addition of glucose (G), addition of glucose and ammonium nitrate (GN), addition of banana straw (BS), addition of banana straw and ammonium nitrate (BSN), addition of banana root (BR), and addition of banana root and ammonium nitrate (BRN). The study found that the change of soil utilization types, glucose addition increased carbon dioxide emissions (Compared with the control, increased by 963.11%, 508.39%, 794.77% and 511.34%, respectively) and enhanced the ability of soil microbial nitrogen fixation. Importantly, natural forest soil microorganisms have a higher biological nitrogen fixation capacity compared to other types of soils. Glucose addition caused the accumulation of ammonium nitrogen (Compared with the control, increased by 426.08%, 934.21%, 420% and 1065.95%, respectively), indicating that microorganisms had higher utilization efficiency of soluble carbon and enhanced the biological nitrogen fixation capacity, and nitrogen addition caused the accumulation of ammonium nitrogen, thereby weakening the biological nitrogen fixation capacity. At the same time, glucose significantly increased the Fimicutes phylum (83.73%, 66.38%, 67.18% and 70.36%) and lowered the level of other bacterial phylums, thereby reducing the bacterial network structure, and the stability of the soil environment has decreased. Forest analysis showed that CO2 was an important factor in predicting the bacterial community structure of different soil types, an increase in CO2 content can predict drastic changes in the bacterial community. Bacteria at the Fimicutes phylum level preferred glucose, which may also have a negative effect on bacteria at the level of other phylums.
Similar content being viewed by others


1. Introduction
Soil microorganisms are an important part of the ecosystem, and they drive soil processes of function, including carbon and nitrogen fixation1,2. Soil microbes are play an important role in essential biological functions that improve soil development of health and inhibit plant disease3, such as by promoting nutrient cycle, SOM conversion, plant production and contributing to the suppression of soil-borne diseases4. As a result of the rapid response of bacteria with changes under the soil condition, bacterial community has served as early biological indicator of soil quality5. Previous results have shown that fertilization impacts soil microbes and properties by addition of nutrients6. Appropriate N input leads to increased soil N content, reduces the restriction of N utilization by soil microorganisms, and promotes the growth of bacterial communities7,8. In forest ecosystems in southern China, N addition reduced soil bacterial community α-diversity because the increase of NO3− reduced soil pH and thus bacterial community α-diversity9, while in grassland and farmland ecosystems, N deposition on bacterial communities was not significant influence10,11. Bacterial biomass generally respond to other factors such as changes in litter and plant root exudates may contribute to the response of bacterial communities to N inputs12.
Tropical soils are of global importance because they store large amounts of terrestrial carbon13, and soil microbes play an important role in regulating C store by the mineralization of straw and SOM. Soil microorganisms are the major decomposers of crop residues because of their specific capability to produce enzymes that decompose both simple molecules and more complex plant-derived compounds14. Crop residues addition is an important way to promote soil fertility and improve crop growth, which is important sustainable agriculture development15,16. Microorganisms can be major drivers of nutrient biogeochemical cycles through their role in breaking down straw in soil. There is a large amount of evidence showing that different straw features and factors of environment can result in substantial changes in the diversity of soil microbial communities17,18. Ma et al. also found this positive and significant effect where straw addition fortifies soil microbial biomass, activity and community structure19,20, and also augments the structure of soil particles that improve soil quality21,22. A previous study found that a significant increase in bacterial and actinomycete biomass occurred under short-term addition of crop straw to a field in Jiang Yan, China5. An easily-absorbed carbon source, glucose can be used rapidly by soil microorganisms. Different amounts of glucose were added to the soil, the greater amounts increased CO2 flux during a 49-day experiment23. CO2 flux, MBC, and dissolved organic C is influenced by the amount of glucose added to the soil. Glucose as an exogenous carbon source to study priming effects, because of glucose is a compound lacking nitrogen, and the application of glucose can even eliminate the effects of nitrogen. Glucose can directly participate in the soil carbon metabolism processes as a source of carbon and energy24.
Biological nitrogen fixation is as the second-largest contributor to nitrogen in soil, comprising approximately 16% of the global nitrogen input25. Nitrogen fertilizer addition inhibited the growth of nitrogen fixation microorganisms, decreasing in the diversity of nitrogen fixation soil microorganisms, and the nitrate nitrogen emerging as the main influencing factors26. However, appropriate nitrogen fertilizer addition can increase soil organic carbon content and abundance of nitrogen-fixing bacteria, thereby improving nitrogen fixation capability of soil microorganisms27. Furthermore, other researchers found that abundance and nitrogen fixation bacteria have not significant differences following nitrogen addition28,29. In conclusion, we propose several hypotheses, (1) Different types of carbon (such as glucose, rice straw and roots) combined with nitrogen addition affect soil bacteria and chemical properties under types of soils; (2) The addition of organic carbon (glucose) promotes the nitrogen fixation capacity of different types of soil microorganisms, especially natural forest soils. We hope to find out the differences in the effects of different types of carbon addition on biological nitrogen fixation in different types of soils, and provide a theoretical basis for the screening of nitrogen-fixing microorganisms in the future.
2. Materials and methods
2.1 Study site description and sampling
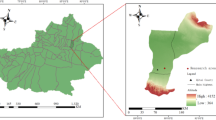
Samples were collected from Natural forest soil (NS), Healthy banana soil (HS), Diseased banana soil (DS, high Fusarium oxysporum f. sp. cubense), and Paddy soil (PS). The sampling locations were adjacent to each other, and HS, DS, and PS all developed from NS, originally in the same ___location in Fig. 1. (HS, DS, and PS evolved from NS.) The soil is classified as lateritic soil that developed from basalt parent material. The four types of soil used in the experiment were gathered in Chengmai, Hainan, China (19°23’, 110°15’). Soil samples were collected from the top 20 cm of soil at four randomly chosen locations. A portion of soil samples were used for soil physicochemical property analysis, and others were stored in -30 °C for DNA extraction.

Distribution of soil sampling sites.
2.2 Experimental design
Soil physicochemical properties were determined from the samples collected from sites representing four different land uses: NS, HS, DS, and PS (Table 1). Samples (10 g) were added to 120 mL culture flasks, along with glucose (8 mg/g soil), ammonium nitrate (1 mg/g soil), organic materials, and 15N2 (15N2: O2: Ar = 20:20:60), banana straw and roots (0.15 g), see Table 1 for all C and N content). Next, they were incubated for 42 days under 28℃ and 60% of field water capacity, and seven treatments were set up (Fig. 2): control (CK), addition of glucose (G), addition of glucose and ammonium nitrate (GN), addition of banana straw (BS), addition of banana straw and ammonium nitrate (BSN), addition of banana root (BR), addition of banana root and ammonium nitrate (BRN). Each treatment was replicated thrice. Gas is collected every seven days to calculate cumulative CO2 emissions and then replace fresh culture gas, determine whether the environment is anaerobic conditions before collection. At the end of incubation, soil inorganic nitrogen content, soil insoluble organic carbon and nitrogen contents, bacterial gene copy number, and bacterial high-throughput sequencing were measured to quantify the carbon and nitrogen utilization and bacterial communities under different soil types.

Schematic diagram of experimental design.
2.3 Determination of soil physicochemical properties
Soil chemical properties were analyzed by the method of Lu (2000)30. The pH was measured by a water-to-soil ratio of 4:1 (v/w). Soil volume weight and water content were determined via the drying methods. SOC was determined by K2Cr2O7 oxidation method. Soil total nitrogen (TN) was measured by the Kjeldahl method of Lu (2000). Soil ammonium nitrogen (NH4+) and nitrate nitrogen (NO3−) were determined by a continuous flow analyzer (AA3, Germany). Soil 13C and 15N abundances were measured by an isotope mass spectrometer (Germany). Soil microbial respiration was measured by closed incubation and CO2 emissions were determined by gas chromatography (Wang et al., 2003).
2.4 Microbial analyses
The complete soil microbial genome was extracted using the FastDNA® Spin Kit for Soil (MPBiomedicals). The DNA was dissolved in 100 µL of DES buffer, and the detailed procedures described in the kit instructions were followed. PCR amplification was used universal primers of bacteria (515 F/907R), and sequencing analysis was used by the Illumina NovaSeq platform. All raw sequences can be found at NCBI under accession numbers PRJNA906297, PRJNA906361, PRJNA906365 and PRJNA906442.
2.5 Statistical analyses
Data record and analysis was used by the Microsoft Excel 2013, and used the SPSS 26.0 to statistics and analysis the data. And all graphs were generated in Origin 2023 soft in this paper. Principal coordinate analysis (PCoA) based on Bray-Curtis distance and permutation multivariate analysis of variance were performed in the R package vegan. In the same package, redundancy analysis (RDA) was used to analysis the correlation between chemical properties and bacterial communities. Network analysis was to reduce complexity, only the OTUs with an average relative abundance > 0.01 were retained to construct the network. The visualization of the correlation network and structural equation model was achieved using R (4.2.3).
3. Results
3.1 Microbial respiration and changes in inorganic nitrogen in the different soil types
The results showed that after 42 days of microcosm incubation, the addition of glucose and ammonium nitrate (GN) increased CO2 emissions more than the addition of glucose alone (G), but this was not true in the banana garden soil (Fig. 3c). In contrast, the addition of banana root and ammonium nitrate (BRN) significantly reduced CO2 emissions, more than the addition of banana root alone (BR). The G and GN treatments resulted in significantly higher CO2 emissions than the other treatments in all four soil types. The CK treatment had the lowest emissions, and the respiration of the banana straw and root addition treatments was lower than that of the glucose addition treatments in NS. Compared with G treatment, it was reduced by 51.87% and 62.12%, which indicates that microorganisms can quickly use glucose and ammonium nitrate and emit large amounts of CO2 in different soil types, but the conversion of organic carbon is relatively poor with banana straw and roots (Fig. 3a, b, c, d). The inorganic nitrogen content in the CK treatment was almost unchanged, and all treatments showed a greater concentration of NO3− than NH4+ at the end. The inorganic nitrogen concentration in the addition of glucose treatment (G) showed a greater concentration of NH4+ than NO3−; but at the same time, the ammonium nitrogen concentration increased significantly with the addition of glucose and ammonium nitrate (GN), and was significantly higher than the nitrate nitrogen concentration (P < 0.05). In all four soil types, the inorganic N contents in the banana straw (BS) and banana root (BR) treatments were significantly lower than those in the banana straw + ammonium nitrate (BSN) and banana root + ammonium nitrate (BRN) treatments. The ammonium N concentration was higher than the nitrate N concentration in the BSN treatment, decreased by 28.44%, 13.25%, 14.7% and 36.43%, respectively, but the opposite result was observed in the BRN treatment. In all four soil types, the treatments showed that nitrate-nitrogen was rapidly consumed after the addition of ammonium nitrate. The GN treatment absorbed and utilized the most nitrate-nitrogen, indicating that microbial utilization of easily-absorbed organic carbon (glucose) requires the participation of inorganic nitrogen, and nitrate-nitrogen is especially effective (Fig. 3e, f, g, h).

Microbial respiration (Carbon dioxide emissions) under the NS (a), HS (b), DS (c) and PS (d); Inorganic N (NH4+, NO3−) content under the NS (e), HS (f), DS (g) and PS (h) after 42 days of cultivation. Different letters represent significant correlations in treatments (P < 0.05). Treatments: CK: no glucose or NH4NO3; G: glucose; GN: glucose and NH4NO3; BS: banana straw; BSN: banana straw and NH4NO3; BR: banana root; BRN: banana root and NH4NO3.
3.2 Insoluble organic carbon and nitrogen content in different soil types
The results showed that after 42 days of incubation, the content of insoluble organic carbon in the CK treatment measured from lowest to highest in PS > DS > HS > NS. In different soil types, the content of insoluble organic carbon in each treatment showed an increasing trend with the addition of organic carbon and inorganic nitrogen. Thus the content of insoluble organic carbon in the BR and BRN treatments was higher than in other treatments in different soil types, and increased by 30.16% and 26.91%, respectively (Fig. 4a, b, c, d). Compared with the control, the addition of glucose (G) reduced the insoluble organic nitrogen in NS, HS and PS soils, and decreased by 9.67%, 3.08% and 2.22%, respectively (Fig. 4e, f, g, h), while the addition of banana straw or root increased the insoluble organic nitrogen in all soils except NS. The insoluble organic nitrogen content was higher in paddy soils than in other soil types, with the GN treatment having the lowest insoluble organic nitrogen content (Fig. 4h). At the same time, the C/N ratio also showed a significant upward trend in all different soil types. The C/N ratio was significantly higher in the BR and BRN treatments than in the others, which indicates that the organic carbon provided by banana roots was more easily converted into soil organic carbon, so it significantly improved the soil C/N ratio, increased soil disease resistance, and fostered soil health (Fig. 4i, j, k, l).

Insoluble organic carbon in NS (a), HS (b), DS (c) and PS (d); Nitrogen content in NS (e), HS (f), DS (g) and PS (h); C/N Ratio in NS (i), HS (j), DS (k) and PS (l). Different letters represent significant correlations in treatments (P < 0.05). Treatments: CK: no glucose or NH4NO3; G: glucose; GN: glucose and NH4NO3; BS: banana straw; BSN: banana straw and NH4NO3; BR: banana root; BRN: banana root and NH4NO3.
3.3 15N abundance in different soil types
The study found that by adding 15N2, the 15N abundance in NS, HS, DS and PS was significantly higher in G treatments than in other treatments (Fig. 5a, b, c, d), with the greatest 15N in NS, which increased by 436.38%, 303.22%, 244.92% and 316.21%, respectively. The addition of other types of carbon (banana straw or root) and nitrogen did not have a significant effect on the conversion of 15N2, which indicates that the addition of glucose can increase the uptake and transformation of N2 by soil microorganisms and can improve the transformation efficiency of N.

15N abundance in NS (a), HS (b), DS (c) and PS (d). Different letters represent significant correlations in treatments (P < 0.05). Treatments: CK: no glucose or NH4NO3; G: glucose; GN: glucose and NH4NO3; BS: banana straw; BSN: banana straw and NH4NO3; BR: banana root; BRN: banana root and NH4NO3.
3.4 Copy number of 16 S gene in different soil types
In general, the bacterial copy number and nifH copy number of BR were significantly higher than those of other treatments in natural forest soil and increased by 74.04%, 82.64%, 69.66%, 94.36%, 48.76% and 56.23%, respectively compared with the control (Fig. 6a, e). In healthy banana soil and rice soil, the bacterial copy number and nifH copy number of BS were significantly higher than those of other treatments (Fig. 6b, d, f, h). In the diseased banana soil, the bacterial copy number and nifH copy number of BSN were significantly higher than those of other treatments (Fig. 6c, g).

Copy number of 16 S gene in NS (a), HS (b), DS (c) and PS (d) and Copy number of nifH gene in NS (e), HS (f), DS (g) and PS (h); Different letters represent significant correlations in treatments (P < 0.05). Treatments: CK: no glucose or NH4NO3; G: glucose; GN: glucose and NH4NO3; BS: banana straw; BSN: banana straw and NH4NO3; BR: banana root; BRN: banana root and NH4NO3.
3.5 Diversity of bacterial communities
In natural forest soils, the bacterial diversity of BR and BRN treatments was significantly higher than that of other treatments. In healthy banana soil and paddy soil, each treatment was significantly lower than that of CK treatment. However, in the diseased banana soil, Chao 1 and ACE were significantly lower than CK treatments, but the BR treatments Shannon and Simpson were significantly higher than those of other treatments (Table 2).
3.6 Community structure of bacteria in different soil types
Firmicutes, Actinobacteria, Proteobacteria, Acidobacteria, and Chloroflexi were the dominant microbial phylum (relative abundance > 1%) in the different soil types. The composition of bacteria varied in each treatment and soil, but in general, the relative abundance of Firmicutes was significantly higher in the G and GN treatments than in the others, with respective values of 83.73% and 76.56% in NS, 66.39% and 54.87% in HS, 67.19% and 60.13% in DS and 70.36% and 65.67% in PS. In HS, DS and PS, the relative abundance of Actinobacteria was higher in the BS, BSN, BR, and BRN treatments than in the others, but it was lower in NS. This indicates that the addition of glucose and ammonium nitrate increased the relative abundance of Firmicutes, independent of soil type, but the addition of organic carbon (banana straw or roots), which is difficult to absorb, increased the relative abundance of Actinobacteria in all treatments except NS. The rise in relative abundance of Actinobacteria in the treatments adding banana straw or roots in NS is more appropriately attributed to soil type (Fig. 7a, b, c, d). The PCoA of bacterial community structure in the different soil types revealed that the first two axes explained 99.39, 97.31, 96.26, and 99.34 of the total variances, respectively (Fig. 7e, f, g, h). In addition, the community structure of bacteria in the CK treatment differed significantly from the other treatments (G, GN, BS, BSN, BR, BRN). In particular, the G and GN treatments also were distinct from other treatments. These results indicate that differences in organic C and/or inorganic N across different soil types can change the bacterial community structure.

Community structure of bacteria in NS (a), HS (b), DS (c) and PS (d); The PCoA of bacterial community structure in NS (e), HS (f), DS (g) and PS (h). Different letters represent significant correlations in treatments (P < 0.05). Treatments: CK: no glucose or NH4NO3; G: glucose; GN: glucose and NH4NO3; BS: banana straw; BSN: banana straw and NH4NO3; BR: banana root; BRN: banana root and NH4NO3.
3.7 Relationships between bacterial community composition and soil properties
The correlations between soil properties and bacterial community structure were assessed using RDA. Overall, the first two axes explained 83.40%, 84.56%, 78.07%, and 81.82% of the total variability in the community structures of bacteria in NS, HS, DS and PS, respectively. Among the multiple soil variables, nutrient parameters primarily influenced bacterial community structure (Fig. 8). Nitrate nitrogen was strongly correlated with bacterial community composition in natural forest soils (Fig. 8a), soil insoluble organic carbon and inorganic nitrogen were strongly correlated with bacterial community composition in healthy plantain and paddy soils (Fig. 8b, d), and insoluble organic nitrogen was least correlated with bacterial community composition in diseased plantain soils (Fig. 8c). In NS, NH4+ significantly affected bacterial colony structure and correlated negatively with bacterial colonies, but soil ION content correlated positively with bacterial colonies, and IOC was significantly and positively correlated with both Actinobacteria and Cyanobacteria (Fig. 8e). In HS and DS, IOC content was significantly and positively correlated with both Actinobacteria and Cyanobacteria (Fig. 8f, g), and in PS, IOC content was significantly and positively correlated with Actinobacteria (Fig. 8h). This indicates that soil IOC content could positively regulate bacterial Actinobacteria and Cyanobacteria in different soil types.

RDA analysis in NS (a), HS (b), DS (c) and PS (d). Relationships between bacterial community composition, and soil properties in NS (e), HS (f), DS (g) and PS (h). D Different letters represent significant correlations in treatments (P < 0.05). Treatments: CK: no glucose or NH4NO3; G: glucose; GN: glucose and NH4NO3; BS: banana straw; BSN: banana straw and NH4NO3; BR: banana root; BRN: banana root and NH4NO3.
3.8 Network analysis between species
We found that the complexity of species networks showed obvious differences across different treatments. Taking each treatment in natural forest soil as an example, the network was most complex in the CK treatment, and the complexity decreased significantly in the G treatment. With the addition of ammonium nitrate concurrently with glucose (GN treatment), the network complexity of bacteria increased significantly. This indicates that bacterial growth requires carbon and nitrogen sources at the same time, and a single carbon source will not increase species richness, but may instead reduce it. At the same time, the banana straw and root addition treatments (BS, BR) showed a decrease in species network complexity compared to the CK treatment, but the supplementation of nitrogen sources improved this phenomenon (Fig. 9).

Network analysis between species in NS (a), HS (b), DS (c) and PS (d). Treatments: CK: no glucose or NH4NO3; G: glucose; GN: glucose and NH4NO3; BS: banana straw; BSN: banana straw and NH4NO3; BR: banana root; BRN: banana root and NH4NO3.
3.9 Correlation analysis of different types of soil properties
We used the structural equation model to analyze the effects of carbon and nitrogen addition on the physicochemical properties and bacterial community characteristics of different soil types, and we found that the effects of carbon and nitrogen addition on soil physical and chemical properties and on bacterial community characteristics differed greatly across different soil types, though the effects were not significant in forest soil and banana soil (Fig. 10a, b, c). In paddy soil, however, carbon and nitrogen addition was significantly negatively correlated with bacterial community structure (Fig. 10d). In all four types of soil, nitrogen addition correlated positively with NH4+ soil content, and carbon addition was significantly positively correlated with poorly-soluble organic carbon content and C/N in all soils except forest soil (Fig. 10). At the same time, the C/N of forest soil was significantly positively correlated with bacterial community structure, and the Chao 1 index was significantly negatively correlated with bacterial community structure (Fig. 10a). NO3− and the Chao 1 index of healthy banana soil were significantly negatively correlated with bacterial community structure (Fig. 10b). The NH4+ and C/N of diseased banana soil were significantly negatively correlated with bacterial community structure (Fig. 10c). NH4+ in paddy soil was significantly positively correlated with bacterial community structure, but the Chao 1 index and BCN were significantly negatively correlated with bacterial community structure (Fig. 10d). Random forest analysis showed that Shannon, Chao1 and CO2 were important factors for predicting bacterial community structure in natural forest soil (Fig. 10e), CO2 and Shannon were important factors for predicting bacterial number in healthy banana and rice soil (Fig. 10f, h), and IOC, CO2 and Shannon were important factors for predicting bacterial number in diseased banana soil (Fig. 10g).

Relationship between bacterial community composition and soil properties under different soil types. Structural equation model in NS (a), HS (b), DS (c) and PS (d). Random forest analysis in NS (e), HS (f), DS (g) and PS (h). CO2: carbon dioxide emissions; IOC: Insoluble organic carbon; ION: Insoluble organic nitrogen; NO3−: nitrate nitrogen; NH4+: ammonium nitrogen; C/N: IOC/ION; BCN: Bacterial copy number.
4. Discussion
4.1 Underground plant residues contributed more to soil organic carbon than the above-ground
Land management was determined by different land utilization types, which thereby impacted soil physical and chemical properties31,32. Soil basic chemical properties indicate the levels of soil nutrients, and the analysis of variance for soil characteristics found that there are significantly different under different land utilization patterns. There are also differences in the use of fertilizers under different soil utilization patterns. Bio-fertilizers, chemical fertilizers and crop residues are well-known additives for preserving and enhancing soil quality33 by preserving or enhancing soil organic carbon34. Thus, these additives can improve microbial activity and function in the field and improve biomass and C sequestration capacity of a plant35. SOM plays a significant role in ecosystem function and soil fertility, and it determines the nutrient sequestration capacity of the soil36. This study showed that the addition of organic carbon (glucose or banana straw) and inorganic nitrogen together increased the soil insoluble organic carbon content and improved the soil organic carbon content. It is speculated that the main reason for this is that the input of carbon and nitrogen nutrients promotes the reproduction of soil microorganisms, which leads to the increase of soil insoluble carbon and nitrogen (mainly microbial biomass carbon and nitrogen), which is generally consistent with results of previous research. Many studies have found that below-ground carbon inputs (root and root exudation) form stable carbon more efficiently than above-ground litter carbon inputs37,38,39. In this study, we found that insoluble organic carbon was higher in the banana root treatment than in the banana straw treatment. This indicates that the underground plant residues contributed more to soil organic carbon than did the above-ground part. This is mainly closely related to the ability of microorganisms to decompose, and it may be that the ratio of carbon to nitrogen in plant roots is more suitable for decomposition by microorganisms, which was also basically consistent with previous studies.
4.2 Efforts of carbon and nitrogen addition on microbial respiration and nitrogen fixation
The average CO2 production rate is typically used as an effective indicator of soil microbial activity, and it also reveals the ability of carbon sources to be utilized by soil microbial communities40, and the higher residue carbon inputs may lead to high rates of carbon sequestration41. It is well known that soil respiration demonstrate biological activity and decomposition of organic residues42,43. Higher respiration rate of microbes in soil may show great microbial transformation efficiency in the environment44. The present study showed that organic carbon and inorganic nitrogen addition increased microbial respiration and improved the conversion efficiency of nutrients, which is consistent with results of previous research. We also found that the same phenomenon occurred in different types of soil microorganisms, indicating that soil types did not affect the assimilation effect of microorganisms on carbon and nitrogen. And the nitrogen fixation capacity of different soil types had a significant decrease after the nitrogen addition. It can be evidenced by a decrease in the abundance of15N. The reason may be that excessive soluble nitrogen addition has a toxic effect on nitrogen-fixing microorganisms, resulting in a decrease in the number of nitrogen-fixing microorganisms, which in turn reduces the overall biological nitrogen fixation capacity45.
4.3 Efforts of carbon and nitrogen addition on bacterial community composition
Microbial diversity is considered to be an important component in global carbon cycle, but bacterial communities composition are often overlooked in stoichiometric nutrient cycling study46,47. Some studies have demonstrated that bacteria in soil could increase growth of plant, nutrient allocation by means of nutrient element turnover in soil, and changes in bacterial community structure and diversity also alter soil quality, pH and other environmental indicators48,49. The composition of soil microbial communities in systems where fertilizer is applied is largely affected by soil chemical properties, which has been reported by many investigators50,51, and factors in environment such as pH, SOM and AP have the potential to affect soil microbial communities in different ecological system52,53, and the bacterial community structure was remarkably correlated with soil organic carbon and total nitrogen54. In this study, nitrate nitrogen was strongly correlated with bacterial community composition in natural forest soils, insoluble organic carbon and inorganic nitrogen in soil were strongly correlated with bacterial community composition in healthy plantain and paddy soils, and insoluble organic nitrogen was least correlated with bacterial community composition in diseased plantain soils, indicating that there were obvious differences in the responses of different types of soil microorganisms to carbon and nitrogen, and the bacterial communities in forest soil were more sensitive to nitrogen. Additionally, the Actinomycetales order is believed a main contributor to carbohydrate (CHO) and amino acid (AA) metabolism under various conditions55,56. We found that the addition of glucose and ammonium nitrate increased the relative abundance of Firmicutes, independent of soil type, but the addition of difficult-to-absorb organic carbon (banana straw or roots) increased the relative abundance of Actinomycetes in all soils except NS. In all ecosystems, soil microbes play an increasingly greater role in decomposition of SOM, N cycle and providing sufficient nutrients for plants. Thus microbes act as a sensitive indicator for forecasting soil biological conditions and recommended agricultural practices in soil ecosystem57. The soil microbial community is closely linked with agricultural production, including plant straw, organic material, and crop management, etc58. Soil organic carbon and soil eco-environment can be significantly improved through the application of crop residues59. These provide carbon and make the environment hospitable for the proliferation of soil microbes. In our study, it was found that the addition of banana straw or root increased the soil organic carbon content, which is consistent with the results of previous research. Frasier et al. found that soil microbial communities were larger in areas where crop residue treatments were applied, compared to those without60. Soil microbes were better able to uptake C sources in zero-tillage conditions than under conventional tillage61. Diversity and stability of soil microbe communities increased under conditions of long-term no-tillage and organic input practice62. Abundance of soil fungi increased under a management plan of no-tillage and straw mulching63. PCoA differed significantly (p < 0.05) in bacterial communities under different treatments, this is in accordance with the results that documented clear separation of bacterial and fungal community composition between those using straw and those not using it64. One study demonstrated no significant difference among varying regimes of long-term maize straw utilization and fertilization on the bacterial community composition under the phylum level65. According to the PCoA results in our study, the addition of banana straw or root resulted in a bacterial community structure that was significantly distinct from the control. These results indicated that the addition of materials with different C/N ratios would have a great impact on the soil bacterial community, which might cause the formation of a large number of carbon-sequestration microorganisms.
5. Conclusion
With the change of different soil types utilization, glucose addition increased carbon dioxide emissions and enhanced the ability of soil microorganisms nitrogen fixation, but caused the accumulation of ammonium nitrogen, indicating that microorganisms had higher utilization efficiency of soluble carbon and enhanced the biological nitrogen fixation capacity, and nitrogen addition caused the accumulation of ammonium nitrogen, thereby weakening the biological nitrogen fixation capacity. At the same time, glucose significantly increased the Fimicutes phylum and lowered the level of other bacterial phylums, thereby reducing the bacterial network structure, and forest analysis showed that CO2 was an important factor in predicting the bacterial community structure of different soil types, indicating that bacteria at the Fimicutes phylum level preferred glucose, which may also have a negative effect on bacteria at the level of other phylums. In summary, in the agricultural production process, the application of organic carbon should be increased, and at the same time attention should be paid to the management of nitrogen, especially to reduce the application of ammonium nitrogen, to ensure the healthy development of soil microbial community.
Data availability
The datasets generated and/or analysed during the current study are not publicly available due but are available from the corresponding author on reasonable request.
References
Finzi, A. C. et al. Rhizosphere processes are quantitatively important components of terrestrial carbon and nutrient cycles. Global Change Biol. 21, 2082–2094 (2015).
Philippot, L. et al. Going back to the roots: the microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 11, 789–799 (2013).
Ling, N. et al. Variation of rhizosphere bacterial community in watermelon continuous mono-cropping soil by long-term application of a novel bioorganic fertilizer. Microbiol. Res. 169, 570–578 (2014).
Pieterse, C. M. J., de Jonge, R. & Berendsen, R. L. The soil-borne supremacy. Trends Plant. Sci. 21, 171–173 (2016).
Chen, Z. et al. Changes in soil microbial community and organic carbonfractions under short-term straw return in a rice–wheat cropping system. Soil. Tillage Res. 165, 121–127 (2017).
Geisseler, D. & Scow, K. M. Long-term effects of mineral fertilizers on soil microorganisms - a review. Soil. Biol. Biochem. 75, 54–63 (2014).
Fierer, N., Strickland, M. S., Liptzin, D., Bradford, M. A. & Cleveland, C. C. Global patterns in belowground communities. Ecol. Lett. 12(11), 1238–1249 (2009).
Redding, M. R. et al. Soil N availability, rather than N deposition, controls indirect N2O emissions. Soil Biol. Biochem. 95, 288–282 (2016).
Wang, H. et al. Nitrogen addition reduces soil bacterial richness, while phosphorus addition alters community composition in an old-growth N-rich tropical forest in southern China. Soil Biol. Biochem. 127, 22–30 (2018).
Fierer, N. et al. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 6(5), 1007–1017 (2012).
Yan, Y. et al. Sensitivity of soil fungal and bacterial community compositions to nitrogen and phosphorus additions in a temperate meadow. Plant. Soil. 471(1–2), 477–490 (2022).
Wei, C. Z. et al. Nitrogen deposition weakens plant-microbe interactions in grassland ecosystems. Glob. Change Biol. 19(12), 3688–3697 (2013).
Jobbágy, E. G. & Jackson, R. B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 10, 423–436 (2000).
Romaní, A. M., Fischer, H., Mille-Lindblom, C. & Tranvik, L. J. Interactions of bacteria and fungi on decomposing litter: differential extracellular enzyme activities. Ecology. 87, 2559–2569 (2006).
Witt, C. et al. Crop rotation and residue management effects on carbon sequestration, nitrogen cycling and productivity of irrigated rice systems. Plant. Soil. 225(1), 263–278 (2000).
Song, L. et al. Study on characteristics of decomposing and nutrients releasing of different proportional mixture of rape straw and Chinese milk vetch in rice field. Soils Fertilizers Sci. China. 3, 100–104 (2015).
Feng, X. & Simpson, M. J. Temperature and substrate controls on microbial phospholipid fatty acid composition during incubation of grassland soils contrasting in organic matter quality. Soil Biol. Biochem. 41(4), 804–812 (2009).
Merilä, P. et al. Soil organic matter quality as a link between microbial community structure and vegetation composition along a successional gradient in a boreal forest. Appl. Soil. Ecol. 46(2), 259–267 (2010).
Ma, L. J. et al. Growth and yield of cotton as affected by different straw returning modes with an equivalent carbon input. Field Crops Res. 243, 1–10 (2019).
Yu, C. R. et al. Effects of wheat straw incorporation in cotton wheat double cropping system on nutrient status and growth in cotton. Field Crops Res. 197, 39–51 (2016).
Fierer, N. Embracing the unknown: disentangling the complexities of the soil microbiome. Nat. Rev. Microbiol. 15, 579–590 (2017).
Yin, W. et al. Wheat and maize relay planting with straw covering increases water use efficiency up to 46%. Agron. Sustain. Dev. 35(2), 815–825 (2015).
Tian, J., Pausch, J., Yu, G., Blagodatskaya, E. & Kuzyakov, Y. Aggregate size and glucose level affect priming sources: a three-source– partitioning study. Soil. Biol. Biochem. 97, 199–210 (2016).
Wild, B., Schnecker, J. & Alves, R. J. E. Input of easily available organic C and N stimulates microbial decomposition of soil organic matter in arctic permafrost soil. Soil. Biol. Biochem. 75, 143–151 (2014).
Ollivier, J. et al. Nitrogen turnover in soil and global change. FEMS Microbiol. Ecol. 78(1), 3–16 (2011).
Hou, H. J. et al. Research progress of the molecular ecology on microbiological processes in soil nitrogen cycling. Res. Agricultural Modernization. 35(5), 588–594 (2014).
Orr, C. H. et al. Impacts of organic and conventional crop management on diversity and activity of free-living nitrogen fixing bacteria and total bacteria are subsidiary to temporal effects. PLoS One. 7(12), 483–496 (2012).
Berthrong, S. T. et al. Nitrogen fertilization has a stronger effect on soil nitrogen-fixing bacterial communities than elevated atmospheric CO2. Appl. Environ. Microbiol. 80(10), 3103–3112 (2014).
Ogilvie, L. A., Hirsch, P. R. & Johnston, A. W. Bacterial diversity of the broadbalk‘classical’winter wheat experiment in relation to long-term fertilizer inputs. Microb. Ecol. 56(3), 525–537 (2008).
LU RK. Methods of Soil Agrochemical Analysis [M] 146–195 (China Agriculture Scientech, 2000).
Wolinska, A. et al. Microbial biodiversity in arable soils is affected by agricultural practices. Int. Agrophys. 31(2), 259 (2017).
Batlle-aguilar, J., Brovelli, A., Porporato, A. M. & Barry, D. A. Modelling soil carbon and nitrogen cycles during land use change. A review. Agron. Sustain. Dev. 31(2), 251 (2011).
Bhattacharyya, P., Pal, R., Chakraborty, A. & Chakrabarti, K. Microbial biomass and activity in a laterite soil amended with municipal solid waste compost. J. Agron. Crops Sci. 187, 207–211 (2001).
Li, L. J., Zhu-Barker, X., Ye, R., Doane, T. A. & Horwath, W. R. Soil microbial biomass size and soil carbon influence the priming effect from carbon inputs depending on nitrogen availability. Soil. Biol. Biochem. 119, 41–49 (2018).
Jin, X. et al. Enhanced conversion of newlyadded maize straw to soil microbial biomass C under plastic film mulching and organic manure management. Geoderma. 313, 154–162 (2018).
Dutta, S., Pal, R., Chakraborty, A. & Chakrabarti, K. Influence of integrated plant nutrient supply system on soil quality restoration in a red and laterite soil: Einfluss integrierter pflanzennährstoff versorgung auf die Wiederherstellun Der bodenqualität Von Rotem und laterit boden. Arch. Agron. Soil. Sci. 49, 631–637 (2003).
Cotrufo, M. F. et al. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat. Geosci. 8, 776–779 (2015).
Sokol, N. W. & Bradford, M. A. Microbial formation of stable soil carbon is more efficient from below-ground than above-ground input. Nat. Geosci. 12, 46–53 (2019a).
Sokol, N. W., Kuebbing, S. E., Karlsen-Ayala, E. & Bradford, M. A. Evidence for the primacy of living root inputs, not root or shoot litter, in forming soil organic carbon. New Phytol. 221, 233–246 (2019b).
Sinsabaugh, R. L., Manzoni, S., Moorhead, D. L. & Richter, A. Carbon use efficiency of microbial communities: stoichiometry, methodology and modelling. Ecol. Lett. 16, 930–939 (2013).
Clapp, C. E., Allmaras, R. R., Layese, M. F., Linden, D. R. & Dowdy, R. H. Soil organic carbon and 13 C abundance as related to tillage, crop residue, and nitrogen fertilization under continuous corn management in Minnesota. Soil. Till Res. 55, 127–142 (2000).
Ferreira, A. C. C., Leite, L. F. C., de Araújo, A. S. F. & Eisenhauer, N. Land-use type effects on soil organic carbon and microbial properties in a semi-arid region of northeast Brazil. Land. Degrad. Dev. 27, 171–178 (2014).
Santos, V. B., Leite, L. F. C., Nunes, L. A. P. L. & Melo, W. J. Soil microbial biomass and organic matter fractions during transition from conventional to organic farming systems. Geoderma. 170, 227–231 (2012).
Islam, K. R. & Weil, R. R. Soil quality indicator properties in mid-atlantic soils as influenced by conservation management. J. Soil. Water Conserv. 55, 69–78 (2000).
Xie, Z. H. Contribution of maize straw nitrogen to soybean nitrogen accumulation in black soil area[D].University of Chinese Academy of Sciences (Northeast Institute of Geography and Agroecology,Chinese Academy of Sciences), (2017).
Graham, E. B. et al. Do we need to understand microbial communities to predict ecosystem function? A comparison of statistical models of nitrogen cycling processes. Soil. Biol. Biochem. 68, 279–282 (2014).
McGuire, K. & Treseder, K. Microbial communities and their relevance for ecosystem models: decomposition as a case study. Soil. Biol. Biochem. 42, 529–535 (2010).
Li, P. et al. Difference in soil bacterial community composition depends on forest type rather than nitrogen and phosphorus additions in tropical montane rainforests. Biol. Fertil. Soils. 55, 313–323 (2019).
Samaddar, S. et al. Long-term phosphorus limitation changes the bacterial community structure and functioning in paddy soils. Appl. Soil. Ecol. 134, 111–115 (2018).
Jiang, S. Q. et al. High-throughput absolute quantification sequencing reveals the effect of different fertilizer applications on bacterial community in a tomato cultivated coastal saline soil. Sci. Total Environ. 687, 601–609 (2019).
Rousk, J. et al. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 4, 1340–1351 (2010).
Liu, J. S. et al. Long-term high-P fertilizer input decreased the total bacterial diversity but not phod-harboring bacteria in wheat rhizosphere soil with available-P deficiency. Soil. Biol. Biochem. 149, 107918 (2020a).
Liu, Z. et al. Shifts in microbial communities and networks are correlated with the soil ionome in a kiwifruit orchard under different fertilization regimes. Appl. Soil. Ecol. 149, 103517 (2020b).
Wu, Z. X. et al. Application of bio-organic fertilizer, not biochar, in degraded red soil improves soil nutrients and plant growth. Rhizosphere. 16, 100264 (2020).
Verastegui, Y. et al.Multisubstrate Isotope Labeling and Metagenomic Analysis of Active Soil Bacterial Communities Vol. 5, e01157–e01114 (mBio, 2014).
De La Cruz-Barrón, M. et al. The Bacterial Community structure and Dynamics of Carbon and Nitrogen when Maize (Zea mays L.) and its neutral detergent Fibre were added to Soil from Zimbabwe with Contrasting Management practices. Microb. Ecol. 73, 135–152 (2016).
Hartmann, M., Frey, B., Mayer, J. & Maeder, P. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 9, 1177–1194 (2015).
Hao, M. M. et al. Shifts in microbial community and carbon sequestration in farmland soil under long-term conservation tillage and straw returning. Appl. Soil. Ecol. 136, 43–54 (2019).
Tang, H. M. et al. Effects of different soil tillage systems on soil carbon management index under double-cropping rice field in southern China. Agron. J. 111, 440–446 (2019).
Frasier, I. et al. High quality residues from cover crops favor changes in microbial community and enhance C and N sequestration. Glob Ecol. Conserv. 6, 242–256 (2016).
Yang, Q., Wang, X. & Shen, Y. Comparison of soil microbial community catabolic diversity between rhizosphere and bulk soil induced by tillage or residue retention. J. Soil. Sci. Plant. Nut. 13, 187–199 (2013).
Wang, Y. et al. Long-term no-tillage and organic input management enhanced the diversity and stability of soil microbial community. Sci. Total Environ. 609, 341–347 (2017).
Wang, H. H. et al. Long-term no-tillage and different residue amounts alter soil microbial community composition and increase the risk of maize root rot in northeast China. Soil. Till Res. 196, 104452 (2020).
Bei, S. K. et al. Response of the soil microbial community to different fertilizer inputs in a wheat-maize rotation on a calcareous soil. Agr Ecosyst. Environ. 260, 58–69 (2018).
Xia, X. Y. et al. Effects of tillage managements and maize straw returning on soil microbiome using 16S rDNA sequencing. J. Integr. Plant. Biol. 61(6), 765–777 (2019).
Acknowledgements
We would like to thank Dr. Daniel Petticord for helping English language of the manuscript. The research was funded by Project of Sanya Yazhou Bay Science and Technology City (SCKJ-JYRC-2022-94), Strategic Priority Research Program of Chinese Academy of Sciences (XDA28020203), Postgraduate Innovation Research Project of Hainan Province (Qhyb2022-67), and PhD Scientific Research and Innovation Foundation of Sanya Yazhou Bay Science and Technology City (HSPHDSRF-2023-12-008).
Author information
Authors and Affiliations
Contributions
ZJ, YR and LM contributed to conceive and design the experiment. CH, KL contributed to perform the experiment and writes this paper. The authors have confirmed the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
He, C., Li, K., Meng, L. et al. Glucose addition in natural forest soils has higher biological nitrogen fixation capacity than other types of soils. Sci Rep 14, 23909 (2024). https://doi.org/10.1038/s41598-024-72796-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-72796-z
