
Abstract
Vertical stratification of forest plays important roles in the local material balance and in maintaining forest health by distributing and redistributing precipitation materials through adsorption, fixation, and release. Differences in runoff nutrient concentrations among vertical layers are closely related to vertical stratification (factors such as the trunk, canopy, forest litter, and soil physical and chemical properties). Long-term forest observations revealed significant spatial differences in Pinus massoniana (Pinus massoniana Lamb.) forests in the Three Gorges Reservoir area. Pinus massoniana forests on downslopes were characterized by a dense canopy, green needles, and rich forest vegetation, while those on upslopes were characterized by low vegetation cover, dead trees, and decreases in the tree height, diameter at breast height, and volume per plant with increasing slope. By analyzing the soil at different sites, we found that the pH of the forest land soil differed significantly among different slope positions. Soil on upper slopes was significantly more acidic than soil on lower slopes, indicating that acidic substances were intercepted by filtration through the broad litter layer and the soil surface layer. This filtration process resulted in a normal rhizosphere environment suitable for the absorption of nutrients by vegetation on the lower slopes. In this way, downhill sites provided a good microenvironment for the growth of Pinus massoniana and other vegetation. Our results show that direct contact between needles and acid rain was not the main cause of root death. Instead, the redistribution of rainfall substances by forest spatial stratification caused changes in the soil microenvironment, which inhibited the absorption of nutrients by the roots of Pinus massoniana and the growth of understory plants in Pinus massoniana forests on upper slopes. These findings emphasize that increasing land cover with forests with vertical structural stratification plays an important role in woodland material redistribution and forest conservation.
Similar content being viewed by others
Introduction
The vertical structure of forests plays a crucial role in the local material balance by partitioning acid rainfall by adsorbing, fixing, and releasing precipitation substances1. This partitioning regulates the distribution of acid rainfall at different positions within the forest; and therefore, it strongly affects the growth of various kinds of vegetation in woodland2. Although many studies have focused on rainfall distribution via the vertical structure of forests, most have concentrated on the distribution of rainfall at a single level (canopy, understory, or ground)3,4. There is still a knowledge gap regarding the above-ground and underground distribution of acid rainfall in a complete forest ecosystem. Furthermore, it is not known which level of acid-rainfall distribution within the forest vertical structure has the greatest influence on forest health. Therefore, studies on acid rainfall distribution as affected by the vertical structure of forests are important to understand how acid rainfall redistribution affects material distribution at different levels and biodiversity in the forest ecosystem.
There are large differences in runoff nutrient concentrations among different layers in the vertical structure of the forest, and this is usually related to multiple factors, including trunk, crown, and forest litter characteristics and soil physical and chemical properties5,6. Under natural conditions, these factors are covariant, so it is difficult to determine their influence on forests by single-level studies. Some studies have indicated that the concentration of acid rainfall at different locations depends on the runoff rate and the original concentration of acid in water, which are influenced by the canopy, trunk, and litter, and by soil adsorption and desorption7,8. It was found that the concentration of substances in runoff water passing through tree trunk, litter layer and soil was significantly different from that before entering forest land9,10. As the third action surface of atmospheric precipitation into the forest ecosystem after the canopy and trunk layers, the forest litter layer has a strong ability to leach and adsorb various materials in rainfall11,12,13. Some studies have shown that the total nitrogen (N) concentration in runoff decreases significantly with the increase in litter coverage14,15. Because of the interaction between runoff and the soil layer, many substances in runoff are adsorbed and immobilized by the soil layer and associated soil microbial biomass. Consequently, a large number of runoff substances are held in the litter layer, slowing down their migration and distribution into the forest land16,17.
As the most widely distributed coniferous pioneer afforestation species in southern China, Pinus massoniana (Pinus massoniana) plays an important role in the formation of regional ecosystem stability18,19. This is because of its excellent characteristics, including its resistance to drought and poor soils, and its strong adaptability. Pinus massoniana forests are widely distributed in Sichuan, Chongqing, Guizhou, and another 11 provinces in China, with a total area of 1.13 × 107 hm2 and a storage volume of 3.41 × 108 m3. However, as trees have been affected by acid rainfall, large areas of Pinus massoniana forests are in decline, and it has been difficult to restore normal growth. In terms of the cause of Pinus massoniana forest decline, some researchers claim that acid rain is mainly affecting the canopy. It has been proposed that direct contact between acid rainfall and the needles causes serious damage to leaf surface structures (e.g. the cuticle) and selective permeable membranes20. The damage impairs normal metabolic processes such as photosynthesis and material transport, which slow growth and metabolism, ultimately leading to the death of Pinus massoniana trees21. Other researchers believe that acid rainfall indirectly affects Pinus massoniana by causing soil acidification22. The accumulation of acidic compounds in the soil increases the content of active aluminum, and this inhibits the division and elongation of root cells. Aluminum can also combine with root cell membrane proteins, thereby causing cell membrane damage23,24. The portion of aluminum that enters the root cells directly affects cellular metabolism by inhibiting the absorption of nutrients such as calcium, phosphorus, and potassium25. At present, it is still unknown which locations within the vertical structure of Pinus massoniana forests are most severely affected by acid rainfall. Few or no field studies have explored this topic, especially in acid-affected mature Pinus massoniana forests in rapidly developing areas such as Southwest China.
Long-term observations have revealed significant differences in canopy density, conifer color, and biodiversity within the artificially cultivated Pinus massoniana forests in The Three Gorges Reservoir area, particularly among different slope positions (Fig. 1A–C). Specifically, Pinus massoniana forests in lower slope positions are characterized by a dense canopy, green needles, and abundant and diverse forest vegetation (Fig. 1B). If there are large differences in forest vegetation under the same site conditions, they are likely due to differences in material exchange between trees and the environment, such as the nutrient absorption efficiency of vegetation roots and the nutrient absorption environment of the roots26,27. The nutrient-uptake sites of forest vegetation are extremely sensitive under the background of acid deposition27,28,29. Based on the results of previous studies, we tested the following hypothesis in this study: Acid rain more strongly affects the roots of Pinus massoniana than the needles. If this hypothesis is confirmed, then the soil microenvironment is the main factor causing Pinus massoniana forest decline, rather than direct contact between acid rain and the needles.

Vegetation and forest features of Pinus massoniana forest land (LS Low slope, MS Middle slope, US upper slope).
To rigorously test this hypothesis, we conducted a comprehensive study that encompassed the systematic collection and scientific analysis of water, soil and vegetation samples from diverse slope positions within Pinus massoniana forests. Our objective was to assess the specific impact of acid rain on both the roots and needles of these trees, and to establish any potential causal linkages between soil chemistry, vegetation health, and biodiversity. By elucidating the underlying mechanisms of the effects of acid rain on Pinus massonianas, we aim to gain a deeper understanding of the broader ecological implications for forest ecosystems in this region, and potentially, develop targeted and effective strategies for their preservation and restoration.
Materials and methods
Study site
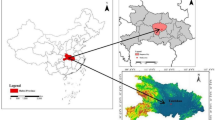
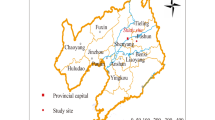
This research was conducted in the eastern Sichuan parallel ridge and valley area (Chongqing region; Fig. 2A, B,E) in the central area of the Three Gorges Reservoir region (105°45’37.84"E, 30°45’37.84” N; 186–274 m.a.s.l.; slope 15–18%) (Fig. 2C). The dominant soil in this area is purplish soil that basically maintains the physical and chemical properties of the parent material, but with a lower organic matter holding capacity (Table 1). The hilly mountain landscape consists of indigenous vegetation and anthropogenic vegetation (especially Pinus massoniana) (Fig. 2F–H).

Includes several parts: (A) shows the geographical ___location of the Three Gorges Reservoir area; (B) displays the ___location of the study area; (C) illustrates the structure and composition of the forestland runoff monitoring plot; (D) depicts a stemflow interceptor; (E) depicts the specific ___location of the forest monitoring plot in Chenjiagou sub-watershed; (F) shows the forest land on the upper slope of the monitoring plot; (G) illustrates the forest land on the mid-slope of the monitoring plot; (H) displays the forest land on the lower slope of the monitoring plot.
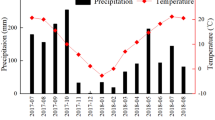
The study area was in the humid subtropical monsoon zone, which is characterized by warm temperatures and high moisture levels, with dry winters and humid summers. The mean annual precipitation was 708.9 mm, ranging 1110 to 1350 mm (2001–2020 dataset), according to long-term observations at the field monitoring station of the Institute of Mountain Hazards and Environment, CAS (Wanzhou Experimental Station). The majority (85%) of rainfall events were concentrated in the warmest months, i.e., May to October (Fig. 3).

Size classes of precipitation events. (A) Average rainfall in mm per month and (B) distribution of precipitation events by size classes in the study area.
Among the rainfall events, 75.88% were small events (< 10 mm) and accounted for 9.38% of the rainfall inputs; whereas 4.65% were large events (from 30 to 50 mm) and represented 71.44% of the rainfall input (Fig. 3B). The mean annual temperature from 2002 to 2020 was 16.5℃ with a minimum and maximum of 10.2 °C and 22.8 °C, respectively (data obtained at the meteorological site of Wanzhou Experimental Station). These conditions were similar to those in the native distribution range of Pinus massoniana (i.e., evergreen coniferous forests of Jiangsu (Liu he, Yi zheng) and Anhui (Huai River Basin, south of Ta-pieh Mountains) in Eastern China).
The Pinus massoniana forests located within the study area were initially planted with the primary objective of mitigating soil erosion in the hilly terrain of eastern Sichuan. Consequently, subsequent to planting, these forests were permitted to develop naturally, without any human intervention. Despite the preponderance of Pinus massoniana trees within the study area’s forests, variations are evident among distinct slope positions (Table 1). Specifically, the upper slope is characterized predominantly by pure stands of Pinus massoniana, whereas the middle slope features Pinus massoniana trees constituting the upper canopy layer, with fern plants dominating the understory vegetation. Conversely, the lower slope typically showcases a mixed forest composition, including Pinus massoniana alongside various broad-leaved tree species, such as Schima superba, Castanopsis eyrei, Castanopsis sclerophylla, Cyclobalanopsis glauca (evergreen broad-leaved species), and Liquidambar formosana, Toxicodendron vernicifluum, and an additional occurrence of Cyclobalanopsis glauca (deciduous broad-leaved species). The herbaceous layer beneath these trees is primarily composed of fern plants and Dicranopteris linearis.
Experimental design and field measurements
Forest stand runoff plot
We collected monitoring data at different slope positions in the forest stand, with measurements taken at a site with a natural concave slope (12 hm2) to monitor the dynamic changes in rainfall. A concrete barrier with a drainage hole was constructed in the lower catchment area of the plot. A drainage pipe was fitted in the hole to direct the runoff into the catchment basin. A runoff device consisting of a diverter groove and a water level monitor was installed below the catchment. The runoff device was triggered whenever runoff was generated, and was used to create flow-producing events (Fig. 2C, H). Every time rainfall triggered the runoff device, the water flowing out of the plot and sink into the catchment was collected for OW(outflow of woodland) data analysis.
Stemflow interceptor
In the forest plot, we chose six Pinus massoniana trees with uniform growth and diameter to install stem interceptors. Each interceptor consisted of a shielding device, a runoff-collecting unit, a drainage-tube, and a water collection manifold. This design ensured that leaf litter could not block runoff collection, thereby allowing for accurate measurements (Fig. 2D). The shielding device was an inverted funnel (20-cm wide) made of polyethylene plastic, which was wrapped around the trunk at a 30° angle (the length depended on the diameter of the trunk). This device prevented needles from falling into the collector. In the lower part of the shielding device, the runoff collection part was connected to a drainage tube. The runoff collected by the collector flowed through the tube into a 20-L graduated water tank on the forest floor. The interceptor was set at a height of about 1.7 m from the base of the trunk. Stem throttling water samples were collected after each flow-producing rainfall. After each rainfall event that generates runoff, collect stem-intercepted water samples for stem interception (SC) analysis.
Rainfall partitioning measurements
Rainwater collectors are installed at the canopy openings in the forest to collect into the forest(IF) rainfall. We collected rainfall partitioning data during 24 rainfall events from 2018 to 2020. During the events that triggered reservoir runoff, we collected samples from all of the triggered runoff meters. We measured rainfall pH using a glass electrode, and nitrate-N, nitrite-N, phosphorus, and dissolved organic carbon (DOC) using an AA3 automatic continuous flow analyzer (SEAL AutoAnalyzer 3, SEAL Analytical GmbH company, Germany)(Table 2). Total nitrogen (TN) was determined by Flow injection chemical analyzer(CFA) and N-(1-naphthyl) ethylene diamine dihydrochloride spectrophotometry. Ammonium-N was calculated using data from the AA3 automatic continuous flow analyzer, with CFA and salicylic acid spectrophotometry. We calculated the ρ mass concentration of TN (mg/L) using Eq. (1):
where y is the measured signal value (peak height), a is the intercept of the standard curve equation, b is the slope of the standard curve equation, and f is the dilution factor.
Assessments of forest vegetation characteristics and tree health
We investigated the vegetation types and quantity in forest plots using the 9-point method (W point method). We recorded weeds (species and quantity) in each quadrat (each quadrat 1 m2,1 m×1 m). We calculated the Shannon-Wiener index as an indicator of vegetation diversity in different positions of woodland, as shown in Eqs. (2, 3):
where N is the total number of plants in the quadrat, ni is the number of individuals of the i species, and SW is the Shannon-Wiener index.
The forest disease index is an objective indicator that reflects the severity of disease. Its calculation typically involves the hierarchical statistics of diseased trees, specifically using the formula:
where DI represents the disease index, xi represents the value of disease severity, and yi represents the number of diseased leaves corresponding to the disease severity represented by xi.
The calculation formula for individual tree volume (also known as single-tree volume) typically relies on the tree’s diameter at breast height (dbh, measured at 1.3 m above ground), tree height, and a form factor that accounts for the influence of trunk shape. The formula is expressed as:
where V represents the individual tree volume(IndvVolm), g1.3 is the basal area at breast height, D represents the diameter of the tree trunk measured at a height of 1.3 m from the ground, H stands for tree height, and 0.42 is generally recognized as the average experimental form factor for neutral coniferous species, which is derived through experimental measurements or empirical formulas.
Soil measurements
Soil organic matter (SOM) was determined by the potassium dichromate method. Soil pH was determined by potentiometry. The TN content in soil was determined by the potassium dichromate digestion method. Total phosphorus (TP) was determined by the molybdenum-antimony colorimetric method. Total potassium (TK) and available potassium (Olsen-K) were determined by flame photometry. Alkali-hydrolyzed-N (alkaline-N) was determined using the diffusion absorption method. Alkali-hydrolyzale nitrogen (AK) was determined by the hydrochloric acid-ammonium fluoride method. The laser particle size method was used to determine the proportions of soil particles of different sizes using an Eyetech-combo particle size analyzer (Rubotherm, Bochum, Germany).
Soil pH value determination
Soil pH is chemically defined as the negative log of H+ ion activity in the soil solution. In this study, pH was measured as water-leached pH and salt-leached pH. Water leaching represents the active acids in soils. Salt leaching involves leaching soil with 1 M KCl solution, and generally reflects the potential acids in soils.
Data analysis
The obtained data was subjected to Kolmogorov-Smirnov (K-S) test analysis, followed by variance analysis. Utilizing the software SPSS 19.0 (IBM, Armonk, NY, USA), an ANOVA test was conducted to delve into potential differences in tree attributes (height, diameter at breast height (DBH), individual volume, disease index, and undercrown height). Additionally, the study examined plant diversity metrics within the forest stand, represented by the Shannon-Wiener index, as well as soil nutrient content, specifically focusing on carbon (C), potassium (K), nitrogen (N), and phosphorus (P). Furthermore, rainfall-runoff characteristics, such as pH, total nitrogen (TN), nitrate-N, ammonium-N, and dissolved organic carbon (DOC), were also analyzed across varying slope positions. Statistical significance was established at a threshold of p < 0.05. To visually depict the findings, the Origin 2018 software (OriginLab, Northampton, MA, USA) was utilized. Additionally, SPSS 19.0 was employed to scrutinize the correlations among soil parameters. The intricate relationship between forest stand characteristics and soil parameters was analyzed and visually presented using the Canoco 5.0 software. According to the RDA based on the forestland and forest characteristics and the soil properties at different slope positions, we identified which soil physical and chemical factors affect the forestland and forest.
The intricate relationship between forest stand characteristics and soil parameters was analyzed and visually presented using the Canoco 5.0 software, allowing us to identify which soil physical and chemical factors affect the forestland and forest based on the RDA analysis conducted on the forestland and forest characteristics and soil properties at different slope positions. Furthermore, a Pearson correlation analysis was performed on rainfall and runoff parameters within the forest ecosystem during the years 2018, 2019, and 2020, leveraging the SPSS 19.0 software.
Results
Pinus massoniana forest characteristics and understory vegetation states
There were significant differences in tree height, diameter at breast height (DBH), and individual volume of trees among different slope positions in the Pinus massoniana forest land (Figs. 1A–C and 4). The tree height was significantly greater in the lower slopes than in the upper and middle slopes (p = 0.04) (Fig. 4D). The mean DBH was 19.30 ± 3.22 cm, 20.16 ± 1.87 cm and 21.46 ± 5.25 cm in the upper (US) middle (MS), and lower slopes (LS), respectively (Fig. 4E). This indicated that tree height, DBH and individual volume decreased with slope elevation, but the individual tree volume decreased much more than did tree height and DBH (Fig. 4F). By comparing the DBH value of Pinus massoniana trees among different positions, it was found that the DBH value was significantly (p = 0.03) lower in the LS position (0.87 ± 0.29 m) than in the MS (1.90 ± 0.17 m) and US positions (2.84 ± 0.38 m).

Vegetation and forest characteristics of Pinus massoniana at different sites. ((A) displays the characteristics of Pinus massoniana needles located on the lower slope; (B) presents the characteristics of Pinus massoniana needles on the mid-slope; (C) illustrates the features of Pinus massoniana needles on the upper slope, with corresponding circular images on the right side providing magnified views of the pine needles from different slope positions; (D) depicts tree height; (E) shows the diameter at breast height; (F) represents the individual tree volume of Pinus massoniana; (G) illustrates the disease index; (H) shows the height to the lowest branches of the trees; (I) displays the biodiversity index of the forest floor vegetation).
A total of 20 species belonging to 19 genera and 16 families were found in this survey. Gramineae had the most species (four), followed by fagaceae (two). The Shannon-Wiener index of forest vegetation differed widely among the different slope positions (Figs. 1B and 4A–C and I). The Shannon-Wiener index in the US position was 0.1875 ± 0.02, and this slope position had the fewest species (10% of the species in the investigated woodland). A total of 3 species belonging to 3 families and 3 genera were found in the US, which is significantly lower than the MS and LS positions. The Shannon-Wiener index in the MS position was 1.3073 ± 0.10, and this slope position had 25% of the species in the investigated woodland. The dominant species at the MS position was Parathelypteris glanduligera (Kze.) Ching (58.33%). The dominant species were Arthraxon hispidus (19.67%) and Dicranopteris linearis (19.13%). The Shannon-Wiener index at the LS position was 2.3646 ± 0.14 (Fig. 4). The mean under crown height of Pinus massoniana trees situated at the upper slope of the forest stands at 2.84 ± 0.38 m, exceeding the average height of 0.87 ± 0.29 m found at the lower slope by a substantial 69.48%. The middle slope position boasts an average under crown height of 1.90 ± 0.17 m, which is 50% lower than the upper slope but still 54.22% higher than the lower slope. Upon examining the prevalence of diseases within the Pinus massoniana forest, notable disparities (p < 0.05) were observed in the average disease index across different slope positions, exhibiting a distinct downward trend from the upper to the lower slope. Specifically, the lower slope exhibited the least severe disease occurrence, with a disease index of 11, whereas the upper slope had the highest, with a disease index of 27. The middle slope fell between these two extremes, with a disease index of 16. The under crown height and disease index show the same positional difference, with the US being greater than the MS, and the MS being greater than the LS(p<0.05) (Fig. 4G, H).
Soil distribution characteristics at different slope positions
Soil pH and TN content
The soil pH value of forestland ranged from 5.07 to 7.62. The soil pH value at the LS position (6.41 ± 0.18, n = 15) was higher than the soil pH values at the MS position (5.96 ± 0.13, n = 13, p < 0.05) and the US position (5.96 ± 0.13, n = 13, p < 0.05) (Fig. 7). There was no significant difference in the soil pH values between MS and US (p > 0.05). The TN content in soil was higher in the US position (1.06 ± 0.29 g/kg) than in the LS position (0.85 ± 0.14 g/kg, p < 0.05) and the MS position (0.98 ± 0.07 g/kg, p > 0.05). Pearson’s correlation analysis showed that the TN content in soil was significantly and positively correlated with SOM (r = 0.8568, p < 0.0001), TP (r = 0.3334, p < 0.0001), alkaline-N (r = 0.4979, p = 0.0001), P2O5 (r = 0.3501, p = 0.0095), and Olsen-K (r = 0.6169, p < 0.0001), and highly significantly negatively correlated with soil pH (r = − 0.5459, p < 0.0001) (Fig. 7).
Stoichiometric characteristics of soil C, K, N and P at different slope positions
Our study demonstrates that no statistically significant disparities exist in the concentrations of SOM, SAN, TP, SAP, TK, and SAK among various slope positions (US, MS, and LS) (Fig. 5). Although the SOM content in the LS slope position is marginally higher (17.31 ± 5.57 g/kg) than in the US (17.10 ± 5.51 g/kg) and MS (17.10 ± 5.51 g/kg) slope positions, this difference does not attain statistical significance. Concerning TP content, its variation trend mirrors that of SOM, with a negligible difference between the LS and MS slope positions (0.01 g/kg, p > 0.05) compared to the MS and US slope positions (0.05 g/kg, p > 0.05), both of which are insignificant. Notably, the TK content peaks in the MS slope position (19.72 ± 6.51 g/kg, n = 13), followed by the US (16.65 ± 6.54 g/kg, n = 23) and LS (15.21 ± 7.01 g/kg, n = 15) slope positions, yet these variations in TK content across slope positions also fail to reach statistical significance (p > 0.05). Similarly, the variations observed in SAN and SAP contents follow a pattern akin to TK, with the highest concentrations recorded in the MS slope position, and the differences between slope positions remain non-significant (p > 0.05). Additionally, the SAK content does not exhibit any statistically significant differences among the three slope positions (p > 0.05).

Variations in SOM, TP, TK, alkaline-N, PZOS, and Olsen-K among different slope positions.
The results showed that on RDA, the values of the first and second ordinate axes were 0.6655 and 0.3345, respectively, indicating that they explained 66.55% and 33.45% of the variations in forestland and trees, and the accumulative interpretation was 100%. Therefore, the first two axes can better reflect the correlation between forest and forestland characteristics and soil physical and chemical factors. The pH value was the main environmental factor affecting DBH, tree height, individual volume, and the Shannon-Wiener index (Fig. 6).

RDA plot indicating relationships between forest environment and soil characteristics. (In the diagram, the four-pointed stars represent sample groups under different environmental conditions. Hollow arrows denote environmental factors, while solid arrows signify forest tree and woodland characteristics. The angles between forest trees and woodlands, and environmental factors represent the positive or negative correlation between species and environmental factors (acute angle: positive correlation; obtuse angle: negative correlation; right angle: no correlation). The horizontal axis is the first axis (RDA1), and the vertical axis is the second axis (RDA2), which respectively represent the directions of the largest and second-largest variations in the data, closely related to the most primary and secondary environmental factors) (Fig. 7).

Pearson’s correlation analyses of soil physicochemical characteristics.
Rainfall
Rainfall pH characteristics
From 2018 to 2020, the average rainfall pH was 6.17 ± 1.04 (n = 321), and which showed obvious spatial and temporal differences. The monitoring results demonstrated that the pH value of the samples flowing out of the Pinus massoniana forests was significantly higher than that of the rainfall entering the forest land (p < 0.0001) (Fig. 8). Over the observed period, the increase in the pH value from IF to OW showed that forestland played a positive role in easing acid rainfall-induced damage to the forest. Pearson’s correlation analysis showed that rainfall pH had non-significant negative relationships with other factors (p > 0.05).

Runoff nutrient characteristics from April 2018 to September 2020 (SC Stem interception, IF Into the forest, OW Outflow of woodland).
Concentrations of TN, total soluble N, nitrate-N, and ammonia-N in runoff water
The properties of runoff water varied greatly among different forest sites at annual scales. In 2018, 2019, and 2020, the mean annual TN concentration in runoff water was 3.75 ± 2.82 mg/kg (n = 77), 9.76 ± 5.24 mg/kg (n = 27) and 5.24 ± 2.38 mg/kg (n = 174), respectively (Fig. 8). After the interception and distribution of precipitation in forestland, the TN concentration in the runoff water samples decreased (p < 0.0001) significantly from IF to OW (Fig. 9), and the contents of both soluble total N and ammonia-N in runoff water showed similar changes. However, the differences in nitrate-N and nitrite-N concentrations among different sites in forest land were not as large.
Concentrations of TP, soluble total phosphorus, and phosphate in runoff water
The total phosphorus and phosphate concentrations in runoff water did not vary significantly among different locations. However, the total soluble phosphorus content was significantly higher in runoff water samples than in forest water samples (Fig. 8).
DOC fluxes in soil leachates
The forestland showed a significant DOC-interception effect. This was confirmed by monitoring of forest water samples to determine DOC from 2018 to 2020 (Fig. 8).

Pearson’s correlation coefficients (R2) of pairwise comparisons between variables (rainfall pH, total nitrogen (TN), nitrate-N, ammonia-N, dissolved organic carbon (DOC)) in the forest catchment in 2018 (n = 77), 2019 (n = 70), and 2020 (n = 174).
Discussion
Contact between acid rain and needles is not the direct cause of acid damage to Pinus massoniana trees
Human activities have significantly exceeded the natural evolution rate of the ecosystem, and acid rain poses a great threat to the global environment30. Among all the components of the ecosystem, plants are the primary receptors of acid rain pollution31. Most studies suggest that acid rain directly damages plants upon contact by causing membrane lipid peroxidation and disrupting photosynthesis in the leaves32,33. However, our results show that direct contact between acid rain and pine needles is not the primary cause of Pinus massoniana tree death. In our study, we found significant differences in health indicators such as the height and disease index of Pinus massoniana trees among different slope positions. The difference in Pinus massoniana growth under the same rainfall conditions in a small area indicates that direct contact between acid rain and conifer needles is not the primary factor causing acid damage to these trees.
When precipitation passes through a forest ecosystem, the chemical substances in the water undergo interception and leaching processes and are redistributed34,35. Upon initial contact with the forest canopy, most of the dust and sediment particles on conifer needles, fruits, and branches are carried along the stems or directly penetrate the canopy and enter the soil. This redistribution process significantly changes the concentration of chemical elements in precipitation, and in doing so, affects the growth and distribution of vegetation in the forest ecosystem. Additionally, the litter layer redistributes the precipitation that enters the forest soil by leaching and adsorbing various substances and altering the chemical characteristics of the water that flows out from the forest36,37. Our research confirmed this point. Meanwhile, the results of the vegetation and tree health analyses revealed that, while acid rain can cause tree deaths and affect vegetation growth, the damage to Pinus massoniana trees was not due to direct damage caused by contact between acid rain and the needles. Rather, it was due to the effects of acid rain on the soil microenvironment. That is, substances carried and redistributed into the soil affected the environment in the root zone. This has also been confirmed through our analysis of the health indicators of Pinus massoniana trees, such as height and disease index, at different slope positions. Together, our results indicate that direct contact between acid rain and conifer needles is not the primary cause of acid damage to Pinus massoniana trees under the same precipitation conditions.
The intricate interactions between acid rain and forest ecosystems are far-reaching and complex, significantly impacting the ecological balance and health of forested areas38. The study conducted in the Pinus massoniana forests of the Three Gorges Reservoir area reveals a nuanced understanding of how acid rain affects these ecosystems not through direct contact with foliage, as commonly presumed, but predominantly through changes in the soil microenvironment caused by the hierarchical vertical redistribution of acidic substances. This redistribution is largely influenced by forest stratification, which plays a critical role in the spatial and compositional diversity of the forest.
Stems alleviate the direct entry of acid rain into forest soil, but also distribute a large amount of dry deposition material into the forest
The vertical stratification of forests, involving different layers from the canopy to the forest floor, acts as a multi-level filtration system that modifies the chemical composition of acid rain as it passes through each layer39. These results indicate that Pinus massoniana forest woodlands can effectively intercept total nitrogen concentration, soluble total nitrogen, and ammonia nitrogen. The findings suggest that the canopy and litter layers have a profound ability to intercept and adsorb harmful substances before they reach the soil, thereby mitigating potential acidification at the root level. This interception is crucial, as the soil layer’s health directly influences nutrient uptake and overall plant health40. Our studies have corroborated these findings, highlighting that enhanced vertical stratification in forests can lead to more effective management of acidic depositions, thereby promoting healthier forest ecosystems. Forest litter can significantly affect the pH of the solution passing through it, and its effect is stronger than that of the forest canopy. The forest canopy mainly provides dry deposition materials, while the litter layer effectively intercepts these materials41,42. When precipitation enters Pinus massoniana forests, the substances on the canopy and stem (such as nutrients, basic ions, heavy metals, and plant secretions) strongly interact with the rainfall, allowing H + in the rain water to be diluted, and increasing the pH of the runoff water that reaches the ground8. Nevertheless, these findings warrant careful consideration, because the only way to evaluate this property experimentally is to artificially add acid to the ecosystem, which may cause effects that would not occur under normal environmental loads.
Changes in the microenvironment of forest soils fundamentally affect the healthy growth of Pinus massoniana trees
We detected significant differences in soil pH among the different slope positions, with the pH of the soil at upper slope positions being significantly lower than that of soil at lower slope positions. This indicates that acidic substances are intercepted by litter and surface soil after passing through the forest canopy. In this way, a normal root environment suitable for substance absorption formed in the lower slopes of the forest, providing a good microenvironment for the growth of Pinus massoniana and other vegetation. Although the soil pH needs to be very low to cause injury to Pinus massoniana in laboratory-controlled experiments, in natural environments, Pinus massoniana is exposed to a slightly acidic environment for a long time, which will undoubtedly interfere with and hinder nutrient absorption by its roots43,44. Therefore, it is possible that the root system of Pinus massoniana is more sensitive to acidic soil conditions. This understanding of the close connection between the health of Pinus massoniana, its root system, and the soil pH provides opportunities to devise strategies to reduce the impacts of acid rain on these forests.
Conclusion
The comprehensive research on the Pinus massoniana forests in the Three Gorges Reservoir area elucidates the significant role of vertical stratification in forest health and material balance, highlighting how this natural structure aids in the absorption, fixation, and exchange of substances entering the forest. Through extensive monitoring, clear spatial disparities in the morphological characteristics and health status of these forests were observed. Lower slope areas, benefitting from the effective interception of acidic substances by litter, exhibit denser canopies, vibrant needles, and higher biodiversity compared to the degraded conditions on upper slopes. This indicates that the primary impact of acid rain on forest decline is mediated not through direct contact with foliage but via alterations in the soil microenvironment that affect the root health and growth of understory vegetation and Pinus massoniana trees. Such insights emphasize the necessity of shifting forest management practices towards enhancing vertical stratification and the forest’s natural buffering capacities to mitigate acid rain effects. This involves strategic planting, conservation of undergrowth, controlled thinning to optimize environmental conditions, and the development of site-specific strategies that consider topographical variations in acid deposition susceptibility. Implementing these ecologically informed practices is crucial for safeguarding forest ecosystems against the evolving global environmental challenges, thereby ensuring their health and sustainability. Future research should persist in exploring these dynamics to develop innovative management strategies that further mitigate the adverse impacts of environmental stressors on forests.
Data availability
The data used to support the findings of this study are available from the corresponding author upon request.
Abbreviations
- BA:
-
Basal area
- CFA:
-
Flow injection chemical analyzer
- DBH:
-
Diameter at breast height
- DOC:
-
dissolved organic carbon
- IF:
-
Into the forest
- LS:
-
Lower slope
- MS:
-
Middle slope
- NW:
-
Northwest
- OW:
-
Outflow of woodland
- RDA:
-
Redundancy analysis
- SAK:
-
Soil available potassium
- SAN:
-
Soil alkaline nitrogen
- SAP:
-
Soil available phosphorus
- SC:
-
Stem interception
- SOM:
-
Soil organic matter
- SW:
-
Shannon-Wiener index
- TK:
-
Total potassium
- TN:
-
Total nitrogen
- TP:
-
Total phosphorus
- US:
-
Upper slope
References
Barbeito, I., Cañellas, I. & Montes, F. Evaluating the behaviour of vertical structure indices in scots pine forests. Ann. Sci. 66, 710 (2009).
Baillie, B. R. & Neary, D. G. Water quality in New Zealand’s planted forests: a review. N Z. j. Sci. 45, 7 (2015).
Sun, X. et al. Effect of canopy openness and meteorological factors on spatial variability of throughfall isotopic composition in a Japanese cypress plantation. Hydrol. Process. 32, 1038–1049 (2018).
Francis, J. R., Wuddivira, M. N. & Farrick, K. K. Exotic tropical pine forest impacts on rainfall interception: Canopy understory and litter. J. Hydrol. 609, 127765 (2022).
Cao, Y. et al. Effects of forest plantations on rainfall redistribution and erosion in the red soil region of Southern China. Land. Degrad. Dev. 19, 321–330 (2010).
Shao, Y. et al. Effects of vertical cover structure of grass and shrub on reducing runoff and soil loss under natural rainfall in the loess hilly region. Acta Ecol. Sin. 42, 322–331 (2022).
Wu, J. et al. Prolonged acid rain facilitates soil organic carbon accumulation in a mature forest in Southern China. Sci. Total Environ. 544, 94–102 (2016).
Zhao, Z. et al. Changes in rainfall pH after passing through the forest canopy: increase in throughfall but decrease in stemflow. J. Hydrol. 624, 129955 (2023).
Kacalek, D. et al. Former agriculture impacts on properties of Norway spruce forest. For. Syst. 20, 437–443 (2011).
Lorenzon, A. S., Dias, H. C. T. & Tonello, K. C. Escoamento Da Água Da Chuva Pelo Tronco das Árvores em uma floresta estacional semidecidual. Rev. Árvore. 39, 3 (2015).
Kikuchi, K., Sasaki, Y. & Suzuki, T. One-to-one correspondence of acidity between Rainfall and Throughfall in Relation to Canopy openness in forests. J. Atmospheric Electricity. 24, 49–60 (2004).
Gou, J. F. et al. Application of stable isotope Tracer to Study Runoff Generation during different types of rainfall events. Water. 10, 538 (2018).
Artemkina et al. Micromosaic structure of vegetation and variability of the Chemical composition of L layer of the litter in dwarf shrub-green Moss Spruce forests of the Northern Taiga. Contemp. Probl. Ecol. 7, 11–19 (2018).
Humberto, B. C. et al. Grass barrier and vegetative Filter Strip Effectiveness in reducing runoff, sediment, Nitrogen, and Phosphorus loss. Soil Sci. Soc. Am. J. 68, 1670–1678 (2004).
Haijun, L. I. et al. The water quality changes during rainfall in natural Picea schrenkiana var.tianschanica forest ecosystem in the Middle Tianshan Mountains. Acta Ecol. Sin. 38, 641–646 (2010).
Martinez-Rodriguez, G. A. & Vazquez, M. Evaluation of best management practices to reduce nutrient contaminant losses in runoff from broiler litter-amended soils. I. Alum additions. J. Agric. Univ. Puerto Rico. 94, 41–59 (2010).
Desie, E. et al. Litter quality and the law of the most limiting: opportunities for restoring nutrient cycles in acidified forest soils. Sci. Total Environ. 699, 134383 (2020).
Li, D. et al. Adsorption behavior and mechanism of modified Pinus massoniana pollen microcarriers for extremely efficient and rapid adsorption of cationic methylene blue dye. J. Hazard. Mater. Mar. 5, 465. https://doi.org/10.1016/j.jhazmat.2023.133308 (2024).
Yan, M. et al. Endofungal bacteria and ectomycorrhizal fungi synergistically promote the absorption of organic phosphorus in Pinus massoniana. Plant. Cell. Environ. 47, 600–610 (2024).
Su, J. W. et al. Effect of needle damage on the release rate of Pinus massoniana (Pinus massoniana) volatiles. J. Plant. Res. 122, 193–200 (2009).
Ren, L. Y. et al. Decomposition characteristics of leaf litter of Pinus massoniana and Schima superba in the acid rain region of Chongqing, China. Ecol. Environ. Sci. 22, 246–250 (2013).
Li, Z. Y. et al. Long-Term effects of Liming on Health and Growth of a Pinus massoniana stand damaged by Soil Acidification in Chongqing, China. PLOS ONE. 9, e94230 (2014).
Víctor, G. et al. Diagnosis for the development of Pinus massoniana and Pinus brutia established in the arboretum at Universidad Austral De Chile. Valdivia Bosque. 27, 57–63 (2006).
Gu, X. et al. Differential aluminum tolerance and absorption characteristics in Pinus massoniana seedlings colonized with ectomycorrhizal fungi of Lactarius deliciosus and Pisolithus Tinctorius. J. Forestry Res. 34, 1523–1533 (2023).
Gu, X. et al. LacBicoloricolor MobilizeslabileLaluminumuminuinorganicrphosphatesphate in Rhizosphere Soil of Pinus massoniana Seedlings grown GrownyellowYacidicAsoilc Soil. Am. Soc. Microbiol. 86 (8), e03015–e03019. https://doi.org/10.1128/AEM.03015-19 (2020).
Mishra, B. A. Study on the micro-environment, litter accumulation on forest floor and available nutrients in the soils of broadleaved, mixed pine and pine forests at two distinct altitudes in Meghalaya, North East India. Curr. Sci. 99, 1829–1833 (2010).
Homeier, J. et al. Tropical andean forests are highly susceptible to Nutrient inputs-Rapid effects of experimental N and P addition to an Ecuadorian Montane Forest. Plos One. 7 (10), e47128. https://doi.org/10.1371/journal.pone.0047128 (2012).
Du, E. et al. NEECF: a project of nutrient enrichment experiments in China’s forests. J. Plant. Ecol. 6, 428–435 (2013).
Callesen, I. Commentary to the paper by Roell, Nord-Larsen et al. ‘Mapping tree species ecograms based on soil pH and soil water availability across Denmark’. Forestry. 95, 740–741 (2022).
Bachman, M. et al. A comparative Assessment of Runoff Nitrogen from Turf, Forest, Meadow, and mixed landuse watersheds. Jawra J. Am. Water Resour. Association. 52, 397–408 (2016).
Li, Z. G. et al. Physiologic responses and sensitivity of six Garden plants to simulated acid rain. Acta Horticulturae Sinica. 38, 512–518 (2011).
Ishibashi, M. & Terashima, I. Effects of continuous leaf wetness on photosynthesis: adverse aspects of rainfall. Plant. Cell. Environ. 18, 431–438 (2010).
Zhou, M. K. et al. Effects of simulated acid rain on photosynthesis of leaves and organic acid secreted by roots of Cycas debaoensis Y. C. Zhong et C. J. Chen. J. South. Agric. 43, 587–591 (2012).
Kim, J. K. et al. Plot-scale study of surface runoff on well-covered forest floors under different canopy species. Quatern. Int. 344, 75–85 (2014).
Thompson, A., Davis, J. D. & Oliphant, A. J. Surface runoff and soil Erosion under Eucalyptus and Oak Canopy. Earth. Surf. Proc. Land. 41, 1018–1026 (2015).
Singh, G. Enhancing growth and biomass production of plantation and associated vegetation through rainwater harvesting in degraded hills in southern Rajasthan, India. New Forest. 43, 349–364 (2012).
Liu, X. J. et al. Runoff velocity controls soil nitrogen leaching in subtropical restored forest in Southern China. For. Ecol. Manag. 548, 121412 (2023).
Mcdonald, R. I. et al. Forest harvesting and land-use conversion over two decades in Massachusetts. For. Ecol. Manage. 227, 31–41 (2006).
Garbarino, M. et al. The larch wood pasture: structure and dynamics of a cultural landscape. Eur. J. for. Res. 130, 491–502 (2011).
Wanderlei, B. et al. Linking above and belowground carbon sequestration, soil organic matter properties, and soil health in Brazilian Atlantic Forest restoration. J. Environ. Manage. 344, 118573 (2023).
Bai, Y. et al. Effects of a broadleaf-oriented transformation of coniferous plantations on the hydrological characteristics of litter layers in subtropical China. Global Ecol. Conserv. 25, e01400. https://doi.org/10.1016/j.gecco.2020.e01400 (2021).
Yang, A. et al. Chronic enhanced nitrogen deposition and elevated precipitation jointly benefit soil microbial community in a temperate forest. Soil Biol. Biochem. 193, 109397 (2024).
Johnson, D. W., Turner, J. & Kelly, J. M. The effects of acid rain on forest nutrient status. Water Resour. Res. 18, 449–461 (1982).
Mulder, J., Pijpers, M. & Christophersen, N. Water Flow paths and the spatial distribution of soils and exchangeable cations in an acid rain-impacted and a Pristine Catchment in Norway. Water Resour. Res. 27, 2919–2928 (1991).
Acknowledgements
We thank the associate editor and the reviewers for their useful feedback that improved this paper. We thank Jennifer Smith, PhD, from Liwen Bianji (Edanz) (www.liwenbianji.cn), for editing the English text of a draft of this manuscript.
Funding
This study was supported by Sichuan Science and Technology Program (2024NSFSC1234) and the Youth Innovation Promotion Association of the Chinese Academy of Sciences (NO.2021375).
Author information
Authors and Affiliations
Contributions
Yan Y. Y. conducted thorough consideration and planning for the overall structure and content of the entire paper. Huang Z. H. provided valuable suggestions for the writing and revision of certain key sections in the paper, especially regarding data analysis and interpretation of results. Yan K. primarily contributed his professional insights and innovative thinking, engaging in in-depth discussions and supplements for certain viewpoints in the paper. Liu Q. also reviewed and revised the paper with great care and attention, ensuring its standardization and accuracy in terms of format, language, and other aspects.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This article does not contain any studies with human participants or animals performed by any of the authors. Regarding the species chosen for our study, we have indeed selected the commonly found Pinus massoniana, or the Pinus massoniana. Given its widespread distribution in various ecological environments and its stable population, we firmly believe that our research activities pose no substantial harm to this species. We have always adhered to the principles of respecting and protecting the natural environment and biodiversity, ensuring that our sample collection and research processes comply with ethical principles and standard operating procedures of scientific research. It is important to note that our research on this species does not require any institutional permits. We clarify this point to ensure that our study remains compliant with all relevant regulations. We are fully committed to conducting our research in a responsible and ethical manner, and we take utmost care to avoid any potential harm to the Pinus massoniana species.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Yan, Y., Huang, Z., Yan, K. et al. Effects of vertical forest stratification on precipitation material redistribution and ecosystem health of Pinus massoniana in the Three Gorges Reservoir area of China. Sci Rep 14, 27596 (2024). https://doi.org/10.1038/s41598-024-79097-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-79097-5
