
Abstract
Fibrosis is involved in 45% of deaths in the United States, and no treatment exists to reverse progression of the disease. To find novel targets for fibrosis therapeutics, we developed a model for the differentiation of monocytes to myofibroblasts that allowed us to screen for proteins involved in myofibroblast differentiation. Inhibition of a novel protein target generated by our model, talin2, reduces myofibroblast-specific morphology, α-smooth muscle actin content, and collagen I content and lowers the pro-fibrotic secretome of myofibroblasts. We find that knockdown of talin2 de-differentiates myofibroblasts and reverses bleomycin-induced lung fibrosis in mice, and further that Tln2−/− mice are resistant to bleomycin-induced lung fibrosis and resistant to unilateral ureteral obstruction-induced kidney fibrosis. Talin2 inhibition is thus a potential treatment for reversing lung and kidney fibroses.
Similar content being viewed by others

Introduction
Fibrosing diseases—including pulmonary fibrosis, congestive heart failure, liver cirrhosis, and end-stage kidney disease—are involved in 45% of deaths in the United States1,2. There are few FDA approved treatments for fibrosis2,3. Currently approved FDA treatments (pirfenidone and nintedanib) slow, but do not reverse, the progression of fibrosis4. Further, the mechanisms of action of pirfenidone and nintedanib are poorly understood5. To date, only one treatment (recombinant pentraxin-2, PRM-151) has shown even a modest ability to reverse fibrosis in some patients6.
The ultimate goal of any fibrosis treatment is to reverse disease1. Interrupting collagen deposition destabilizes scar tissue and is a necessary prerequisite for reversing fibrosis1. Another prerequisite for reversing fibrosis is removing deposited extracellular matrix (ECM) while regenerating tissue, and monocyte-derived cells are capable of both ECM-removal and tissue regeneration7,8.
Myofibroblasts are key to fibrosis progression, but the term “myofibroblast” denotes the function of a cell rather than a precise cell lineage9. Myofibroblasts can arise from several progenitors, including hepatocytes, fibroblasts, epithelial cells, and monocytes1,9,10. Myofibroblasts promote scar tissue formation and maintenance by actively depositing ECM, secreting pro-fibrotic signals, and physically stiffening tissues by tension force generated by their cytoskeleton1,11,12. Myofibroblast contractility is key to the progression of fibrosing diseases, and tissue stiffness can activate myofibroblast phenotypes independent of TGF-β13. Myofibroblasts are formed or recruited in response to injury10. Normal, undamaged tissues “stress-shield” cells from tension, with intact ECM supporting the stress. By reducing the stiffness of the environment, normal healthy tissues reduce myofibroblast activation14. Among myofibroblast precursors, monocytes are unique in that they can be recruited from the bloodstream to different tissues in the body15. These monocyte-derived myofibroblasts may provide up to a third of myofibroblasts in liver16, kidney17, lung18,19, and skin fibroses20.
Myofibroblast activation in damaged tissue can either be resolved, as in the case of scarless wound healing, or can lead to the formation of scar tissue12. As myofibroblasts are key to scar tissue formation and maintenance, reversing fibrosis will involve de-activating or removing myofibroblasts from scar tissue12. It has been suggested that apoptosis21 or de-differentiation10 are the two main pathways by which the body de-activates myofibroblasts22.
Mechanosensing is key to maintenance of the myofibroblast phenotype12. Mechanosensing is a dynamic process integrating multiple signals between cell surface integrins, intracellular machinery, and secreted signals11. In mechanosensing, changes in interactions between integrins, intracellular proteins, and the actin cytoskeleton can lead to signaling changes in what might otherwise appear as a static environment23. Among these changes are the maturation of focal adhesions (FAs), which are a dynamic, responsive link at the cell surface between the actin cytoskeleton and the ECM23,24. Mechanoproteins interact in FAs, leading to a complex signaling and force-distribution environment25. FAs exist in a dynamic state that ranges from transient—in migrating cells—to super-mature in some less motile myofibroblasts26. Fibrillar adhesions (FBs) are a complex of cytoskeletal machinery and actin that connect to FAs and localize into long intracellular bundles27,28 .
Talin is an intracellular tension-sensing adapter protein that is key to FAs and FBs27. Talin has two isoforms (1 and 2) that share ~ 76% homology and are both 270 kDa29. Talin is composed of an N-terminal head ___domain and a C-terminal rod ___domain30,31. Talin1 and talin2 have different expression patterns in different tissues29,32,33. Talin1 and talin2 have distinct but overlapping interactions34,35,36, cellular localizations27,34, and functions34,36. Talin2 is sometimes capable of rescuing phenotypes caused by talin1 knockout, and vice-versa37. Talin2 has splice-variants, though the function of these has not yet been determined29,32. Talin1 is ubiquitously expressed in vertebrates, and the knockout is embryonic lethal38. Talin2 is expressed only in certain tissues, and knockout mice are either asymptomatic39 or mildly dystrophic at advanced ages29.
Talins are key to mechanosensing and mechanotransduction34,36. Talins function as adapter proteins, classically explained as binding integrins with their head domains, and binding vinculin and other FA and FB proteins with their rod domains31,40,41,42,43. Recent studies have shown that talins interact more broadly with the adhesome, as talin’s individual domains each have affinities of their own for integrins or cytoskeletal proteins34,37,44,45. Talins provide a framework on which all components of focal adhesions are built34.
Here we show that monocytes and fibroblasts only differentiate into myofibroblasts when adhered to a surface of sufficient stiffness. We were motivated to target talin2 by an mRNA sequencing (RNAseq) study comparing gene expression of myofibroblasts cultured on pro-fibrotic, sufficiently stiff surfaces versus culture on anti-fibrotic, insufficiently stiff surfaces. Knockdown of talin2 interrupts the ability of monocytes to sense stiffness, prevents monocyte-myofibroblast differentiation, and de-differentiates myofibroblasts. Validating the role of talin2 in fibrosis, we show that a Tln2−/− mouse demonstrates a phenotype of reduced fibrosis. We further show that knockdown of talin2 may have therapeutic utility in a pulmonary fibrosis model in mice.
Results
Myofibroblast differentiation is governed by adhesion and substrate stiffness
We began our studies by investigating the features of cell adhesion that lead to efficient differentiation of monocytes into myofibroblasts, seeking to identify a key pathway that could be inhibited to prevent differentiation. This led to the identification of a key role of talin2, knockdown of which both prevented differentiation of monocytes into myofibroblasts and moreover dedifferentiated existing myofibroblasts. We then investigated talin2 knockdown in murine models of pulmonary fibrosis and kidney fibrosis, which confirmed a key role of talin2.
Our results demonstrate that monocyte differentiation into myofibroblasts is governed by the cell adherent state, including the elastic modulus of the adhesion substrate. Adding pro-fibrotic factors (tryptase, IL-13) for 1 h to human monocytes in suspension, and afterwards culturing these cells adherently, does not result in an increase in myofibroblast differentiation (as determined by morphological demonstration of a clear spindle shape) when compared to monocytes exposed to the same pro-fibrotic factors for the same amount of time while adhered (Fig. 1A). Adhered monocytes also increase their double-positive αSMA and collagen I content (Fig. 1B) when compared to suspended-then-adhered monocytes, as determined by flow cytometry.

Monocyte-to-myofibroblast differentiation requires multiple adhesion-related checkpoints to proceed. (A) Freshly isolated human monocytes were split into two groups, both treated with or without pro-fibrotic factors (tryptase, IL-13): one group was adhered to a tissue-culture treated surface and treated with pro-fibrotic factors (tryptase, IL-13), and then washed, while the second group was treated with pro-fibrotic factors (tryptase, IL-13) for 1 h in suspension, and was then washed and cultured on a tissue-culture treated surface. Both groups then differentiated in fresh medium lacking the pro-fibrotic factors over 3 days into myofibroblasts adherent to the tissue-culture treated surface. Differentiation into myofibroblasts was determined by a highly-elongated, spindle-shaped morphology (see Figure S3 for example). Due to donor variability, readouts are normalized to the same individual donor’s adhered myofibroblast counts, untreated with tryptase or IL-13. (B) Myofibroblasts were then removed from the surface and assessed for the percentage of αSMA- and collagen I-positive cells by flow cytometry, again normalized for each individual donor. (C) Monocytes plated on fibronectin-coated surfaces differentiated into myofibroblasts more on stiffer than on softer surfaces, with or without a pro-fibrotic factor present. (D and E) Monocytes differentiate into myofibroblasts at increasing frequency when cultured at fibronectin-coated 1 kPa, 12 kPa, or functionally infinite kPa surfaces (tissue culture treated plastic), as measured by (D) morphology (normalized to 12 kPa values, statistics vs. 12 kPa) and (E) percentage of cells that are αSMA and collagen I double-positive by flow cytometry (normalized to 12 kPa values, statistics vs. 12 kPa). Due to donor cell variability, readouts are normalized and compared to readout at 12 kPa for each individual donor. (E) Donor PBMC, isolated monocytes, and PBMC depleted of monocytes were assessed by flow cytometry and normalized to the 12 kPa readouts as well. (F) Surfaces of 1, 12, and functionally infinite kPa stiffness induce human fibroblast cultures to become increasingly αSMA- and collagen I-positive by flow cytometry. n ranges from 3 to 20. * = statistical significance of P < 0.05, < 0.01, or < 0.001, Statistical comparisons are to the 12 kPa value for each dataset to account for the variability of responses of primary human monocytes (D–F), 2-way ANOVA for panel C, Sidak post-test, 1-way ANOVA with Dunnett’s post-test for other panels. Each of panels D, E, and F are significant by ANOVA post-test for trend between column-based datasets. Error bars are mean ± SEM.
Second, we observed that human monocytes are unable to differentiate into myofibroblasts under pro-fibrotic conditions on surfaces that are softer than 1 kPa even when adherent; by contrast, monocyte-myofibroblast differentiation can occur and be potentiated at 12 kPa (Fig. 1C) in the presence of a pro-fibrotic factor. For reference, Supplemental Table 1 (data from46,47,48,49) shows the stiffnesses of various human tissues. Culture on surfaces that had been pre-coated with the ECM protein fibronectin supported myofibroblast differentiation even without pre-treatment with a pro-fibrotic factor, yet in a manner that was dependent on the substrate stiffness, with monocytes cultured on 1, 12, and functionally infinite kPa fibronectin-coated surfaces showing increasing amounts of myofibroblast differentiation (Fig. 1D), including increasing percentages of cells double-positive for αSMA and collagen I (Fig. 1E).
While monocytes differentiate into myofibroblasts, these monocyte-derived myofibroblasts are a minority population in scars. Fibroblast-derived myofibroblasts constitute a majority of cells in scar tissue16,17,18,19,20. Cultured fibroblasts also increase the percentage of αSMA and collagen I-double-positive cells on higher surface stiffness fibronectin-coated substrates, both in the absence and presence of pro-fibrotic factors (Fig. 1F).
Talin2 is upregulated in myofibroblasts cultured on stiff surfaces
Using these results (Fig. 1C-E) as a guide, we cultured human monocytes from three donors on 1 and 12 kPa fibronectin-coated surfaces, isolated total mRNA from the population, and performed an RNAseq investigation (Supplemental tables 2 and 3). This RNAseq analysis revealed both individual genes and pathways that were differentially expressed between monocytes cultured at 1 and 12 kPa. Upregulated mRNA sequences included collagens (e.g., collagen XXII upregulated 22-fold) and chemotactic factors (CCL22 upregulated 13-fold). However, analysis of upregulated pathways allowed for a more inclusive and complete picture of the changes induced by culturing monocytes on stiffer surfaces under pro-fibrotic conditions. Specifically, upregulated pathways related to cell adhesion included the paxillin and integrin pathways. Among the individual genes that were lower-expressed at 1 kPa (not supporting myofibroblast differentiation) relative to 12 kPa (supporting), we noticed the stress sensor talin2, which was reduced by 3-fold on the surface that did not allow monocyte-to-myofibroblast differentiation.
Based on this observation, we focused the remainder of this study on talin2, an intracellular tension-sensing modulator of adhesion mechanics. Talin2 transcripts are also upregulated in monocyte-derived cells in idiopathic pulmonary fibrosis patients (IPF, Figure S1, data from the IPF cell atlas50).
We explore the potential of integrins, also involved in stress regulation, to modulate myofibroblast differentiation in a companion study51.
To confirm that the measured mRNA reduction in the sequence for talin2 at low stiffness corresponded to a drop in the protein expression level of talin2, we cultured monocytes and fibroblasts on surfaces of increasing stiffness in the presence and absence of a pro-fibrotic signal. Expanding on the staining for collagen I and aSMA in Fig. 1E and F, talin2 was low at 1 kPa and increased in both human monocytes (Fig. 2A) and human fibroblasts (Fig. 2B) on stiffer surfaces in more pro-fibrotic environments.

Increasing surface stiffnesses and pro-fibrotic conditions increase the intensity of talin2 immunostaining in human cells. (A) Freshly purified primary monocytes, and (B) immortal fibroblasts, were cultured on surfaces of increasing stiffness and in the presence of pro-fibrotic factors. n ranges from 4 to 8. * = statistical significance of P < 0.05, < 0.01, or < 0.001, Statistical comparisons are to the 12 kPa value for each dataset, 1-way ANOVA with Dunnett’s post-test, error bars are mean ± SEM. (A) and (B) are significant by ANOVA post-test for a trend between column-based datasets.
In preparation for future mouse fibrotic model experiments, we wanted to determine if mouse cells also increase talin2 in more pro-fibrotic environments on stiffer surfaces. We cultured monocytes purified from mouse spleens and mouse fibroblasts under conditions similar to those used in our study of human cells reported in Figs. 1 and 2. Mouse monocytes differentiated into morphologically spindle-shaped myofibroblasts with increasing surface stiffness (Fig. 3A), increasing positivity for αSMA and collagen I staining (Fig. 3B), and increasing talin2 staining (Fig. 3C). Mouse fibroblasts also increase in αSMA and collagen I staining (Fig. 3D) and talin2 staining (Fig. 3E) on stiffer surfaces in presence of tryptase as a pro-fibrotic agent.

Increasing surface stiffnesses and pro-fibrotic conditions increases myofibroblast differentiation and the intensity of talin2 immunostaining in mouse cells. Freshly purified mouse monocytes cultured at 1, 12, and functionally infinite kPa form myofibroblasts at increasing amounts, as measure by (A) morphology and (B) percentage of cells that are αSMA and collagen I double-positive. (C) These same populations show increasing talin2 staining intensity. Mouse fibroblasts cultured under identical conditions also show increasing numbers of (D) cells that are αSMA and collagen I double-positive and (E) increased talin2 staining intensity. n ranges from 2 to 8. Statistical comparisons are to the 12 kPa value for each dataset. * = statistical significance of P < 0.05, < 0.01, or < 0.001, 1-way ANOVA with Dunnett’s post-test, error bars are mean ± SEM. Panels A, B, and C were tested for trend by ANOVA post-test for a trend between column-based datasets. Each panel is significant. Error bars are mean ± SEM.
Inhibition of Talin2 reverses myofibroblast differentiation and existing fibrosis
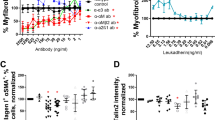
Talin2 is an intracellular protein, and to our knowledge no small-molecules or cell-penetrating peptides exist to inhibit talin2’s protein-protein interactions. To determine if inhibition of talin2 expression could de-differentiate myofibroblasts, we allowed human and mouse monocytes to become myofibroblasts, and treated those myofibroblasts with a mixture of 4 non-targeting silencing RNAs (siRNA), 4 human talin2-targeting siRNAs, and 4 mouse talin2-targeting siRNAs. The mouse and human talin2 siRNA mixtures share one sequence in overlap. To establish a dose range, we treated human monocyte-derived myofibroblasts with the talin2 siRNA mixture, yielding an IC50 of 15 nM (Fig. 4A) for inhibition of myofibroblast morphology. Treatment with 50 nM of talin2 siRNA also reduced the percentage of αSMA and collagen I double-positive human monocytes (Fig. 4B), more than the control siRNA mixture. Human talin2 siRNA significantly reduced the amount of talin2 (Fig. 4C) for human monocyte-derived myofibroblasts.

Silencing RNA against talin2 dedifferentiates human and mouse myofibroblasts. Freshly purified human monocytes allowed to become myofibroblasts were treated with (A) a mixture of 4 non-targeted (control) siRNAs and 4 talin2 siRNAs at the indicated concentrations. (B) Myofibroblasts differentiated from human monocytes were treated with 50 nM mixtures of non-targeted (control) siRNA, and mixtures of siRNA targeting human or mouse talin2, and assessed for (B) the number of αSMA+ collagen I+ double positive cells and (C) the amount of talin2. Myofibroblasts differentiated from mouse monocytes were treated identically, except that individual siRNA was also used in addition to the mixtures, and were assessed for (D) the number of αSMA+ collagen I+ cells and (E) the amount of talin2. n ranges from 4 to 10. * = statistical significance of P < 0.05, < 0.01, or < 0.001. For (A), 2-way ANOVA with Sidak post-test for comparison between PBS, control SiRNA, and Talin-2 siRNA comparison. For (B–E), 1-way ANOVA with Dunnett’s post-test, Student’s t-test for indicated direct comparisons, error bars are mean ± SEM.
Treatment with fluorescently labeled siRNA (siRNA-AF-488) indicated that 50 nM siRNA was capable of entering human monocyte-derived myofibroblasts without the use of transfection reagents (Figure S2). The mixture of talin2 siRNA reduced not only the spindle-shaped morphology within the monocyte-derived myofibroblast population, but also the presence of FAs at the cell periphery, FBs within cells, and the localization of talin2 to the periphery of the cell (Figure S3).
In order to simplify treatment in anticipation of in vivo testing in mice, we treated mouse myofibroblasts with individual siRNAs against mouse talin2, in addition to the same non-targeting siRNA mixture, human talin2 siRNA mixture, and mouse talin2 siRNA mixture as we treated the human monocytes. While the siRNA mixtures trended toward decreasing the number of αSMA and collagen I double-positive cells (Fig. 4D), only talin2 siRNA #2 (sequence: 5’ CUGGAAAAUUCAGUGAUGA 3’ and antisense 5’ UCAUCACUGAAUUUUCCAG 3’) significantly inhibited myofibroblast differentiation by itself. All mixtures and individual siRNAs trended towards decreasing talin2 concentration within the cells, but only talin2 siRNA #2 and #3 were significant (Fig. 4E).
To determine if silencing of talin2 can reduce fibroblast-myofibroblast differentiation, we added siRNAs individually and in mixtures at 50 nM to human and mouse fibroblast cultures. The mixture of human talin2 siRNA reduced both αSMA and collagen I double-positive (Fig. 5A) and talin2-positive (Fig. 5B) myofibroblasts. Mouse siRNA #2 most reduced both αSMA and collagen I double-positive (Fig. 5C) and talin2-positive (Fig. 5D) myofibroblasts in mouse cell cultures.

Silencing RNA against talin2 dedifferentiates human and mouse myofibroblasts. Fibroblasts were cultured on an infinite kPa tissue culture surface and treated with TGF-β, inducing them to become myofibroblasts. These myofibroblasts were treated with 50 nM mixtures of 4 control siRNAs, mixtures of 4 siRNAs targeting human talin2, mixtures of 4 siRNAs targeting mouse talin2, or 4 individual siRNAs targeting mouse talin2. Human myofibroblasts were assessed for (A) the number of αSMA+ collagen I+ cells and (B) the amount of talin2, and mouse myofibroblasts were assessed (C) the number of αSMA+ collagen I+ cells and (D) the amount of talin2. n ranges from 2 to 5. * = statistical significance of P < 0.05, < 0.01, or < 0.001, 1-way ANOVA with Dunnett’s post-test vs. myofibroblast data (A,B) and vs. siRNA control (C,D), error bars are mean ± SEM.
To determine if reductions in myofibroblast spindle-shaped morphology, αSMA and collagen I content, and talin2 content correlated with a reduction in secreted pro-fibrotic factors, we assayed conditioned media from human and mouse monocyte-derived and fibroblast-derived myofibroblasts by ELISA. Treatment with talin2 siRNA did not significantly affect the amount of secreted anti-fibrotic IL-10 from mouse monocyte-derived myofibroblasts (Figure S4A)52 but did reduce the amount of pro-fibrotic cytokines including, IL-23 (Figure S4B)53, CCL22 (Figure S4C)54, IL-6 (Figure S4D)55, CCL17 (Figure S4E)54,56, IL-12 subunit p40 (Figure S4F)57, CXCL1 (Figure S4G)58, TNF-a (Figure S4H)59, and IL-1β (Figure S4I)60. While TGF-b was below the detection limit for this assay, IL-6 is sometimes sufficient to induce myofibroblast differentiation61. While the secretome for human cells was more frequently below the detection limit for this assay, similar results were seen, with reductions in pro-fibrotic TNF-a, MCP-162, and IL-6 (Figure S5). This indicates that knockdown of talin2 in not simply changing myofibroblast morphology, or reducing aSMA and collagen I expression, but also dramatically altering the overall secretome of the treated myofibroblasts.
To determine if treatment with talin2 siRNA could rescue the damage from lung fibrosis in a treatment (i.e., not prophylactic) model, we insulted the lungs of male and female mice with bleomycin. We treated the mice via lung instillation with 50 µL of 200 nM of talin2 siRNA #2 at 7, 9, 11, 14, 16, and 18 days post insult, and euthanized the mice on day 21 post-insult. While no treatment significantly altered mouse weight at day 21, treatment with talin2 siRNA showed a lesser transient weight decrease than did the controls (Fig. 6A). Talin2 siRNA treatment also reduced the amount of collagen in the lungs as measured by a hydroxyproline assay (Fig. 6B and C). Ashcroft scoring63 of the Masson’s trichrome-stained lung sections, performed by a researcher blinded to treatment grouping, confirm that talin2 siRNA treatment improves the histological readout of lung fibrosis (Fig. 6D). The individual talin2 siRNA treatment rescued lung fibrosis compared to non-targeting siRNA and untreated fibrotic lungs (Fig. 6E-L). Thus, talin2 siRNA treatment rescues lung fibrosis in mice in both quantitative (hydroxyproline) and qualitative (Ashcroft scoring) measures, even when treatment was provided 7 days after the inflammatory insult to the lung.


Talin2 siRNA rescues the fibrotic damage from bleomycin insult to mouse lungs. 50 µL of 200 nM talin2 siRNA was administered to mouse lungs 7, 9, 11, 14, 16, and 18 days after insult by bleomycin. (A) Mouse weights after treatment. (B) Collagen content from the right, multi-lobed lung assessed by hydroxyproline assay. (C) Data from B divided by dry weight of right lobes of mouse lungs. (D) Blinded Ashcroft scoring. (E–H) Representative images of left, single lobed lungs stained with Massons’s trichrome. (I–L) Inset of lungs. (E,I) Uninjured lung, (F,J) fibrotic lung, (G,K) fibrotic lung treated with siRNA control, and (H,L) fibrotic lung treated with talin2 siRNA. n ranges from 6 to 8. * = statistical significance of P < 0.05, < 0.01, or < 0.001, significance vs. fibrotic lungs unless otherwise indicated, 2-way ANOVA for panel A, Sidak post-test, 1-way ANOVA with Dunnett’s post-test for other panels, comparisons between siRNA control and talin2 siRNA are Student’s t-test. Error bars are mean ± SEM. Scale bar is constant between (E–H).
To determine if treatment with talin2 siRNA improved lung fibrosis in both male and female mice, we analyzed the hydroxyproline-based collagen data (Figure S6A, B, D, E) and Ashcroft-based qualitative data (Figure S6C and F). The talin2 siRNA treatment improved collagen readouts in male mice and trended toward an improved Ashcroft score (Figure S6A-C). For female mice, only in the Ashcroft score vs. fibrosis was significantly improved (Figure S6D-F), though this is only with n = 3 mice per group.
Comparing the broncheo-alveolar lavage (BAL) of untreated fibrotic and talin2 siRNA-treated mouse lungs, there was reduced talin2 staining intensity in CD45+ cells, and a trend towards reduced numbers of CD45+ talin2+ double-positive cells (Figure S7).
To determine how well our talin2 siRNA treatment of fibrotic lungs compared with a complete reduction of talin2 in a genetically-engineered mouse model, we insulted the lungs of Tln2−/− mice with bleomycin. Tln2−/− lungs were resistant to fibrosis, showing reduced collagen deposition and a significantly improved Ashcroft score (Figure S8). While talin2 siRNA significantly reduced existing fibrosis in the treatment model, talin2 knockout mice were much more resistant to fibrosis overall. Talin2 siRNA treatment reduced the accumulated collagen content of fibrotic lungs by ~ 60% closer to healthy control, while Tln2-/- mice had no increase in collagen content vs. the healthy control (Fig. 6A, Figure S8A). Similarly, talin2 siRNA treatment reduced the Ashcroft score of fibrotic lungs by about 33% closer to the healthy control, while Tln2-/- mice had no increase in Ashcroft score (Fig. 6C, Figure S8C). These changes are probably due to the Tln2-/- mice lacking talin2, and thus functioning as almost a prophylactic model of lung fibrosis as opposed to the treatment model of lung fibrosis used for talin2 siRNA.
To determine if talin2 contributes to kidney fibrosis as well as lung fibrosis, we performed a unilateral ureteral obstruction (UUO) of the left kidney in Tln2−/− mice, allowed the kidneys to become fibrotic for 14 days, sacrificed the mice, and resected the kidneys; the Tln2−/− mice are in the C57BL6 background, so we used C57BL6 mice as controls. The UUO-injured left C57BL6 kidney (Fig. 7B) showed significant fibrosis and damage, which was much reduced in the UUO-injured left Tln2−/− kidney (Fig. 7D). The overall amount of collagen I-positive IHC stained tissue (brown) was much reduced in the Tln2−/− kidney (Fig. 7E), and compared favorably to targeted anti-integrin treatments of kidney fibrosis in our companion work51. Tln2−/− mice that have undergone UUO also have decreased blood urea nitrogen (BUN) and increased bilirubin levels compared to UUO-injured C57BL6 mice (Figure S9). UUO-injured Tln2−/− mice also have higher alanine transferase (ALT) and aspartate transferase (AST) levels than UUO-injured C57BL6 mice (Figure S9E and F). ALT and AST are commonly used markers of liver damage, but kidney fibrosis can counterintuitively cause ALT and AST levels to drop64. Several variables showed no significant change, including albumin, amylase, calcium, creatinine, creatine kinase, and uric acid.

Tln2−/− mice are resistant to UUO-induced kidney fibrosis. Mouse left kidneys were injured by UUO ligation. Kidneys were sectioned and IHC stained using α-collagen-I as a fibrosis marker. (A) Right, healthy C57BL6 kidney. (B) Left, fibrotic C57BL6 kidney. (C) Right, healthy Tln2−/− kidney. (D) Left, fibrotic Tln2−/− kidney. (E) The ratio of fibrotic to healthy tissue in each kidney section. n = 6. * = statistical significance of P < 0.05, < 0.01, or < 0.001, Student’s t-test, error bars are mean ± SEM. Scale bar is constant between images.
Discussion
In this paper, we show that adhesion mechanics plays a key role in myofibroblast differentiation (from monocytes and fibroblasts) and myofibroblast stability, and we show the utility of modulating adhesion machinery to propose a novel treatment for reversing fibrosis. We have shown that culture on somewhat stiffer (12 kPa) surfaces induces the expression of talin2, a key mechanosensing and mechanotransduction component of the cytoskeleton, compared to that on 1 kPa surfaces, which do not support monocyte-myofibroblast differentiation. Inhibition of talin2 can even de-differentiate myofibroblasts, changing their morphology, lowering the amount of αSMA and collagen I produced by myofibroblasts, and changing their secretion profile from pro- to anti-fibrotic. Inhibition of talin2 reverses established lung fibrosis in a murine model, and Tln2−/− mice are resistant to bleomycin-induced lung fibrosis and UUO-induced kidney fibrosis.
Stepwise models of fibroblast-myofibroblast differentiation have previously been proposed that involve the presence of fibrosis-specific ECM and high extracellular stress65. Some studies have shown the importance of stiffness in the development of actin networks66, finding that αSMA stress fibers begin to form at 3–6 kPa67,68, and form fully around 20 kPa69,70, consistent with our findings.
Fibroblast-myofibroblast differentiation on surfaces of different stiffness has also been investigated as it relates to morphology36, and fibroblasts have been reported to form myofibroblasts beginning at 25 kPa22. Hepatocytes have also been shown to become myofibroblasts on a surface of 15 kPa71, similar to our findings for monocytes and fibroblasts. Adhesion has also been recognized as important for monocyte differentiation72, though not in the context of fibrosis.
Cells activate their mechanosensing machinery to assess the composition and stiffness of their environment, which allows them to respond to binding to a surface or a change in rigidity caused by tissue damage66. Classically, this activation begins when talin1 or talin2 activate integrins upon a change in their mechanical environment31, the integrins then bind to the surface ECM. However, the order in which proteins assemble to form FAs has been called into question73. Monocytes could assess surface stiffness with talin1 or talin2 first, or assess the ECM composition of the surface using integrins. Previous studies have found pro-fibrotic74,75 and anti-fibrotic responses (Fig. 6 in76) to various ECM coatings.
Talin1 and talin2 have overlapping but distinct (1) affinities for integrins and intracellular cytoskeletal proteins77, (2) localizations within the cell27, and (3) responsibilities in mechanosensing and mechanotransduction78. Because each talin isoform is mechanosensitive and part of a dynamic mechanosensing and mechanotransduction process, it can be difficult to universally parse what the responsibilities are for talins 1 and 223. The adhesome has been calculated to include almost 700 possible interactions79. Talin2 appears to be responsible for both the initial recruitment of FA and FB proteins at low elastic moduli, and also for mechanosensing and mechanotransduction through FBs, though not for mechanosensing in mature FAs36. However, knockout of talin2 has been shown to suppress large FA formation, while talin1 knockout allowed large FAs to remain78. Further complicating matters, talins also open multiple cryptic protein-binding sites based on their response not just to surface stiffness, but also to tension, and fast and slow applications of force23,37.
Hinz proposes that super-mature FAs are a phenotypic checkpoint critical to a mechanical feedback loop of “extracellular stress and intracellular tension” that regulates the myofibroblast state12. Super-mature FAs form only under tension stress69 and can be eliminated by re-culturing cells on softer surfaces69,70 or by reducing the FA’s maximum available size12. That super-mature FAs might be key control points matches well with our observation that adhesion mechanics modulates monocyte-myofibroblast differentiation. Perhaps the reason that monocytes cannot be induced to become myofibroblasts before being adhered or when adhered to soft surfaces is because they cannot form super-mature FAs. Super-mature FAs may also be involved in a mechanism for scarless healing, as myofibroblasts that become stress-shielded by a fully repaired ECM may then lose the tension necessary to maintain super-mature FAs.
However, even with the diversity of talin1 and talin2 interactions with the cytoskeleton and adhesome30, our immunofluorescence images also match phenotypes observed in other talin2 knockdown and knockout experiments. Hinz’s “super-mature focal adhesions” seen in fibroblasts look remarkably similar in size (30 μm) and morphology to those seen in the monocyte-derived myofibroblasts shown in Figures S3A and S3B26. Similarly, Figure S3C appears phenotypically identical to monocytes cultured on a stiff surface but without pro-fibrotic signals to induce myofibroblast differentiation77. Interestingly, this similarity includes the subcellular localization of talin2 in macrophages77. The edges of talin2 siRNA-treated monocyte-myofibroblasts (Figure S3C) appear to have similarly diffuse actin staining, lacking FAs and FBs, similar to fibroblasts treated with talin2 siRNA80 and in fibroblasts constitutively expressing a talin2 mutant that abrogated its interaction with the cytoskeleton36. This is not merely a difference in a macrophage vs. myofibroblast phenotype, however, as cells from the monocyte-myofibroblast population that failed to form elongated, spindle shaped cells did display increased actin development and adhesion architecture (Figure S3D) compared to monocyte-myofibroblasts dedifferentiated with talin2 siRNA (Figure S3C).
Talin2 is key to both the formation of FAs81 and FBs27,36 and is key to mechanotransduction and the generation of tension36. Talin1 and talin2 respond to cell stress and tension by unfolding in different ways, which exposes cryptic binding sites for integrins, actin, paxillin, and other members of the cytoskeleton and adhesome34,37. Additionally, each subdomain of talin is itself mechanoresponsive and capable of binding to components of the members of the cytoskeleton and adhesome34. Reduction in the overall amount of talin2 might well be responsible for the reduction in super-mature FAs, which is turn could lead to the de-activation of myofibroblasts through apoptosis or dedifferentiation12.
Whether myofibroblasts remain active or de-activate determines whether damaged tissue repairs or scars, as repaired ECM is capable of stress-shielding resident cells and reducing the stress and tension on myofibroblasts14,82,83,84, which could prevent further scarring. Our goal was to induce myofibroblasts to de-activate (either through differentiation or apoptosis) by modulating adhesion machinery to convince myofibroblasts that they are no longer bound to a surface of sufficient stiffness. Several mechanisms have been proposed to de-activate myofibroblasts. Dedifferentiation has been achieved by re-culturing myofibroblasts on softer surfaces69,70,85. Interestingly, a recent mechanobiology investigation resulted in an experimental candidate drug that resolves fibrosis by means of apoptosis21, based on changes in the metabolome of fibroblasts cultured on stiff vs. soft surfaces.
Monocyte-myofibroblast differentiation was far more inhibited on low elastic modulus surfaces than was fibroblast-myofibroblast differentiation (Figs. 1E-F and 2). This could be explained by a phenomenon called “mechanical-memory”, where fibroblasts that have been cultured long-term on functionally infinite modulus surfaces which are sufficient by themselves to induce myofibroblast differentiation13, where the cells may carry with them a mechanical memory that makes de-differentiation difficult to achieve over short timescales22.
Talin2 may be an excellent target for disrupting a myofibroblast’s mechanosensing capability that determines whether a myofibroblast remains a myofibroblast. Talin2 provides a framework on which all other components of the adhesome and cytoskeleton34, and is involved in mechanosensing36,81 and mechanotransduction36. Knockouts of talin2 have few (or no) phenotypic irregularities29,39, while talin1 knockouts are embryonic lethal38. Talin1 is expressed ubiquitously, while talin2 is expressed in monocyte-derived cells, in muscle tissue, and in the brain29,32,33. Thus, the selection of talin2 rather than talin1 for knockdown may be opportune.
To our knowledge, the only method of targeting talin other than siRNA is the small molecule KCH-1521, which decreases adhesion-dependent angiogenesis. KCH-1521, however, does not distinguish between talin1 and talin286. By contrast, siRNA treatments permit a distinction between talin2 and talin1. Fortunately, Accell siRNA offers promise for delivering talin2 knockdown to the lungs, and it could potentially even be delivered as a dry powder by inhalation87. Our doses are a factor of 10- to 100-fold lower than have been used in a previous pulmonary study using Accell siRNA for other targets88.
One of the limitations of this study is that we do not have data on the other adhesome proteins that form the complete focal adhesion, nor do we have data for the contractility of our myofibroblasts before or after inhibition with talin2. The conformational changes in talin induced by culture at 1 kPa, 12 kPa, and infinite kPa are not explored in this study, nor are the possible changes and interactions in the adhesome proteins that form the focal adhesion.
In this study, we show that the mechanical environment of monocytes and fibroblasts is a key checkpoint for their differentiation into myofibroblasts. We also show that the presence of talin2 is important for the differentiation of myofibroblasts and the maintenance of the myofibroblast phenotype, both in culture and in mouse models of lung and kidney fibrosis. We believe that the proteins involved in mechanosensing are potential drug targets for the treatment of fibrosis.
Experimental procedures
Study design
This study examines two related hypotheses. The first is that myofibroblast differentiation (from monocytes and fibroblasts) is governed by adhesion mechanics, and the second is that modulation of adhesion mechanics machinery can be used to guide novel treatment discovery for fibrosing diseases. Specifically, we tested whether inhibition of the intracellular tension sensor talin2 could be used to treat lung fibrosis in mice. Group size was selected based on experience with the pulmonary fibrosis model, particularly based on a pilot experiment using talin2 knockdown in lung fibrosis. Mice were randomized into treatment groups within a cage to eliminate cage effects from the experiment. Treatment was performed by multiple researchers over the course of this study, to ensure reproducibility. Lungs were also resected by multiple researchers, and blinded scoring was used on the fibrosis histology images.
All mouse experiments were performed under supervision with protocols approved by the University of Chicago IACUC, and this study was performed in accordance with the relevant guidelines and regulations. The results of these experiments are reported in this manuscript using ARRIVE guidelines.
Purification of human monocytes
In order to isolate more peripheral blood mononuclear cells (PBMC) than is possible through a single blood donation, PBMCs were purified from leukocyte reduction filters obtained from the University of Chicago blood donation center, in accordance with human subject protocol at the University of Chicago. All leukocyte reduction filters were de-identified and taken from random blood donors regardless of age, race, or gender. Blood was filtered, and leukocytes purified, the same afternoon as a morning blood donation, to reduce the amount of time that PBMCs could adhere to the filter.
The leukocyte reduction filter was sterilized with 70% ethanol, and the blood flow tubes on either end were clamped to prevent flow. The tube through which filtered blood had exited the filter was cut below the clamp, and a syringe containing 60 mL phosphate buffered saline (PBS) was inserted into the tube. Following this, the tube through which unfiltered blood had entered the filter was unclamped, and PBS was slowly pushed through the filter in the opposite direction of the original blood flow. This direction of flow provided the highest recovery of PBMC, approximately 300 million cells per filter.
The collected blood from the filter was then layered with lymphocyte separation media (LSM), and centrifuged at 1300 xg for 20 min. The PBMC layer was then removed by pipetting.
These PBMCs were further purified by use of a negative selection kit for human monocytes (Stemcell, Cambridge, MA), per the manufacturer’s instructions. The expected yield from each filter was approximately 20 million monocytes. Monocytes were then washed by PBS using five successive 300 xg centrifugation steps, in order to remove EDTA from the resulting population. Monocytes were checked for purity using flow cytometry, and average purity was above 95%. Monocytes were cultured immediately following purification, at 100,000 monocytes/cm2.
Purification of mouse monocytes
All mouse experiments were performed under supervision with protocols approved by the University of Chicago IACUC, and the study was performed in accordance with the relevant guidelines and regulations.All PBS, plasticware, filters, glassware and magnets were pre-chilled to 4 C, and kept cold throughout this procedure, to limit the clumping of cells. Importantly, ACK lysis buffer caused cell death, and so was not used in this procedure.
Spleens were resected from healthy C57BL/6 mice, pooled, and were placed in PBS with 1 mM EDTA, 2% fetal calf serum (FCS) to prevent subsequent clumping of cells. Spleens were pushed through a 100 μm filter to disassociate the cells. Monocytes were purified from disassociated cells by use of a negative selection kit (Stemcell), following the manufacturer’s instructions. The purified monocytes were then washed by PBS using five successive 300 xG centrifugation steps, in order to remove EDTA from the resulting population. Monocytes were checked for purity using flow cytometry, and average purity was above 95%. The average yield was 1.5 million monocytes per spleen.
Monocytes were cultured immediately following purification, at 250,000 monocytes/cm2.
Culture of human and mouse monocytes
Human and mouse monocytes were cultured as previously described, using serum-free media (SFM)89. Briefly, SFM for human cells is composed of fibrolife (Lifeline, Frederick, MD), with 1x ITS-3 (Sigma, St. Louis, MO), 1x HEPES buffer (Sigma), 1x non-essential amino acids (Sigma), 1x sodium pyruvate (Sigma), and penicillin-streptomycin with glutamate (Sigma). For mice, 2x concentrations of ITS-3, HEPES buffer, non-essential amino acids, and sodium pyruvate are added. For mouse monocytes, 50 mM beta-mercaptoenthanol (ThermoFisher) was also added, as were pro-fibrotic supplements M-CSF (25 ng/mL, Peprotech, Rocky Hill, NJ) and IL-13 (50 ng/mL). Additionally, M-CSF and IL-13 were refreshed in the media of mouse monocytes after 3 days of culture.
Addition of tryptase (Fitzgerald, Acton, MA) to media was as previously described90. Monocytes were allowed to differentiate for 5 days, and counted based on morphology as previously described91. Examples of myofibroblast morphology are in Figure S3.
To account for donor variability, human cell experiments were normalized to individual donor control groups for each experiment.
Culture of human and mouse fibroblasts
Human fibroblasts (MRC-5, ATCC, Manassas, VA) and mouse fibroblasts (NIH-3T3, ATCC) were cultured in SFM composed for human cells, with 1x concentrations of additives. TGF-b (Peprotech) was added to induce myofibroblast formation at 5 ng/mL92. Cells were cultured at 10,000/cm2, and TGF-β was refreshed in cultures weekly to maintain the myofibroblast phenotype.
Methods for rolling monocytes in the presence of pro-fibrotic factors
100,000 human monocytes were suspended in SFM, and placed onto tissue culture treated plastic, or into low adhesion microcentrifuge tubes (protein lo-bind, ThermoFisher). Tryptase was added at 12.5 ng/mL90, while IL-13 was added at 50 ng/mL. Monocytes were incubated for 1 h, either adhered to the tissue-culture treated plastic or rotating in low adhesion microcentrifuge tubes. After 1 h, the adhered monocytes and rotating monocytes were gently washed with four successive PBS washes to remove both tryptase and IL-13 from the surface of the monocytes. The washed monocytes (including those from low adhesion microcentrifuge tubes) were then cultured on tissue culture treated plastic for 5 days, and the number of myofibroblasts was observed through counting of spindle shaped morphological cells and through analysis of alpha-smooth muscle actin positive (αSMA+) and collagen I + cells. αSMA and collagen I are widely used markers of myofibroblast differentiation93. Antibodies used were anti-collagen I (Biolegend, San Diego, CA), anti-alpha smooth muscle actin (αSMA) (R and D systems, Minneapolis, MN).
Preparation of low surface stiffness plates
Low stiffness tissue-culture plates (1 and 12 kPa) were ordered from Matrigen (San Diego, CA). Culture of myofibroblasts on low stiffness surfaces was similar to the culture conditions on tissue-culture treated plasticware. However, low stiffness surfaces were coated in 10 mg/mL fibronectin (Millipore Sigma) for 1 h at 37 C. Unbound fibronectin was gently removed with 3 successive PBS changes. For experiments involving the comparison of infinite binding surfaces to softer surfaces, the tissue culture treated plasticware was also fibronectin-coated. Data for Fig. 1C was generated using a 96 well plate with a full series of elastic moduli (Matrigen).
mRNA purification and RNAseq
mRNA was purified using the Trizol method94. RNA sequencing was performed by the University of Chicago Center for Research Informatics. Briefly, there were total six samples from three donors, each with paired myofibroblast and control samples. RNA sequencing was run on one flow cell (HFW2JBBXX) using Illumina HiSeq 4000 sequencer. An average of 66.7 million single-end 50 bp sequencing reads were generated for each sample.
The following steps and programs were used for data processing and analyzing. All programs were run using default settings except for otherwise specified. The raw read quality was assessed by FastQC v0.11.2, and reads were aligned to the Human reference genome GRCh38.
(Gencode v24) using STAR v2.5.2a95. The distribution of reads aligned to genome regions were collected by Picard v2.2.4. Gene expression levels were quantified by featureCount96. Expression data were transformed, normalized and filtered; and differentially expressed genes were detected by97. Track hubs were made to visualize reads aligned to genome in UCSC genome browser. Pathways and GO terms enrichment were explored by Ingenuity Pathway Analyses and DAVID tools v6.8 Beta98.
The sequencing quality of raw reads was assessed by FastQC to explore quality distribution per base and other sequence characteristics. It output the overview of sequencing quality for each fasta file to identify possible technical problems. Raw reads were aligned to the Human reference genome GRCh38 (Gencode v24) using the RNA-Seq aligner STAR. Various quality metrics were collected using Picard.
Read counts mapped to 58,656 genes were summarized by featureCount, and then processed by edgeR. Counts for each gene were normalized to the effective library size and transformed to CPM (count-per-million). Low expressed genes were filtered out using cutoff CPM > 0:2 (about 10 reads) in at least 2 samples.
Likelihood ratio test (LRT) in edgeR was used to detect differentially expressed genes (DEGs) between groups. Donor was used as a factor for the statistical test. Benjamini-Hochberg procedure was used to correct for multiple hypothesis testing error in RNA-Seq analysis. False Discovery Rate (FDR) is the proportion of false positives among all positives.
535 differential expressed genes were detected with False Discovery Rate (FDR) corrected p-value < 0:05 and fold-change > 1:5 comparing myofibroblast to control group.
The 535 genes were further explored functional annotation clusters and pathway enrichment analyses. Biological processes related to immune responses are the top terms enriched for this gene list.
SiRNA treatment of human and mouse cells
Monocytes are difficult to transfect99. As monocytes are non-proliferating cells99, plasmid-based siRNA platforms that require entry into the nucleus are also unlikely to inhibit protein production across the entire population.
Accell Silencing RNA (siRNA, Dharmacon, Lafayette, CO) is specifically engineered to penetrate cells without the use of transfection reagents, due to a cholesterol modification at one terminus of the sequence. Additionally, Accell siRNA is well suited for a study for silencing proteins in otherwise non-proliferating cells, because each non-replicating siRNA molecule does not have to enter the nucleus, a necessary prerequisite for plasmid-based siRNA platforms. Thus, Accell siRNA targets all cells in a population, not just those that are replicating. Further, that the Accell siRNA does not replicate inside a cell is ideal from a dose-recovery standpoint. Lastly, Accell siRNA contains a modification that inhibits enzymatic digestion within cells. Thus, Accell siRNA made a natural choice for this experiment, and all siRNAs used in this study are Accell siRNA. siRNA was resuspended in RNAse-free ultrapure water, as per the manufacturer’s instructions.
The SMARTpool of siRNA targeting human talin2 contained four target sequences, while the anti-parallel corresponding siRNA sequences actually silenced the mRNA. Target sequences: #1 (CCAGAAAACUGAACGAUUA), #2 (GCCCUGUCCUUAAAGAUUU), #3 (CGACUGUGGUUAAAUACUC), and #4 (CGAGAAAGCUUGUGAGUUU). The SMARTpool targeting mouse talin2 also contained four target sequences: #1 (CGACUGUGGUUAAAUACUC), #2 (CUGGAAAAUUCAGUGAUGA), #3 (CCCUGGAUUUUGAAGAACA), and #4 (CCAUCGAGUACAUAAAACA). Control non-targeting SMARTpools contained 4 target sequences, including: #1 (CCAGAAAACUGAACGAUUA), #2 (GCCCUGUCCUUAAAGAUUU), #3 (CGACUGUGGUUAAAUACUC), and #4 (CGAGAAAGCUUGUGAGUUU). Individual siRNA tested in mice was talin2 siRNA #2, target: 5’ CUGGAAAAUUCAGUGAUGA 3’, anti-sense: 5’ UCAUCACUGAAUUUUCCAG 3’. Control non-targeting siRNA used in the mouse in vivo study was (CCAGAAAACUGAACGAUUA).
Flow cytometry
Myofibroblasts were removed from their tissue culture surfaces by the use of cold trypsin-EDTA (Sigma), followed by mechanical agitation by a rubber policeman. Cells were fixed and permeabilized using Cytofix/Cytoperm (BD biosciences, Franklin Lakes, NJ). Live dead stain was live-dead aqua, used per manufacturer’s instructions (ThermoFisher). Compensation was performed via UltraComp beads (ThermoFisher) per the manufacturer’s instructions. Antibodies used were anti-collagen I (Biolegend), anti-alpha smooth muscle actin (αSMA) (R and D systems), anti ki-67 (BD biosciences), and anti-talin2 (R and D systems).
SiRNA penetration into cells
siRNA complexed to AF-488 (Dharmacon) was added to human and mouse monocytes and fibroblasts at 50 nM, to determine if the siRNA was penetrating the cell and remaining for up to 2 days. Cells were removed from their culture surface as previously indicated, fixed, and analyzed via flow cytometry.
Mouse lung instillation
Male and female mice were acquired at 8 weeks of age (Jackson laboratories, Bar Harbor, ME) with the intent to be used at 12 weeks of age. However, due to delays regarding COVID19 lockdown and the allowed resumption of non-COVID19 research, the mice were 32 weeks old when the study progressed. Mouse lungs were instilled with bleomycin (0.075 units, 75 µg, Fresenius Kabi, Switzerland) and siRNA (0.2 µM), suspended in endotoxin-free PBS, as previously described100, following a protocol approved by the UChicago IACUC (protocol 72470, lung fibrosis study approved on 4/10/2019). Control mice were instilled with PBS alone.
First, mice were anesthetized via isoflurane inhalation (2%). Mice were then placed upright on an angled surface, their tongue pulled to the side, and a 200 mL narrow pipet was placed at the entrance of their throat. 50 mL of PBS (alone, with bleomycin, or with siRNA) was dispensed to the entrance of the throat, and mice were allowed to inhale. Administration to the lungs was confirmed by listening to the mouse’s breathing for popping noises. Mice were then weighed and placed on a heating pad to recuperate.
Following bleomycin insult, mice were treated with siRNA using an identical installation procedure. The dose schedule was 7, 9, 11, 14, 16, and 18 days following bleomycin insult. Mice were euthanized at 21 days post insult via injecting of euthasol (Covetrus, Portland, ME) instead of CO2 inhalation, which could damage the lungs.
Lung resection and fibrosis scoring
Lungs were resected, and perfused with 5 mL of PBS via cardiac puncture. BAL involved exposing the trachea and penetrating the trachea using an 18 gauge needle. The BAL was performed by inserting a catheter needle (Exel International, Redondo Beach, CA) into the trachea, and slowly moving 800 mL of PBS into and out of the lungs. If blood entered the lavage it was discarded. In a previous pilot study testing the efficacy of talin2 siRNA treatment, the lungs were then broncheo-alveolar lavaged (BAL), and the resulting lavage frozen in 10% DMSO. Due to limitations associated with COVID19 scheduling, BAL was not possible in the larger study. This lavage was thawed, fixed, immunostained, and analyzed via flow cytometry to provide the data on talin2’s in vivo reduction following siRNA treatment.
After resection, the right and left lobes were separated. The left lobe was fixed in 4% paraformaldehyde overnight, mounted in paraffin, sectioned into 5 mm slices, and stained using Masson’s trichrome. Stained lungs were scanned at high resolution using a CRi Panoramic SCAN 40x Whole Slide Scanner (Perkin-Elmer, Waltham, MA), and were read for fibrosis using a modified Ashcroft method, as previously described63.
The right lobe of the lung was frozen, and dehydrated using a tissue lyophilizer (Labconco, Kansas City, MO). This dehydrated lung was weighed, and was assessed for collagen content by hydroxyproline assay, as previously described101. Briefly, dried lungs were digested in 6 N HCl/PBS at 100 C for 24 h. The supernatant from this digestion was added to 96 well plates and treated sequentially with chloramine-T solution and Ehrlich’s solution at 65 C for 15 min to facilitate the color change reaction. Color was read at 561 nm. Quantification was provided by use of a hydroxyproline (Sigma) dilution series, which was transformed into a standard curve.
Tln2 -/- mice
Tln2-/- mice (MGI 1917799) were the kind gift of Drs. Roy Zent and David Critchley. Mouse genomic DNA was isolated through ear punches, and Tln2-/- mice were validated genomic DNA by PCR (Transnetyx).
Kidney unilateral ureteral obstruction (UUO) fibrosis model
UUO surgery was performed as previously described102, with adjustments, following a protocol approved by the UChicago IACUC (protocol 72470, kidney fibrosis study approved on 2/17/2021). Briefly, mice were anesthetized via 2% isoflurane inhalation, and injected with meloxicam (1 mg/kg), buprenorphine (0.1 mg/kg) in a saline solution, subcutaneously. Mice were laid on their right side and an abdominal incision used to visualize the left ureter. The left ureter was ligated in the middle section of the ureter with two ties (2 mm apart) using 7 − 0 silk sutures. Peritoneum is then closed with 5 − 0 vicryl and skin is closed with 5 − 0 nylon. Mice were observed for distress following the surgery. Mice were also weighed daily to make sure that weight loss did not exceed 10% of body weight, and in all cases mice continued to gain weight following the UUO surgery. This weight gain was not significantly different between healthy mice and mice following UUO surgery. 14 days following the UUO ligation, the mice were euthanized via injection with sodium pentobarbitol, and the kidneys resected. For all of the mice, the UUO ligation was still in place following resection.
Assessment of fibrosis in kidneys
Right and left kidneys were placed in 4% PFA for 24 h, mounted in paraffin, sectioned into 5 μm full kidney slices, and stained using immunohistochemistry for collagen I (1:4000, polyclonal rabbit, lifespan biosciences, Seattle WA) via a Bond-Max autostaining system (Leica biosystems, Lincolnshire, IL). Stained kidneys were scanned at high resolution using a CRi Panoramic SCAN 40x Whole Slide Scanner (Perkin-Elmer).
Images were equalized in size, and converted to .tif files using CaseViewer. Images were then imported into imageJ, scale set for conversion between microns and pixels, and deconvoluted with the “H DAB” deconvolution option. The blue stain was thresholded at 215 to see how many pixels were positive for nuclear staining but negative for collagen I, and the brown (IHC positive) image thresholded at 185 to see how many pixels were positive for collagen I. Machine-staining allowed these kidneys to be compared with high reproducibility.
Blood analysis for markers of kidney damage
At the time of euthanasia, blood was collected via submandibular bleed into protein low-bind tubes and allowed to coagulate for 2 h on ice. Coagulated blood was then centrifuged at 10,000 xg for 10 min, and serum collected. Serum was then diluted 4x in MilliQ water before being placed on deck on an Alfa Wassermann VetAxcel Blood Chemistry Analyzer. All tests requiring calibration were calibrated on the day of analysis and quality controls were run before analyzing samples. Serum tests were run according to kit instructions, and creatinine kinase was normalized to calcium ion concentrations where indicated to account for sample hemolysis.
Immunofluorescence
Human monocytes from 3 donors were cultured in 8-well chamber slides (Millipore-Sigma) in SFM, and allowed to become myofibroblasts over 3 days. Myofibroblasts were then treated with SMARTpools for non-targeting control siRNA and siRNA targeting human talin2. After 1 week of de-differentiation, the slides were dried quickly using the airflow from a laminar flow hood, in order to preserve cellular morphology as accurately as possible. Cells were then fixed with ice cold 4% PFA, permeabilized with saponin (Sigma). Primary antibodies (anti-talin2, novus) were added at 5 mg/mL overnight. Cells were washed 3x in PBS, and were exposed to DAPI and F-actin-phalloidin-488 (ThermoFisher) for 1 min. Cells were mounted using water-based mowiol mounting media (Southern Biotech, Birmingham, AL) to preserve fluorescence. Slides were imaged immediately using a confocal microscope (Olympus, Shinjuku City, Tokyo).
Legendplex ELISA
Supernatant from experiments involving culturing human and mouse monocytes and fibroblasts into myofibroblasts on surfaces of different stiffnesses and treatment with siRNA were thawed at 4 C, and centrifuged at 4 C and 10,000 xg force to pellet cell debris. Supernatant from 96 well culture plates was taken and analyzed in triplicate. Legendplex (Biolegend) beads against general inflammation markers were added, according to the manufacturer’s instructions. Sample readouts were normalized to each individual experiment and donor control to account for donor variability.
Statistical analysis
Statistical analyses were performed using GraphPad Prism software, and P < 0.05 was considered statistically significant. 2-way ANOVA and 1-way ANOVA using Dunnett’s post-test for multiple comparisons were used to compare groups. Student’s t-test was used to make direct comparisons between two data sets. 1-way ANOVA with a trend post-test was used to compare the trend relationships in Figs. 1, 2 and 3 regarding culturing cells on surfaces of increasing stiffness.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Abbreviations
- αSMA:
-
Alpha-smooth muscle actin
- ALT:
-
Alanine transferase
- AST:
-
Aspartate transferase
- BAL:
-
Broncheo-alveolar lavaged
- BUN:
-
Blood urea nitrogen
- FCS:
-
Fetal calf serum
- FAs:
-
Focal adhesions
- FBs:
-
Fibrillar adhesions
- IPF:
-
Idiopathic pulmonary fibrosis
- IHC:
-
Immunohistochemistry
- MRC-5:
-
Human fibroblasts
- kPa:
-
Kilopascals
- MCP1:
-
Macrophage-chemotactic protein-1
- NIH-3T3:
-
Mouse fibroblasts
- PBMC:
-
Peripheral blood mononuclear cells
- PBS:
-
Phosphate buffered saline
- SFM:
-
Serum-free media
- siRNA:
-
Silencing RNA
- TGFβ:
-
Transforming growth factor β
- Tln2:
-
Talin2
- TNFα:
-
Tumor necrosis factor α
- UUO:
-
Unilateral ureteral obstruction
References
Wynn, T. A. Cellular and molecular mechanisms of fibrosis. J. Pathol. 214(2), 199–210 (2008).
Wynn, T. A. Fibrotic disease and the T(H)1/T(H)2 paradigm. Nat. Rev. Immunol. 4(8), 583–594 (2004).
Spagnolo, P. Novel treatments for idiopathic pulmonary fibrosis. Am. J. Med. (2015).
Xaubet, A., Serrano-Mollar, A. & Ancochea, J. Pirfenidone for the treatment of idiopathic pulmonary fibrosis. Expert Opin. Pharmacother. 15(2), 275–281 (2014).
Knuppel, L. et al. A novel antifibrotic mechanism of nintedanib and Pirfenidone. Inhibition of collagen fibril assembly. Am. J. Respir Cell. Mol. Biol. 57(1), 77–90 (2017).
Raghu, G. et al. Long-term treatment with Recombinant human pentraxin 2 protein in patients with idiopathic pulmonary fibrosis: an open-label extension study. Lancet Respir Med. 7(8), 657–664 (2019).
Madsen, D. H. et al. M2-like macrophages are responsible for collagen degradation through a mannose receptor-mediated pathway. J. Cell. Biol. 202(6), 951–966 (2013).
Bianchetti, L. et al. Extracellular matrix remodelling properties of human fibrocytes. J. Cell. Mol. Med. 16(3), 483–495 (2012).
Hinz, B. & Gabbiani, G. Fibrosis: recent advances in myofibroblast biology and new therapeutic perspectives. F1000 Biol. Rep. 2, 78 (2010).
Hinz, B. Mechanical aspects of lung fibrosis: a spotlight on the myofibroblast. Proc. Am. Thorac. Soc. 9(3), 137–147 (2012).
Klingberg, F., Hinz, B. & White, E. S. The myofibroblast matrix: implications for tissue repair and fibrosis. J. Pathol. 229(2), 298–309 (2013).
Hinz, B. Formation and function of the myofibroblast during tissue repair. J. Invest. Dermatol. 127(3), 526–537 (2007).
Liu, F. et al. Feedback amplification of fibrosis through matrix stiffening and COX-2 suppression. J. Cell. Biol. 190(4), 693–706 (2010).
Tomasek, J. J. et al. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell. Biol. 3(5), 349–363 (2002).
Abe, R. et al. Peripheral blood fibrocytes: differentiation pathway and migration to wound sites. J. Immunol. 166(12), 7556–7562 (2001).
Forbes, S. J. et al. A significant proportion of myofibroblasts are of bone marrow origin in human liver fibrosis. Gastroenterology 126(4), 955–963 (2004).
Direkze, N. C. et al. Multiple organ engraftment by bone-marrow-derived myofibroblasts and fibroblasts in bone-marrow-transplanted mice. Stem Cells. 21(5), 514–520 (2003).
Hashimoto, N. et al. Bone marrow-derived progenitor cells in pulmonary fibrosis. J. Clin. Invest. 113(2), 243–252 (2004).
Nakashima, T. et al. Lung bone marrow-derived hematopoietic progenitor cells enhance pulmonary fibrosis. Am. J. Respir Crit. Care Med. 188(8), 976–984 (2013).
Mori, L. et al. Fibrocytes contribute to the myofibroblast population in wounded skin and originate from the bone marrow. Exp. Cell. Res. 304(1), 81–90 (2005).
Lagares, D. et al. Targeted apoptosis of myofibroblasts with the BH3 mimetic ABT-263 reverses established fibrosis. Sci. Transl Med., 9(420). (2017).
Balestrini, J. L. et al. The mechanical memory of lung myofibroblasts. Integr. Biol. (Camb). 4(4), 410–421 (2012).
Hoffman, B. D., Grashoff, C. & Schwartz, M. A. Dynamic molecular processes mediate cellular mechanotransduction. Nature. 475(7356), 316–323. (2011).
Geiger, B., Spatz, J. P. & Bershadsky, A. D. Environmental sensing through focal adhesions. Nat. Rev. Mol. Cell. Biol. 10(1), 21–33 (2009).
Roca-Cusachs, P. et al. Clustering of alpha(5)beta(1) integrins determines adhesion strength whereas alpha(v)beta(3) and Talin enable mechanotransduction. Proc. Natl. Acad. Sci. U S A. 106(38), 16245–16250 (2009).
Dugina, V. et al. Focal adhesion features during myofibroblastic differentiation are controlled by intracellular and extracellular factors. J. Cell. Sci. 114(Pt 18), 3285–3296 (2001).
Praekelt, U. et al. New isoform-specific monoclonal antibodies reveal different sub-cellular localisations for talin1 and talin2. Eur. J. Cell. Biol. 91(3), 180–191 (2012).
Morton, P. E. & Parsons, M. Dissecting cell adhesion architecture using advanced imaging techniques. Cell. Adh Migr. 5(4), 351–359 (2011).
Debrand, E. et al. Mice carrying a complete deletion of the talin2 coding sequence are viable and fertile. Biochem. Biophys. Res. Commun. 426(2), 190–195 (2012).
Anthis, N. J. et al. Structural diversity in integrin/talin interactions. Structure 18(12), 1654–1666 (2010).
Critchley, D. R. Biochemical and structural properties of the integrin-associated cytoskeletal protein Talin. Annu. Rev. Biophys. 38, 235–254 (2009).
Debrand, E. et al. Talin 2 is a large and complex gene encoding multiple transcripts and protein isoforms. FEBS J. 276(6), 1610–1628 (2009).
Manso, A. M. et al. Talin1 has unique expression versus Talin 2 in the heart and modifies the hypertrophic response to pressure overload. J. Biol. Chem. 288(6), 4252–4264 (2013).
Gough, R. E. & Goult, B. T. The Tale of two talins - two isoforms to fine-tune integrin signalling. FEBS Lett. 592(12), 2108–2125 (2018).
Anthis, N. J. et al. The structure of an integrin/talin complex reveals the basis of inside-out signal transduction. EMBO J. 28(22), 3623–3632 (2009).
Austen, K. et al. Extracellular rigidity sensing by Talin isoform-specific mechanical linkages. Nat. Cell. Biol. 17(12), 1597–1606 (2015).
Haining, A. W. et al. All subdomains of the Talin rod are mechanically vulnerable and May contribute to cellular mechanosensing. ACS Nano. 10(7), 6648–6658 (2016).
Monkley, S. J. et al. Disruption of the Talin gene arrests mouse development at the gastrulation stage. Dev. Dyn. 219(4), 560–574 (2000).
Chen, N. T. & Lo, S. H. The N-terminal half of talin2 is sufficient for mouse development and survival. Biochem. Biophys. Res. Commun. 337(2), 670–676 (2005).
Kanchanawong, P. et al. Nanoscale architecture of integrin-based cell adhesions. Nature 468(7323), 580–584 (2010).
del Rio, A. et al. Stretching single Talin rod molecules activates vinculin binding. Science 323(5914), 638–641 (2009).
Liu, J. et al. Talin determines the nanoscale architecture of focal adhesions. Proc. Natl. Acad. Sci. U S A. 112(35), E4864–E4873 (2015).
Goksoy, E. et al. Structural basis for the autoinhibition of Talin in regulating integrin activation. Mol. Cell. 31(1), 124–133 (2008).
Ingmar Schoen, B. L. P. & Vogel, V. The Yin-Yang of rigidity sensing: how forces and mechanical properties regulate the cellular response to materials. Annu. Rev. Mater. Sci. 43, 589–618 (2013).
Hytonen, V. P. & Vogel, V. How force might activate Talin’s vinculin binding sites: SMD reveals a structural mechanism. PLoS Comput. Biol. 4(2), e24 (2008).
Wells, R. G. The role of matrix stiffness in regulating cell behavior. Hepatology 47(4), 1394–1400 (2008).
Handorf, A. M. et al. Tissue stiffness dictates development, homeostasis, and disease progression. Organogenesis 11(1), 1–15 (2015).
Booth, A. J. et al. Acellular normal and fibrotic human lung matrices as a culture system for in vitro investigation. Am. J. Respir Crit. Care Med. 186(9), 866–876 (2012).
Islamoglu, M. S., Gulcicek, S. & Seyahi, N. Kidney tissue elastography and interstitial fibrosis observed in kidney biopsy. Ren. Fail. 44(1), 314–319 (2022).
Neumark, N. et al. The idiopathic pulmonary fibrosis cell atlas. Am. J. Physiol. Lung Cell. Mol. Physiol. 319(6), L887–L893 (2020).
White, M.J.V., et al., Blocking antibodies against integrin-α3, -αM, and -αMβ2 de-differentiate myofibroblasts, and improve lung fibrosis and kidney fibrosis. Scientific Reports, 2024. 14(1): p. 21623.
Shi, J. et al. Anti-fibrotic actions of interleukin-10 against hypertrophic scarring by activation of PI3K/AKT and STAT3 signaling pathways in scar-forming fibroblasts. PLoS One. 9(5), e98228 (2014).
Gasse, P. et al. IL-1 and IL-23 mediate early IL-17A production in pulmonary inflammation leading to late fibrosis. PLoS One. 6(8), e23185 (2011).
Yogo, Y. et al. Macrophage derived chemokine (CCL22), thymus and activation-regulated chemokine (CCL17), and CCR4 in idiopathic pulmonary fibrosis. Respir Res. 10, 80 (2009).
Le, T. T. et al. Blockade of IL-6 trans signaling attenuates pulmonary fibrosis. J. Immunol. 193(7), 3755–3768 (2014).
Belperio, J. A. et al. The role of the Th2 CC chemokine ligand CCL17 in pulmonary fibrosis. J. Immunol. 173(7), 4692–4698 (2004).
Huaux, F. et al. A profibrotic function of IL-12p40 in experimental pulmonary fibrosis. J. Immunol. 169(5), 2653–2661 (2002).
Sahin, H. & Wasmuth, H. E. Chemokines in tissue fibrosis. Biochim. Biophys. Acta. 1832(7), 1041–1048 (2013).
Oikonomou, N. et al. Soluble TNF mediates the transition from pulmonary inflammation to fibrosis. PLoS One. 1, e108 (2006).
Borthwick, L. A. The IL-1 cytokine family and its role in inflammation and fibrosis in the lung. Semin Immunopathol. 38(4), 517–534 (2016).
Gallucci, R. M., Lee, E. G. & Tomasek, J. J. IL-6 modulates alpha-smooth muscle actin expression in dermal fibroblasts from IL-6-deficient mice. J. Invest. Dermatol. 126(3), 561–568 (2006).
Iyonaga, K. et al. Monocyte chemoattractant protein-1 in idiopathic pulmonary fibrosis and other interstitial lung diseases. Hum. Pathol. 25(5), 455–463 (1994).
Hubner, R. H. et al. Standardized quantification of pulmonary fibrosis in histological samples. Biotechniques 44(4), 507–511 (2008).
Ray, L. et al. A comparative study of serum aminotransferases in chronic kidney disease with and without end-stage renal disease: need for new reference ranges. Int. J. Appl. Basic. Med. Res. 5(1), 31–35 (2015).
Hinz, B. et al. The myofibroblast: one function, multiple origins. Am. J. Pathol. 170(6), 1807–1816 (2007).
Pinon, P. et al. Talin-bound NPLY motif recruits integrin-signaling adapters to regulate cell spreading and mechanosensing. J. Cell. Biol. 205(2), 265–281 (2014).
Discher, D. E., Janmey, P. & Wang, Y. L. Tissue cells feel and respond to the stiffness of their substrate. Science 310(5751), 1139–1143 (2005).
Yeung, T. et al. Effects of substrate stiffness on cell morphology, cytoskeletal structure, and adhesion. Cell. Motil. Cytoskeleton. 60(1), 24–34 (2005).
Hinz, B. Masters and servants of the force: the role of matrix adhesions in myofibroblast force perception and transmission. Eur. J. Cell. Biol. 85(3–4), 175–181 (2006).
Goffin, J. M. et al. Focal adhesion size controls tension-dependent recruitment of alpha-smooth muscle actin to stress fibers. J. Cell. Biol. 172(2), 259–268 (2006).
Wells, R. G. The role of matrix stiffness in hepatic stellate cell activation and liver fibrosis. J. Clin. Gastroenterol. 39(4 Suppl 2), S158–S161 (2005).
Tarasova, N. K. et al. Proteomics reveals a role for attachment in monocyte differentiation into efficient Proinflammatory macrophages. J. Proteome Res. 14(9), 3940–3947 (2015).
Lawson, C. et al. FAK promotes recruitment of Talin to nascent adhesions to control cell motility. J. Cell. Biol. 196(2), 223–232 (2012).
Pilling, D. et al. TNF-alpha-stimulated fibroblasts secrete lumican to promote fibrocyte differentiation. Proc. Natl. Acad. Sci. U S A. 112(38), 11929–11934 (2015).
Ashley, S. L. et al. Periostin regulates fibrocyte function to promote myofibroblast differentiation and lung fibrosis. Mucosal Immunol. 10(2), 341–351 (2017).
Pilling, R. G. D. Fibrocytes: New Insights into Tissue Repair, in Fibrocytes: New Insights into Tissue Repair (eds Bucala, R.) 268 (Yale University Press, 2007). Editor.
Lim, J. et al. An essential role for Talin during alpha(M)beta(2)-mediated phagocytosis. Mol. Biol. Cell. 18(3), 976–985 (2007).
Qi, L. et al. Talin2-mediated traction force drives matrix degradation and cell invasion. J. Cell. Sci. 129(19), 3661–3674 (2016).
Zaidel-Bar, R. et al. Functional atlas of the integrin adhesome. Nat. Cell. Biol. 9(8), 858–867 (2007).
Zhang, X. et al. Talin depletion reveals independence of initial cell spreading from integrin activation and traction. Nat. Cell. Biol. 10(9), 1062–1068 (2008).
Theodosiou, M. et al. Kindlin-2 cooperates with Talin to activate integrins and induces cell spreading by directly binding paxillin. Elife 5, e10130 (2016).
Muiznieks, L. D. & Keeley, F. W. Molecular assembly and mechanical properties of the extracellular matrix: A fibrous protein perspective. Biochim. Biophys. Acta. 1832(7), 866–875 (2013).
Eckes, B. & Krieg, T. Regulation of connective tissue homeostasis in the skin by mechanical forces. Clin. Exp. Rheumatol. 22(3 Suppl 33), S73–S76 (2004).
Marenzana, M. et al. The origins and regulation of tissue tension: identification of collagen tension-fixation process in vitro. Exp. Cell. Res. 312(4), 423–433 (2006).
Marinkovic, A., Liu, F. & Tschumperlin, D. J. Matrices of physiologic stiffness potently inactivate idiopathic pulmonary fibrosis fibroblasts. Am. J. Respir Cell. Mol. Biol. 48(4), 422–430 (2013).
Lim, I. R. et al. Talin modulation by a synthetic N-Acylurea derivative reduces angiogenesis in human endothelial cells. Int. J. Mol. Sci., 18(1). (2017).
Thanki, K. et al. Formulation of RNA interference-based drugs for pulmonary delivery: challenges and opportunities. Ther. Deliv. 9(10), 731–749 (2018).
Zafra, M. P. et al. Gene Silencing of SOCS3 by SiRNA intranasal delivery inhibits asthma phenotype in mice. PLoS One. 9(3), e91996 (2014).
Pilling, D., Vakil, V. & Gomer, R. H. Improved serum-free culture conditions for the differentiation of human and murine fibrocytes. J. Immunol. Methods. 351(1–2), 62–70 (2009).
White, M. J. & Gomer, R. H. Trypsin, tryptase, and thrombin polarize macrophages towards a Pro-Fibrotic M2a phenotype. PLoS One. 10(9), e0138748 (2015).
Crawford, J. R., Pilling, D. & Gomer, R. H. Improved serum-free culture conditions for spleen-derived murine fibrocytes. J. Immunol. Methods. 363(1), 9–20 (2010).
Hinz, B. et al. Myofibroblast development is characterized by specific cell-cell adherens junctions. Mol. Biol. Cell. 15(9), 4310–4320 (2004).
Baum, J. & Duffy, H. S. Fibroblasts and myofibroblasts: what are we talking about? J. Cardiovasc. Pharmacol. 57(4), 376–379 (2011).
Rio, D. C. et al. Purification of RNA using trizol (TRI reagent). Cold Spring Harb Protoc. 2010(6), pdbprot5439 (2010).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29(1), 15–21 (2013).
Liao, Y., Smyth, G. K. & Shi, W. FeatureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30(7), 923–930 (2014).
Robinson, M. D., McCarthy, D. J. & Smyth, G. K. EdgeR: a bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26(1), 139–140 (2010).
Huang, D. W. et al. DAVID bioinformatics resources: expanded annotation database and novel algorithms to better extract biology from large gene lists. Nucleic Acids Res.. 35(Web Server issue), W169–W175 (2007).
Moradian, H. et al. mRNA Transfection-Induced activation of primary human monocytes and macrophages: dependence on carrier system and nucleotide modification. Sci. Rep. 10(1), 4181 (2020).
Katsumata, K. et al. Targeting inflammatory sites through collagen affinity enhances the therapeutic efficacy of anti-inflammatory antibodies. Sci. Adv. 5(11), eaay1971 (2019).
Bergheim, I. et al. Critical role of plasminogen activator inhibitor-1 in cholestatic liver injury and fibrosis. J. Pharmacol. Exp. Ther. 316(2), 592–600 (2006).
Cochrane, A. L. et al. Renal structural and functional repair in a mouse model of reversal of ureteral obstruction. J. Am. Soc. Nephrol. 16(12), 3623–3630 (2005).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. U S A. 102(43), 15545–15550 (2005).
Acknowledgements
We thank the Human Tissue Resource Center of the University of Chicago for histology analysis. We thank the Integrated Light Microscopy Core of the University of Chicago for Imaging. We thank the Genomics core at the University of Chicago for RNAseq analysis. We also acknowledge blind scoring from Margo MacDonald, and guidance on fibrosis models from Dr. Anne Sperling.
Funding
This work was supported in part by the University of Chicago (to JAH).
Author information
Authors and Affiliations
Contributions
Conceptualization: MJVW, JAH Methodology: MJVW. Investigation: MJVW, MO, JEGM, AS. Surgical Lead: AS. Visualization: MJVW. Funding acquisition: JAH. Project administration: MJVW, JAH. Supervision: MJVW, JAH. Writing – original draft: MJVW. Writing – review & editing: MJVW, JAH.
Corresponding author
Ethics declarations
Ethics approval
All mouse experiments were performed under supervision with protocols approved by the University of Chicago IACUC, and this study was performed in accordance with the relevant guidelines and regulations. The results of these experiments are reported in this manuscript using ARRIVE guidelines.
Competing interests
MJVW and JAH are inventors on U.S. Patent WO2022256824A1. The other authors declare that they have no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
White, M.J.V., Ozkan, M., Medellin, J.E.G. et al. Inhibition of Talin2 dedifferentiates myofibroblasts and reverses lung and kidney fibrosis. Sci Rep 15, 18010 (2025). https://doi.org/10.1038/s41598-025-00939-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-025-00939-x
