
Abstract
Newly acquired skill memory can generalize/transfer to the untrained arm. Such interlimb generalization of a learned skill has been shown to be symmetric in our previous work and can be influenced by cognitive processes that emerge during skill learning. However, it is unknown whether engaging in other cognitively demanding tasks following skill acquisition can influence skill generalization. Our research goal was to uncover how a secondary task, involving working memory, interacts with a newly formed skill memory and influences subsequent interlimb generalization. To test this idea, we conducted a set of three experiments by recruiting right-handed young healthy individuals (N = 92) who learned a novel motor skill (long or short training on a skilled reaching task) followed by performing a working memory or control task with the right arm. Finally, all individuals were tested for immediate or delayed (after 24 h) interlimb skill generalization to the untrained left arm. We found significant immediate as well as delayed generalization in individuals who received long training on the motor skill task, irrespective of whether they performed working memory or control task. On the other hand, performing the working memory but not control task following short skill training impaired generalization when the untrained arm was tested 24 h later. These findings indicate that short training reflecting early stages of skill learning and the subsequent skill memory stabilization are dependent on working memory such that the underlying neural interactions mediating these processes can have implications for skill generalization.
Similar content being viewed by others

Introduction
Motor learning is a unique aspect of human behavior and a critical research area in psychology and neuroscience. Typically, motor learning abilities are characterized by acquisition of novel skilled movements (motor skills) and refinement of the existing ones (motor adaptation). Motor skill learning leads to reduced movement errors, improved movement speed and accuracy, and eventually results into stable memory formation of the newly learned movements1,2,3,4,5. On the other hand, learning during motor adaptation requires correcting for movement errors resulting from an external perturbation6,7,8. Such human behavior pertaining to skill learning as well as motor adaptation can be considered robust if such abilities can generalize or transfer to untrained contexts9,10. Understanding how learning generalizes to unpracticed conditions can provide deeper insights into the underlying human motor learning mechanisms.
One form of generalization that specifically interests us, given its relevance for motor rehabilitation, is interlimb generalization in which learning acquired using one effector such as the arm is transferred to the other untrained arm. Studies have shown that such interlimb generalization is possible, but it is variable and influenced by task conditions11 and individual movement characteristics12. For example, many studies have shown that generalization of motor adaptation is asymmetric and unidirectional, i.e., from non-dominant arm to the dominant arm13,14,15,16,17,18. However, factors such as workspace ___location19,20, handedness21,22,23, movement speed23, magnitude of the angular rotation24 and the amount of training18 can influence the magnitude and direction of such interlimb generalization of motor adaptation. More specifically and relevant for our work, Poh and colleagues20 found that explicit components developed during visuomotor adaptation can generalize across arms giving rise to symmetric transfer. The transfer of implicit components of such learning however was limited and depended on the joint-based compatibility of the workspace for the two arms. Interestingly, in the context of our previous work on interlimb generalization of newly learned motor skills, we also found that explicit/strategic components acquired during training were transferred to the untrained arm giving rise to symmetric, bidirectional (i.e., from non-dominant to dominant arm and vice versa) skill generalization25. However, factors that can influence such generalization of a newly acquired motor skill memory to the untrained arm (specially the non-dominant arm) are poorly understood. Intrigued by this, we asked a simple question- can generalization of a newly acquired skill memory be impacted by performing a post-skill training secondary task that is cognitively demanding?
Skill motor behavior is dependent on formation and stabilization of motor memories. However, newly formed memories are unstable, and liable to disruption and interference1,26,27,28. Studies exploring the fate of a memory post-training suggest that instability of the memory provides a time window for interaction that may allow interference and impact information generalization29,30,31. On the other hand, memory stability can be enhanced by increasing training duration as experts who have mastered a skill train repeatedly to stabilize that skill and make it less vulnerable to disruption by interference. Based on such an interplay between memory stability and interference, we probed whether skill generalization of a newly acquired motor memory after a long versus short training is influenced by engaging in a secondary memory task (such as working memory). To test this idea, we conducted a series of three different experiments in which young healthy individuals trained on a novel motor skill task (variable task structure requiring fast and accurate movement to random targets) using their dominant right arm. Following training, one set of individuals performed a visuo-spatial working memory task known to be mediated by neural substrate such as dorsolateral prefrontal cortex (DLPFC) which is interestingly also involved in memory stabilization following variable skill training32,33,34,35,36. The other set of individuals performed a control task with no working memory requirement with their right arm. Finally, we tested the untrained left arm of all individuals for immediate or delayed (24 h later) interlimb skill generalization to help us uncover interactions between newly formed skill memory resulting from long versus short training and a cognitively engaging secondary task of working memory. Our results showed significant immediate and delayed generalization after long skill training regardless of the secondary tasks, but performing the working memory task after short training impaired generalization when untrained arm was tested 24 h later.
Methods
Participants
A total of 92 young healthy individuals with normal or corrected-to-normal vision (age range: 18–38 years, mean ± SD: 22.663 ± 3.61 years, 38 Females) successfully completed the study. All participants gave written informed consent. This study was approved by the Institute Ethics Committee of the Indian Institute of Technology Hyderabad and was conducted according to the principles of the Declaration of Helsinki. All participants were right-handed (Edinburgh Handedness Inventory37). No individual reported any history of neurological, medical, or psychiatric abnormality before taking part in the study. All participants were given monetary compensation for taking part in this study.
Study apparatus
The experimental setup consisted of a digitizing tablet (GTCO CalComp) placed on a frame over which a computer screen (1920 × 1080 pixels) was mounted horizontally with a reflective mirror placed in-between (Fig. 1a). Participants sat on an adjustable chair in front of this setup and the height of the chair was adjusted for every participant so that they could clearly see the stimuli reflecting on the mirror. Direct vision of the hand was blocked and a yellow dot (2 mm) indicating ___location of the handheld stylus was provided to guide participant’s movement during the experiment. The hand position was recorded at 120 Hz sampling rate on the digitizing tablet. The tasks were designed using Psychtoolbox on MATLAB38.
Task Procedure
Motor skill task
In the beginning of the experiment all subjects had a brief familiarization session about the setup and the task. For the motor skill task (Fig. 1b), participants were asked to perform fast and accurate planar reaching movement between a start circle (green circle, 10 mm diameter) and a target circle (blue circle, 10 mm diameter) with the right hand holding the stylus on the digitizing tablet. Each trial started with the participant moving his/her right hand to the start circle. After 500 ms at this start circle, an audio cue (beep) was presented, and one of five target circles appeared (pseudorandomized) on the screen. These two circles were 150 mm apart and connected with parallel yellow lines/path. Participants were instructed to move to the target circle as soon as they heard the audio cue while trying to remain within the yellow path. The yellow dot indicating the hand position was visible throughout the movement. Thus, to make fast and accurate skilled movements on this task participants were explicitly told to finish the movement as fast as possible (i.e., within 650 ms based on our pilot data; indicating temporal accuracy) and reach the target circle while moving within the yellow path (indicating spatial accuracy). Trial duration was 2 s and upon completion, movement performance feedback was provided through visual cues and a numerical score- visual cues comprised of a red line indicating part of the movement trajectory that was outside the specified yellow path (presence of this red line indicated low spatial accuracy or high spatial error), and a red square indicating hand position at 650 ms (farther this red square from the target circle, lower the temporal accuracy or higher the temporal error).
Based on these two performance feedback, we computed a single numeric “motor error” score which was simultaneously presented as an additional feedback at the top corner of the screen at the end of each movement. This motor error was a cumulative sum of spatial error and temporal error which were calculated using position coordinates. Data points from the red line were used to calculate the path length (in mm) that was outside the specified path (indicating spatial error). On the other hand, the red square provided hand position at 650 ms that was used to compute the distance (in mm) of the hand from the target circle (indicating temporal error). Finally, both these error values (in mm) were summed to calculate the motor error (in mm). While participants were not told about how the motor error was calculated (to avoid any bias towards one or the other error components), they were instructed to make fast and accurate movement that will lead to lower motor error. Thus, for instance, if the participant successfully completed the movement on a given trail by reaching the target circle within 650 ms and within the specified path, the motor error on that trial will be close to 0. These feedback (visual cues and motor error score) were presented on the screen at the end of each trial. Total trial duration was 2 s. Note that the motor error score was our primary endpoint measure in this study.
Working memory and control task
After the motor skill task, participants were required to perform a secondary task for working memory engagement or a control task (based on the group type described in the next section). The working memory task used in our study was a visuo-spatial Corsi-block tapping task (Fig. 1c). In this task, a blue circle (10 mm diameter) first appeared centrally on the computer screen. Upon entering the circle (with the hand-held stylus), participants heard an audio beep after 750 ms, followed by display of nine white squares. Some of these squares (3–5 out of 9) were flashed (for 750 ms each) in red color in a sequential manner. The order in which the squares changed color from white to red was random. Participants were instructed to carefully watch the flashing squares and remember the order and spatial ___location of the squares that changed the color. Following a second audio beep, participants were required to move their hand (displayed on screen as a yellow dot) to tap the squares that flashed red in the correct sequential order. The time to finish tapping was k + 1 s, where k was the number of squares that flashed red on each given trial (in our case 3–5 squares out of 9). After the participants finished tapping all the squares, feedback was provided on the screen as ‘Trial Successful’ if they tapped the squares in the correct sequence, otherwise ‘Trial Unsuccessful’. Subjects performed a total of 80 trials. The Corsi-block tapping task was performed with the same right arm that was used to train on the motor skill task. For subjects who performed the control task we modified this Corsi-block task such that there was no working memory requirement, and the rest of the task procedure was similar. The squares (3–5) that turned from white to red remained red and subjects were simply asked to tap all the red squares (visible on screen throughout the trial) in any order without touching the white squares. In both the control and working memory tasks, all subjects included in the analysis achieved an accuracy of 70% and above.
Experimental design
As mentioned in the Introduction, we conducted three experiments (with two groups of subjects in each) as part of this study. All three experiments (Exp-1, Exp-2, Exp-3) consisted of an initial training on the motor skill task with dominant right arm, followed immediately by performing a secondary task (working memory/control task) with the same trained right arm. Finally, the untrained left arm was tested for interlimb skill generalization (see Fig. 1d). In Exp-1 (Long-Immediate), subjects received long training on the motor skill task (160 trials) on Day-1, then performed the secondary task (working memory, N = 16 versus control group, N = 16), and were immediately tested for interlimb generalization on the motor skill task (160 trials). For Exp-2 (Long-Delayed), subjects trained on the skill task (160 trials) on Day-1, then performed a secondary task (working memory, N = 15 versus control group, N = 15), and were tested 24 h later for skill generalization (160 trials). Finally based on the results of Exp-1 and Exp-2, in Exp-3 (Short-Delayed), subjects received a short training on the skill task (reduced to 50 trials to prevent asymptotic phase of learning) on Day-1, then performed the secondary task (working memory, N = 15 versus control group, N = 15), and were tested for interlimb skill generalization (160 trials) after 24 h on Day-2.

(a) Setup with TV screen (top), reflective screen (middle) and digitizing tablet (bottom) on which individuals made planar movements and performed all tasks. This was made in Adobe Illustrator 2023 (Order number AD02368900213EIN). (b) Motor skill task with a representative movement from start (green) circle to one of the five target (blue) circles at a distance of 150 mm, connected by yellow lines. Note that only one target appears on each trial. Distance between the red square (indicating hand position at 650 ms of the executed movement) and target circle is considered temporal error. Red line indicating hand movement outside the yellow path is considered spatial error. A composite sum of these two errors is indicated as Motor Error. Participants are instructed to reduce this Motor Error during training. (c) Working memory task indicating start (presentation of nine white squares with blue start circle and yellow dot indicating hand position), cue (in this representation four out of nine squares changed from white to red. Note that the color change was sequential i.e., white squares flashed in red for 750 ms which was to be recalled later testing the working memory. However, for the control task the red squares were present on the screen for the entire trial duration), and the response screen (during which participants moved to touch the squares that turned red). (d) Experimental design depicting Exp-1 (Long-Immediate) with long-training, secondary task (working memory, N = 16; control, N = 16) with right hand, and testing untrained left arm for immediate interlimb generalization. Same for Exp-2 (Long-Delayed, N = 30) except that generalization was delayed by 24 h (working memory, N = 15; control, N = 15). Finally, Exp-3 (Short-Delayed, N = 30) contained short-training, secondary tasks and generalization at 24 h (working memory, N = 15; control, N = 15).
Data analysis and statistical tests
Hand position data (sampled at 120 Hz) were used to assess the motor error on our skill task. Data were analyzed using Matlab R2022b and R studio. Our primary measure of interest in this study was motor skill error which was computed as a sum of spatial and temporal errors (described above and similar to our previous work of Yadav and Mutha25,39). The composite error value (in mm) was used for assessing learning and generalization. We averaged the motor error values for five consecutive trials (based on the five directional targets presented during the task) to reduce any directional bias and obtained bin values (32 bins in Exp-1 and Exp-2 and 10 bins in Exp-3). For each bin, we also computed peak speed (cm/s) and movement preparation time (ms) taken to initiate the movement after the go audio cue (movement start time minus go audio cue time). Finally, we performed mixed analysis of variance (ANOVA) to statistically assess learning, and interlimb generalization across the three experiments with significance level set at 0.05. Pairwise t-tests were conducted when significant main effects were observed in ANOVA and corrected with Bonferroni-holm method for multiple comparisons. We have provided omega-square for effect sizes.
Learning and generalization
Reduction in the skill error, as computed above, over bins indicate successful skill learning during the training session. We performed two-way repeated-measures ANOVA with bin (first, last training) as within-subject and group (working memory, control) as between-subject factors to assess learning on the skill task in all three experiments. Next, for skill generalization from right to the left arm, the first trial of the generalization session was excluded based on the findings of Wang and Sainburg15, who suggested that the nervous system uses the initial trial to assess relevance of prior learning in a new context. Finally, we performed ANOVA with bin (first training, first generalization) as within subject-factor and group as between-subject factor (working memory, control) in our three experiments separately.
Strategies used for learning and generalization
Based on our past work on skill generalization25, we were interested in understanding whether participants developed any strategy during learning and if the same strategy was transferred to the other arm when tested for generalization after engaging in the secondary task in this current study. During skill learning, reduction of motor errors on our task required participants to make fast and accurate movements. We found that participants achieved this goal by making fast movements (i.e., increasing peak speed), or preparing to move as soon as they received the go cue (decreasing movement preparation time) or both. To quantify this, we computed peak movement speed, and movement preparation time for the first and last bins of the training session and assessed if there was a significant change in these parameters. Next, we performed statistical tests to analyze if the strategy adopted during training was also transferred to the untrained arm by comparing the last bin of training and first bin of generalization session in each of the three experiments.
Results
Engaging in working memory task after long-training on a novel motor skill does not prevent immediate or delayed generalization
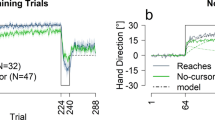
In Exp-1 (Long-Immediate), participants learned to reduce the motor errors on our skill task over the course of training. Hand trajectories of one representative subject are shown in Fig. 2a, where the hand position at 650 ms (denoted by square) in the first bin (first five trials) is close to the start position instead of the target circle (blue circle), and movements are curved outside the specified path. However, on the last bin of the training session, hand positions are close to the target circle, and movements are within the specified path. We found that participants in both groups (working memory, control) improved their performance (reduced motor errors) over the course of training (Fig. 2b). Statistical examination revealed a significant effect of bin (first training, last training) [F[1, 30] = 305.51, p < 0.001, ω2 = 0.20] and no group [F[1, 30] = 1.93, p = 0.17, ω2 = 0.0009] or bin X group interactions [F[1, 30] = 0.01, p = 0.90, ω2 = − 0.001] suggesting similar levels of learning in both groups. Interestingly, participants in both groups exhibited interlimb generalization of learning to the untrained arm (lower motor errors) when tested immediately after engaging in the secondary task. Figure 2a shows that hand paths of the left untrained arm (first bin of generalization) are within the specified path and close to the target circle at 650 ms. We noted a significant effect of bin (first bin training, first bin generalization) [F[1, 30] = 26.47, p < 0.001, ω2 = 0.02], but no group [F[1, 30] = 0.03, p = 0.85, ω2 = − 0.001] or bin X group interactions [F[1, 30] = 0.94, p = 0.33, ω2 = − 0.00004] suggesting similar levels of interlimb generalization following engagement in any secondary task (Working memory or Control task, see Fig. 2c).
Similarly in Exp-2 (Long-Delayed), participants successfully learned the skill over the course of training session (see hand paths in Fig. 3a, and group data in Fig. 3b) with a significant effect of bin (first training, last training) [F[1, 28] = 323.73, p < 0.001, ω2 = 0.21] but no group [F[1, 28] = 0.097, p = 0.75, ω2 = − 0.001] or bin X group interaction [F[1, 28] = 0.00002, p = 0.99, ω2 = − 0.001]. Here too, interlimb generalization was evident when participants were tested 24 h later- significant effect of bin [first bin training, first bin generalization] (F[1, 28] = 82.98, p < 0.001, ω2 = 0.06), with no group [F[1, 28] = 0.24, p = 0.62, ω2 = − 0.001] or bin X group [F[1, 28] = 0.13, p = 0.71, ω2 = − 0.001] interactions. Thus, engaging in a visuo-spatial working memory or control task after learning a novel skill led to similar levels of interlimb generalization to the untrained arm even at 24 h post training (Fig. 3c).
Taken together, findings of Exp-1 and Exp-2 suggest that performing a secondary task (working memory or control task) after learning a novel skill task led to similar amounts of interlimb generalization when the untrained arm is tested immediately or 24 h later. One plausible reason for this generalization could be the long training session with the right arm during which all participants reached the asymptotic performance around the 10th bin of the training session and repeated the learned skilled movement for the rest of the session (22 bins). We conjecture that such repetitive skill behavior may have reduced the requirement for working memory. Longer training may instead engage reinforcement mechanisms where successful actions are selected and executed without much cognitive resources making the memory hyper-stable40,41. To directly test this idea, we performed a third experiment (Short-Delayed) as described earlier in which we shortened the training session to 10 bins. Here again, we tested two groups (Working memory, Control) for interlimb skill generalization 24 h after training.

In Exp-1 (Long-Immediate), engaging in working memory or control task after long skill training does not prevent immediate interlimb generalization. (a) Movement trajectories to the five targets (blue circle) of first and last bin of skill training and generalization sessions, respectively are shown for one participant from each group (working memory in purple, control in green). Note that as compared to the first bin of training, position of square (hand position at 650 ms) is close to the target on all other bins, indicating improved performance by the end of training and subsequent generalization of that performance to the untrained arm in participants from both groups. (b) Change in mean motor error during the training and generalization sessions for the working memory (N = 16) and control group (N = 16). Shaded regions represent standard error (SE). (c) Bar plots comparing motor errors on first, last bins of training and first bin of generalization for the control (left side) and working memory (right side) groups are shown. Lines represent individual participants. Error bars represent SE.

In Exp-2 (Long-Delayed), engaging in working memory or control task after long skill training does not prevent delayed interlimb generalization. (a) Movement trajectories to the five targets (blue circle) of first and last bin of skill training and generalization sessions, respectively are shown for one participant from each group (working memory in purple, control in green). Note, like Exp-1 here too as compared to the first bin of training, position of square (hand position at 650 ms) is close to the target on all other bins, indicating learning and subsequent generalization of that performance to the untrained arm in participants of both groups. (b) Change in mean motor error during training and generalization sessions for the working memory (N = 15) and control group (N = 15). Shaded regions represent SE. (c) Bar plots comparing motor errors on first, last bins of training and first bin of generalization for the control (left side) and working memory (right side) groups are shown. Lines represent individual participants. Error bars represent SE.
Engaging in working memory after short training on skill task impairs delayed interlimb generalization
All participants showed improvements in skill motor performance (reduced motor errors) even with the short training of 50 trials/10 bins. This reduction in the motor errors was found to be significant from first bin to last bin of the training session [F[1, 28] = 147.03, p < 0.001, ω2 = 0.14] and similar for the two groups [F[1, 28] = 0.08, p = 0.76, ω2 = 0.001] with no bin X group interaction [F[1, 28] = 0.13, p = 0.71, ω2 = − 0.001]. However, unlike the previous two experiments, the working memory group exhibited impairment in interlimb generalization as compared to the Control group when tested 24 h later (see hand paths in Fig. 4a). In fact, participants in the working memory group made higher errors throughout the generalization session compared to the control group (see Fig. 4b). Statistically, here we found a significant bin (first training, first generalization) X group (working memory, control) interaction (F[1, 28] = 5.10, p = 0.031, ω2 = 0.003). Post-hoc comparisons revealed that motor errors on the first bin of the generalization session were significantly higher in the working memory group as compared to that of the control group (p = 0.0015), see Fig. 4c.
Next, we wanted to understand how engaging in the working memory task affected delayed interlimb generalization at 24 h depending on the training duration. For this we pooled and directly compared the working memory groups from this experiment (Exp-3 Short-Delayed) and Exp-2 (Long-Delayed). We found that the amount of learning (with a shorter versus longer training) was not different [F[1, 28] = 0.21, p = 0.64, ω2 = − 0.001] in the two groups (Working memory Short-Delayed, Long-Delayed). Next, for interlimb generalization, we found a significant group X bin interaction [F[1, 28] = 4.67, p = 0.039, ω2 = 0.003]. Post-hoc analyses revealed that the working memory group who had received shorter training exhibited significantly higher motor errors on the first bin of generalization as compared to working memory group who received longer training on day-1 (p = 0.008). This suggests that engaging in a secondary working memory task after short, but not long skill training impairs interlimb generalization when tested 24 h later. Also, as evident from Fig. 4b, the working memory group showed higher motor errors throughout the generalization session on Day-2. Thus, to obtain an understanding of the overall difference in the performance of the two groups on Day-2 we calculated the mean motor error of the all bins of the Generalization session and performed a two-sample t-test which revealed a significant difference in the total motor errors of the two groups (t[28] = 2.27, p = 0.03, Cohen’s d = 0.83). This suggests a relatively strong and long-lasting impairment to skill generalization at 24 h that resulted by engaging in working memory following a short training on Day-1.

In Exp-3 (Short-Delayed), engaging in working memory but not control task after short skill training impairs interlimb generalization. (a) Movement trajectories to the five targets (blue circle) of first and last bin of training and generalization sessions, respectively are shown for one participant from each group (working memory in purple, control in green). Note that as compared to the first bin of training, square (hand position at 650 ms) is close to the target on all other bins, indicating improved performance during training and subsequent generalization of that performance to the untrained arm in Control, but not the participant who performed working memory task following skill training. (b) Change in mean motor error during the training and generalization sessions for the working memory (N = 15) and control group (N = 15). Shaded regions represent SE. (c) Bar plots comparing motor errors on first bin, last bin of training and first bin generalization for the control (left side) and working memory (right side) groups are shown. Lines represent individual participants. Error bars represent SE.
Strategies adopted during skill learning and interlimb generalization
We analyzed movement speed and preparation time data to understand strategies adopted by participants to learn the motor task and whether the same strategy was transferred to the untrained arm or interfered with by the secondary task. In Exp-1 (Long-Immediate, see Fig. 5a and b), in majority of the participants we found significant improvements over first and last bins of the training session for both parameters - increase in speed [working memory (13 out of 16): t[12] = − 4.3647, padj = 0.002; control (16 out of 16): t[15] = − 6.3765, padj < 0.001] as well as reduction in preparation time [working memory (15 out of 16): t[14] = 5.3979, padj < 0.001; control (15 out of 16): t[14] = 4.7639, padj < 0.001]. Participants who showed the significant change in the preparation time during training maintained their strategy when generalization to the untrained arm was tested. Their preparation time on the first bin of the generalization session was similar to the last bin of training (working memory: t[14] = 0.3103, padj = 0.761; control: t[14] = 0.1550, padj = 0.879). However, participants of both groups could not transfer the improved movement speed acquired during training to the generalization session. Speed on the first bin of the transfer was significantly lower than speed on the last bin of training (working memory: t[12] = 2.4867, padj = 0.029; control: t[15] = 2.7602, padj = 0.015). We believe this decrease in speed during generalization observed in both groups could be due to task related fatigue since all three sessions (training, secondary task and generalization) were performed in succession.
In Exp-2 (Long-Delayed, see Fig. 5c and d), most participants also exhibited significant changes in both speed [working memory (14 out of 15): t[13] = − 8.5423, padj < 0.001; control (14 out of 15): t[13] = − 5.6208, padj < 0.001] and preparation time [working memory (14 out of 15): t[13] = 5.0196, padj < 0.001; control (12 out of 15): t[11] = 5.2696, padj < 0.001] during the training session. In contrast to Exp-1, both preparation time and speed strategies were transferred to the untrained arm when tested 24 h later. Participants in both groups showed similar preparation time on the first bin of generalization and last bin of training session (working memory: t[13] = − 0.5600, padj = 0.585; control: t[11] = 0.5418, padj = 0.599). Similarly, speed on the first bin of the generalization session was not different from the last bin of training (working memory: t[13] = 1.9589, padj = 0.072; control: t[13] = 1.4585, padj = 0.168).
Despite reduced training, most participants in Exp-3 (Short-Delayed, see Fig. 5e,f) also showed changes in both speed [working memory (9 out of 15): t[8] = − 5.7564, padj < 0.001; control (11 out of 15): t[10] = − 3.9344, padj = 0.006] as well as preparation time [working memory (14 out of 15): t[13] = 3.4755, padj = 0.008; control (10 out of 15): t[9] = 5.0684, padj = 0.001] during training. Interestingly, participants in the control group maintained the strategy in the generalization session when first bin of transfer was compared to last bin of training- comparable speed (t[10] = 1.032, padj = 0.326) with a further reduction in preparation time (t[9] = 3.6521, padj = 0.005). On the other hand, participants in the working memory group who improved (a decrease) their preparation time during the training could not maintain this acquired strategy from end of training to the first bin of generalization session; with worsening (an increase) in preparation time on first generalization bin as compared to last training bin (t[13] = 2.3455, padj = 0.036, see Fig. 5e). An outlier can be seen in Fig. 5e which may have skewed these results. As a robustness check, we excluded this subject and reanalysed the data and found that the preparation time on the first bin of the generalization session was still significantly different (worse) from last bin of training for the working memory group (t[12] = 3.479, padj = 0.005). Interestingly, however, participants of this group who improved speed (an increase) during training maintained this strategy on the first transfer bin as in the last bin of training (t[8] = 0.6615, padj = 0.527).

Strategy adopted during training and its subsequent generalization. For Exp-1 (Long-Immediate, top plots) (a) mean movement preparation time and (b) mean peak speed for individual participants (control indicated in green, working memory in purple) on first, last bins of training and first bin of generalization are shown. Bars represent group-averaged values. Most participants (see Results section for details) reduced (a) preparation time and increased (b) movement speed over the first and last bin of training in both groups and transferred only improved preparation time strategy to the untrained arm when tested for generalization. In Exp-2 (Long-Delayed, middle plots) most participants decreased (c) preparation time, and (d) increased speed during training with both strategies transferring to the untrained arm (see first gen) in both groups. Finally, in Exp-3 (Short-Delayed, bottom plots) most participants in both groups decreased (e) preparation time and increased (f) speed during training. However, here the control group transferred both strategies to the untrained arm, but the working memory group only transferred improved speed strategy with worsening of preparation time (see first gen). Error bars represent SE.
Overall, we found that both movement speed and preparation time underwent a significant change during training in all groups across our three experiments which helped participants learn the novel skill task. In the context of interlimb generalization, we found that the strategies that were transferred to the untrained arm depended on the experiment and group type. In Exp-1 with long training and immediate generalization test, participants in both groups transferred the movement preparation time strategy only (but not speed, possibly due to task fatigue) to the untrained arm. For Exp-2 with long training and delayed generalization test, participants in both groups transferred movement preparation time as well as speed strategies to the untrained arm. Interestingly, in Exp-3 with short training and delayed generalization test, the control group transferred both strategies of increased speed, with a further improved preparation time, whereas the working memory group transferred only speed, with worsening of preparation time. These results suggest that movement planning linked to preparation time that evolved during short training is dependent on working memory processing. On the other hand, motor control processes such as movement speed are likely not dependent and thus immune to working memory interference.
Discussion
In this study we demonstrate that engaging in a visuo-spatial working memory task as compared to control task immediately after a short skill training interferes with interlimb skill generalization tested at 24 h. On the other hand, such generalization when tested immediately or at 24 h is robust, and therefore not prone to any interference from working memory when the preceding skill training is long. This was surprising as the amount of learning was similar across all groups (despite long or short training) who successfully acquired the skill by adopting strategies such as increased movement speed and decreased preparation time by the end of the training session. While further exploring the specific nature of the interference caused by working memory to interlimb generalization in the short training group, we found that the strategy related to improved movement preparation time is not transferred (and in fact gets worse) to the untrained arm when it is tested at 24 h. This suggests that engaging in a secondary cognitively demanding task engaging working memory immediately after short skill training affects skill memory stabilization in a specific manner such that the strategy related to movement preparation, but not speed, is impaired and thus not transferred to the untrained arm.
Humans have a large capacity to perform many different tasks in quick succession. However, little is known about the consequence of engaging in cognitively intensive tasks in quick succession and its impact on task memory stabilization. For instance, Cohen and Robertson27 show that engaging in different cognitive tasks such as skill acquisition and memorizing a list of words in quick succession can result in an underlying neural competition for memory processing. Working memory driven impairment in interlimb generalization of skill memory observed in the short skill training group is suggestive of a possible common processing bottleneck. Moreover, participants in our study were learning the skill under a variable condition which requires constant processing of the visual cues (as target ___location is random) before planning and executing the movement. Consequently, such learning is associated with higher cognitive load for successful performance on the task41. In other words, variable skill learning requires trial-by-trial update of the visual cue information to aid movement execution. This working memory processing requirement is higher in the early stages of skill learning in variable task condition, such as ours, and likely more dependent on prefrontal brain areas36,42,43.
Some prior work suggests that tasks requiring relational or associative processing elicit DLPFC activity44,45,46. It therefore appears that motor skill task with variable structure and the working-memory Corsi block-tapping task both share this underlying feature of relational or associative processing related to movement planning. Both these tasks, therefore, depend on DLPFC which plays an important role in initially retaining spatial information for a brief duration to allow further processing by cortical and subcortical areas to generate the desired movement47. In our study when participants performed the working memory task after short skill training, the stabilization/consolidation of the newly acquired motor memory was affected which resulted in impaired generalization to the untrained arm when tested the subsequent day. We propose that engaging in the working memory task likely led to an impairment in post-learning motor memory processing related to planning, as movement preparation strategy was not transferred to the untrained arm. This impairment was not observed for participants who performed the control task after short skill training, possibly because of the lack of underlying neural competition in this case. Moreover, both working memory and control groups could transfer the strategy related to speed, which we believe could be the result of motor control processes that rely on other brain areas such as the primary motor cortex, involved in processing movement kinematics in humans48. This suggests that at least one component (linked to movement planning) of the newly formed motor skill memory acquired after short training, reflecting the early stage of skill learning, in our study is critically dependent on working memory processing for its further stabilization and subsequent generalization to the untrained arm.
Interestingly, long skill training resulted in significant immediate (Exp-1) as well as delayed (Exp-2) skill generalization with no interference from the subsequent working memory or control tasks. Moreover, in these experiments with long training, strategies that generalized to the untrained arm (movement preparation time transferred in Exp-1, and both movement time and speed transferred in Exp-2) did not differ for working memory or control group. We believe long training leading to the late stage of skill learning may sufficiently stabilize the newly formed skill memory with lesser reliance on working memory processing and therefore immune to interference from the secondary working memory task. This view of ours is supported by work of Shibata and colleagues40 who suggest that extra training after performance has reached an asymptote can make the newly acquired memory hyper-stable, thus impervious to subsequent interference from other tasks. As previously described in the introduction, skill human behavior is contingent on stable memory representations. Long training, therefore, is a simple way to make skill memory representation more stable and protect it from further interference from secondary follow-up tasks. Furthermore, interlimb generalization observed in all these groups (right to left arm transfer, except for Exp-3 Short-Delayed working memory group) suggests that the memory representation formed during novel skill learning has effector-independent features which can transfer to the untrained non-dominant arm- a phenomenon often not observed in interlimb generalization of motor adaptation14,15,17,18. Moreover, some interesting work in the context of generalization of effector-independent components following motor adaptation49,50 has shown that interlimb transfer is highly context dependent and certain offline processes (such as passage of time and/or moving under same conditions as learning) may facilitate it. On the other hand, such generalization may diminish if the effector independent representations decay when learning is immediately followed by moving in a novel context. Such views are in line with our current study wherein learning skill in short-training condition (which isn’t conducive for offline processes to fully preserve the effector-independent representation) and followed by engaging in a secondary novel task became detrimental for interlimb skill generalization. The specific nature of such newly acquired effector-independent skill representations and how those are modulated by memory stabilization (or decaying) processes that can influence generalization requires further detailed investigations.
Taken together, our findings show that newly learned skill can generalize to the untrained arm when tested immediately or 24 h later following a secondary task performed after long skill training. However, such robust interlimb generalization is absent when individuals receive short skill training followed by engaging in a working memory task. Our work, therefore, highlights an underlying interaction between hitherto two distinct memory systems, i.e., skill and working memory, and the critical role of working memory in early stages of skill learning and its generalization to the untrained effector.
Data availability
The datasets used and/or analysed during the current study available from the corresponding author on reasonable request.
References
Robertson, E. M. From creation to consolidation: a novel framework for memory processing. PLoS Biol. 7, e19 (2009).
Reis, J. et al. Noninvasive cortical stimulation enhances motor skill acquisition over multiple days through an effect on consolidation. Proc. Natl. Acad. Sci. 106, 1590–1595 (2009).
Shmuelof, L., Krakauer, J. W. & Mazzoni, P. How is a motor skill learned? Change and invariance at the levels of task success and trajectory control. J. Neurophysiol. 108, 578–594 (2012).
Telgen, S., Parvin, D. & Diedrichsen, J. Mirror reversal and visual rotation are learned and Consolidated via separate mechanisms: recalibrating or learning De Novo? J. Neurosci. 34, 13768–13779 (2014).
Yadav, G. & Duque, J. Reflecting on what is skill in human motor skill learning. Front. Hum. Neurosci. 17, 1117889 (2023).
Shadmehr, R. & Mussa-Ivaldi, F. Adaptive representation of dynamics during learning of a motor task. J. Neurosci. 14, 3208–3224 (1994).
Thoroughman, K. A. & Shadmehr, R. Learning of action through adaptive combination of motor primitives. Nature 407, 742–747 (2000).
Taylor, J. A., Krakauer, J. W. & Ivry, R. B. Explicit and implicit contributions to learning in a sensorimotor adaptation task. J. Neurosci. 34, 3023–3032 (2014).
Poggio, T. & Bizzi, E. Generalization in vision and motor control. Nature 431, 768–774 (2004).
Krakauer, J. W., Mazzoni, P., Ghazizadeh, A., Ravindran, R. & Shadmehr, R. Generalization of motor learning depends on the history of prior action. PLoS Biol. 4, e316 (2006).
Joiner, W. M., Brayanov, J. B. & Smith, M. A. The training schedule affects the stability, not the magnitude, of the interlimb transfer of learned dynamics. J. Neurophysiol. 110, 984–998 (2013).
Renault, A. G. et al. Individual movement features during prism adaptation correlate with after-effects and interlimb transfer. Psychol. Res. 84, 866–880 (2020).
Criscimagna-Hemminger, S. E., Donchin, O., Gazzaniga, M. S. & Shadmehr, R. Learned dynamics of reaching movements generalize from dominant to nondominant arm. J. Neurophysiol. 89, 168–176 (2003).
Wang, J. & Sainburg, R. L. Mechanisms underlying interlimb transfer of visuomotor rotations. Exp. Brain Res. 149, 520–526 (2003).
Wang, J. & Sainburg, R. L. Interlimb transfer of novel inertial dynamics is asymmetrical. J. Neurophysiol. 92, 349–360 (2004).
Bao, S., Lei, Y. & Wang, J. Experiencing a reaching task passively with one arm while adapting to a visuomotor rotation with the other can lead to substantial transfer of motor learning across the arms. Neurosci. Lett. 638, 109–113 (2017).
Kumar, N., Kumar, A., Sonane, B. & Mutha, P. K. Interference between competing motor memories developed through learning with different limbs. J. Neurophysiol. 120, 1061–1073 (2018).
Kumar, A., Panthi, G., Divakar, R. & Mutha, P. K. Mechanistic determinants of effector-independent motor memory encoding. Proc. Natl. Acad. Sci. 117, 17338–17347 (2020).
Wang, J. & Sainburg, R. L. Interlimb transfer of visuomotor rotations depends on handedness. Exp. Brain Res. 175, 223–230 (2006).
Poh, E., Carroll, T. J. & Taylor, J. A. Effect of coordinate frame compatibility on the transfer of implicit and explicit learning across limbs. J. Neurophysiol. 116, 1239–1249 (2016).
Inui, N. Lateralization of bilateral transfer of visuomotor information in right-handers and left-handers. J. Mot. Behav. 37, 275–284 (2005).
Chase, C. & Seidler, R. Degree of handedness affects intermanual transfer of skill learning. Exp. Brain Res. 190, 317–328 (2008).
Lefumat, H. Z. et al. To transfer or not to transfer? Kinematics and laterality quotient predict interlimb transfer of motor learning. J. Neurophysiol. 114, 2764–2774 (2015).
Thompson, A. K. B. & Henriques, D. Y. P. Visuomotor adaptation and intermanual transfer under different viewing conditions. Exp. Brain Res. 202, 543–552 (2010).
Yadav, G. & Mutha, P. K. Symmetric interlimb transfer of newly acquired skilled movements. J. Neurophysiol. 124, 1364–1376 (2020).
Robertson, E. M., Pascual-Leone, A. & Miall, R. C. Current concepts in procedural consolidation. Nat. Rev. Neurosci. 5, 576–582 (2004).
Cohen, D. A. & Robertson, E. M. Preventing interference between different memory tasks. Nat. Neurosci. 14, 953–955 (2011).
Robertson, E. M. New insights in human memory interference and consolidation. Curr. Biol. 22, R66–R71 (2012).
Seidler, R. D. Multiple motor learning experiences Enhance Motor adaptability. J. Cogn. Neurosci. 16, 65–73 (2004).
Mosha, N. & Robertson, E. M. Unstable memories create a high-level representation that enables learning transfer. Curr. Biol. 26, 100–105 (2016).
Robertson, E. M. Memory instability as a gateway to generalization. PLoS Biol. 16, e2004633 (2018).
Sala, J. B., Rämä, P. & Courtney, S. M. Functional topography of a distributed neural system for spatial and nonspatial information maintenance in working memory. Neuropsychologia 41, 341–356 (2003).
Klauer, K. C. & Zhao, Z. Double dissociations in visual and spatial short-term memory. J. Exp. Psychol. Gen. 133, 355–381 (2004).
Fregni, F. et al. Anodal transcranial direct current stimulation of prefrontal cortex enhances working memory. Exp. Brain Res. 166, 23–30 (2005).
Sun, R., Zhang, X., Slusarz, P. & Mathews, R. The interaction of implicit learning, explicit hypothesis testing learning and implicit-to-explicit knowledge extraction. Neural Netw. 20, 34–47 (2007).
Kantak, S. S., Sullivan, K. J., Fisher, B. E., Knowlton, B. J. & Winstein, C. J. Neural substrates of motor memory consolidation depend on practice structure. Nat. Neurosci. 13, 923–925 (2010).
Oldfield, R. C. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9, 97–113 (1971).
Brainard, D. H. The Psychophysics Toolbox. Spat. Vis. 10, 433–436 (1997).
Yadav, G. & Mutha, P. K. Deep breathing practice facilitates retention of newly learned motor skills. Sci. Rep. 6, 37069 (2016).
Shibata, K. et al. Overlearning hyperstabilizes a skill by rapidly making neurochemical processing inhibitory-dominant. Nat. Neurosci. 20, 470–475 (2017).
Haith, A. M. & Krakauer, J. W. The multiple effects of practice: skill, habit and reduced cognitive load. Curr. Opin. Behav. Sci. 20, 196–201 (2018).
Kumar, N., Sidarta, A., Smith, C. & Ostry, D. J. Ventrolateral prefrontal cortex contributes to human motor learning. Eneuro 9, ENEURO.0269-22.2022 (2022).
Miller, E. K. & Cohen, J. D. An integrative theory of prefrontal cortex function. Annu. Rev. Neurosci. 24, 167–202 (2001).
Smith, E. E. & Jonides, J. Working memory: a View from Neuroimaging. Cogn. Psychol. 33, 5–42 (1997).
Nelson, C. A. et al. Functional neuroanatomy of spatial working memory in children. Dev. Psychol. 36, 109–116 (2000).
Robertson, E. M., Tormos, J. M., Maeda, F. & Pascual-Leone, A. The role of the dorsolateral prefrontal cortex during sequence learning is specific for spatial information. Cereb. Cortex. 11, 628–635 (2001).
Pochon, J. B. et al. The role of dorsolateral prefrontal cortex in the preparation of forthcoming actions: an fMRI study. Cereb. Cortex. 11, 260–266 (2001).
Truccolo, W., Friehs, G. M., Donoghue, J. P. & Hochberg, L. R. Primary motor cortex tuning to intended movement kinematics in humans with tetraplegia. J. Neurosci. 28, 1163–1178 (2008).
Kitago, T., Ryan, S. L., Mazzoni, P., Krakauer, J. W. & Haith, A. M. Unlearning versus savings in visuomotor adaptation: comparing effects of washout, passage of time, and removal of errors on motor memory. Front. Hum. Neurosci. 7, 307 (2013).
Bao, S., Wright, D., Buchanan, J. & Lei, Y. The decay and consolidation of effector-independent motor memories. https://doi.org/10.21203/rs.3.rs-154034/v1 (2021).
Acknowledgements
We would like to thank Bhoomika Sonane and Rechu Diwakar for their help with task setup.
Funding
This work was supported by DST CSRI (DST/CSRI/2021/164(C)(G)) and SERB SRG (SRG/2021/001125) grants to NK, and postdoctoral research grant by FNRS (FC 41003) to GY.
Author information
Authors and Affiliations
Contributions
G.Y., R.P. and N.K. designed the study and wrote the paper. R.P. and S.M. collected and analysed the data.All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Yadav, G., Pal, R., Matkar, S. et al. Role of working memory in interlimb generalization of newly learned skills. Sci Rep 15, 10026 (2025). https://doi.org/10.1038/s41598-025-88997-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-025-88997-z
