
Abstract
Fritillaria taipaiensis P. Y. Li has received much research attention because of its high medicinal value and edibility. Analysis of the areas of suitable habitat for F. taipaiensis can aid the conservation of its germplasm resources. We used the Maxent model to predict areas with suitable habitat under current and future climate scenarios; we also determined the primary climatic factors affecting the distribution of F. taipaiensis. Currently, suitable habitats for F. taipaiensis are concentrated in the western and central regions of China. The area of suitable habitat in this region will decrease in the future; some patches of suitable habitat will decrease in size and migrate to higher latitudes. Pivotal environmental factors that affect the distribution of F. taipaiensis is temperature. Northeastern Chongqing, southern Shaanxi, western Hubei, and northwestern Yunnan were the suitable areas for F. taipaiensis, which was basically consistent with the predicted areas in the Maxent model. The findings in the study are important for the conservation and use of F. taipaiensis specifically and Fritillaria plants more generally.
Similar content being viewed by others


Introduction
Fritillaria taipaiensis P. Y. Li are fritillaries in the family Liliaceae, one of the six basal plants of Sichuan Beimu, and they are mainly distributed in the Qinling Mountains in Shaanxi, Chongqing, Sichuan, and other provinces. F. taipaiensis can be used to moisten the lungs, relieve cough, as well as eliminate phlegm and wheezing; it is thus widely used to treat lung heat, dry cough, and lung-Yin deficiency. F. taipaiensis, a medicinal species with centuries of traditional use1. An increasing number of studies have examined the medicinal value and health-promoting properties of F. taipaiensis, and this has contributed to increases in the demand for F. taipaiensis. The scale of artificial cultivation is also expanding, and this has been accompanied by the excessive consumption and even exhaustion of wild F. taipaiensis resources2. The germplasm resources, ecological environment, and other variables correlated with the distribution of F. taipaiensis have been studied through a review of previous studies and field research in the production areas. However, few studies have attempted to predict the distribution of F. taipaiensis under current conditions and future climatic scenarios. Zou Meng et al.3. predicted the distribution of suitable habitat for F. taipaiensis in the core area of its range based on current climatic conditions, but the prediction accuracy under future climatic conditions was poor; furthermore, their discussion of future spatial changes was incomplete. The White Paper on Biodiversity Conservation in China indicated that the conservation of biodiversity should become an important national strategy and emphasized the need to confirm priority areas for conservation to effectively protect key ecosystems, species, as well as their habitats4. Therefore, future studies are needed to predict the potential distribution of habitats for F. taipaiensis and identify the future suitable habitat area for F. taipaiensis, the primary environmental factors influencing its geographic distribution, and ecological niche differences.
Climate change can affect plant growth and development and population sizes5, especially geographical distributions. Large changes in global climate have occurred in recent centuries due to human and natural factors6. Global warming has been accompanied by gradual increases in the carbon dioxide concentration, and this has induced shifts in the distributions of plants to higher latitudes. Climate change affects plants in various ways7; thus, studies regarding the responses of plants to climate change are critically important in biodiversity conservation. Species diversity constitutes the basis of biodiversity, and conventional sampling site-based field surveys of biodiversity measures have various limitations8. Data derived from comparisons of biodiversity trends across multiple taxa are limited, and direct comparisons between studies are hampered by significant differences in their temporal and spatial coverage and data resolution9. The conventional species-based approaches are time- and labor-consuming, and they do not provide spatial-temporal biodiversity information; they are thus not applicable to large-scale biodiversity assessments and conservation programs10. GIS-supported species distribution information and corresponding spatial environmental covariates can be used to determine species ranges, identify suitable habitats, set conservation priorities, and predict range changes under future climate change scenarios11. Previous studies have also been conducted to create multivariate statistical models and perform spatial or temporal modelling to aid inferences of the area of potentially suitable habitats12.
Maximum Entropy (Maxent) models are based on the ecological niche principle. They utilize species distribution ___location information and environmental variable data to generate a probability distribution of maximum entropy values to predict the potential distribution of species13. Hence, information regarding the species’ potential geographic distribution locations can be rapidly and accurately obtained14. Relative to other species distribution prediction models (the Ecological Niche Factor Model (ENFA), the Bioclimatic Analysis System (BIOCLIM), and the Genetic Algorithm for Rule-set Production (GARP)), the Maxent model shows higher modelling accuracy15,16, which allows better predictions to be obtained, even in the absence of occurrence records with accurate coordinates17. This model is also robust to small sample sizes, and the statistical significance of the output can be tested. This model can also handle environmental variables for continuous and categorical data, and the output comprises easily interpretable continuous probabilities18,19,20. It can be used in assessments of the environmental variables affecting species distributions and the prediction of species’ geographic distributions under global climate change12; such analyses have been performed on rare and endangered plants21, beneficial plants22, invasive plants23, and quarantine pests and diseases24. For example, Chinese caterpillar fungus25, Codonopsis pilosula26. However, there are very few studies on F. taipaiensis.
Ecological niche theory, namely ecological niche overlap theory (ENOT) and width theory (ENWT), can reveal interactions among species, organisms, and the environment. ENOT can quantify the overlap in the use of and competition for resources by species in various environments within biomes; ENWT explains the dominant resource range within biomes27,28. ENMTools can be used to analyse changes in species ecotopes and calculate ecotope widths, as well as to quantify ecological niche overlap and width index. Analyses of differences in species ecotope overlap and ecotope width with time and climate change can help determine species ecotope characteristics and verify the model’s prediction accuracy for the potential distribution of species.
This study aims to (1) predict the potential distribution patterns of F. taipaiensis and analyze its spatiotemporal dynamics using the Maxent model; (2) identify key environmental determinants influencing the species’ distribution; and (3) investigate ecological niche differentiation under climate change scenarios. These findings provide critical insights for reducing unsustainable wild harvesting of F. taipaiensis. Through integration of multiple Shared Socioeconomic Pathways (SSPs) and comprehensive species occurrence data, we conducted a holistic analysis of the species’ ecological adaptability. The results establish a scientific foundation for developing effective conservation strategies and optimizing utilization of germplasm resources.
Materials and methods
Data source and processing
Data source and processing of F. taipaiensis occurrence records
We initially collected thirty-eight occurrence records of F. taipaiensis through systematic field surveys conducted across Chongqing, Shaanxi, Sichuan, and Yunnan provinces in China. Supplementary occurrence data were acquired from two authoritative repositories: the Chinese Virtual Herbarium (CVH; http://www.cvh.ac.cn) and the Global Biodiversity Information Facility (GBIF; https://www.gbif.org), with temporal filtering restricted to post-1980 observations. The combined dataset comprised 53 georeferenced occurrences. To address potential geospatial redundancies and positional inaccuracies, we implemented a spatial filtering protocol using the ENMTools package in R software. This rigorous validation process yielded 23 spatially unique occurrence records suitable for subsequent analyses (Table 1).
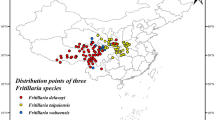
We saved the F. taipaiensis occurrence records as a .csv file in Microsoft Excel, which contained data on species name, longitude, and latitude for Maxent modelling (Fig. 1).

Distribution position of sampling points of F. taipaiensis, the map creation using ArcMap v10.2 (URL: https://www.arcgis.com/index.html).
We screened 37 environmental factors, including 19 bioclimatic factors (mean annual temperature; mean diurnal temperature difference (MDTD), isotherm, temperature seasonality, max temperature in the hottest month, min temperature in the coldest month, annual change in temperature, mean temperature in the wettest season, mean temperature in the driest season, mean temperature in the hottest season, and mean temperature in the coldest season, mean annual precipitation, precipitation in the wettest month, precipitation in the driest month, and precipitation (Variance of Change, Wettest Quarterly Precipitation, Driest Quarterly Precipitation, Hottest Quarterly Mean Precipitation, and Coldest Quarterly Mean Precipitation); 2 topographic factors (elevation and slope); and 16 soil factors (basic saturation, carbonate or lime content, sulfate content, cation exchange capacity, clayey soils, clay content of soil cation exchange capacity, percentage by volume of crushed rock, exchangeable sodic salts, electrical conductivity, organic carbon content, pH, soil capacity, sand content, chalk content, exchangeable substrate classification, and USDA soil texture).
Current (1970–2000) and future (2040s: 2041–2060, 2060s: 2061–2080) climate data were obtained from the World Climate Database Worldclim2.1 (http://www.worldclim.org/), with each period covering the 19 bioclimatic factors: bio1–bio19. The future data are selected from the prediction data of the BCC-CSM2-MR model in the Sixth International Coupled Model Intercomparison Project (CMIP6)29, This model is applicable to China ‘s geographical environment, which is a combined scenario of Shared Socio-Economic Pathways (SSPs) and RCPs, instead of a Typical Concentrated Pathway (RCP) scenario in CMIP5. It has implications for future socio-economic development30.
The 3 combined scenarios, SSP1-2.6, SSP2-4.5, and SSP5-8.5, denote the low, medium, and high radiative forcing scenarios, respectively. The data have a spatial resolution of 2.5 arc-minutes (~ 5 km). Soil factors were derived from the World Soil Database (HWSD)-based soil dataset from the Cold and Arid Regions Science Data Centre (CARSDC), and topographic data were derived from the Geospatial Data Cloud (http://www.gscloud.cn/). Suitable areas were predicted, assuming that the soil and topographic factors would remain unchanged over the following 70 years31.
We systematically evaluated 37 environmental variables through a multi-step screening protocol. Initial variable selection employed Spearman’s rank correlation analysis (ρ > 0.8 threshold) combined with jackknife test evaluation of variable importance (Figure S1), implemented using R software. Following this statistical filtering, we conducted iterative manual screening to: (1) Eliminate variables demonstrating < 1% contribution to model performance based on permutation importance metrics; (2) Address multicollinearity by retaining only the most ecologically interpretable variable from highly correlated pairs (|ρ| ≥ 0.8)32.
According to the above method, 10 of the 37 environmental factors were identified as relatively important factors according to their influence on the geographical distribution of F. taipaiensis (Table 2).
Maxent model construction and optimization
We used 75% of the samples in the Maxent model as the training dataset, and 25% of the samples as the test dataset for model validation; we repeated the modelling procedure 10 times after setting the max number of iterations to 10,000. We selected bootstrap as the repetition type, and the distribution values were output in the logistic format. The model accuracy assessment relied on the area under the curve (AUC) value in the range of 0.5 to 1.0. An AUC value approaching 1 indicated an increase in the prediction accuracy; 0.5–0.7, 0.8–0.9, and 0.9–1.0 indicated poor, good, and very accurate prediction, respectively33.
Adjustments were performed on the feature combination (FC) and the regularization multiplier (RM) to optimize the model via the ENMeval package in R software. The Maxent model possesses 5 feature combinations: linear, quadratic, product, threshold, and segmented features34. In the Maxent software, the default parameters were RM = 1 and FC = LQHPT; for achieving Maxent model optimization, RM was in the range of 0.5 to 4 and increased by 0.5 each time. Eight adjustment multipliers were performed, and 6 combinations of ≥ 1 features were adopted, including L, L and Q, H, L, Q, H and H, L, Q, H and P, L, Q, H, P and T. These permutations and combinations were taken into account to obtain 48 parameter combinations. After testing the 48 combinations, we examined the model complexity using the delta values. Values of AICc and (auc.train auc.diff.avg) could also be used to evaluate model accuracy, and lower values denoted higher prediction accuracy.
Changes in the area of suitable habitat for F. taipaiensis
Firstly, ArcGIS natural breakpoint method was used to divide the suitable area of F. taipaiensis into four grades : highly suitable area (P ≥ 0.425), moderately suitable area (0.201 ≤ P < 0.425), lowly suitable area (0.056 ≤ P < 0.201) and unsuitable area (P < 0.056). Then, the reclassification tool in ArcGIS 10.8 software was applied to the data binarization of the potential geographic distribution of F. taipaiensis under two scenarios, and a relevant presence/absence (0,1) matrix was created. Suitable areas (species presence) were represented by a value of “1,” and “0” represented unsuitable areas (species absence). Based on this matrix, the spatial patterns of potentially suitable areas for F. taipaiensis were further analysed; the areas were then classified into three different categories: newly suitable areas (matrix values 0→1), retreating suitable areas (1→0), and stable suitable areas (1→1)35,36.
Niche analysis
We quantitatively analysed the ecological niches of F. taipaiensis in different periods and under different climate scenarios based on environmental factor layers and distribution layers; we also visualized the ecological niche through the “ecospat” package in R software37. We then calculated the ecological niche overlap index (overlap percentage index), denoted by the letter D. Values of this index ranged from 0 to 1, which corresponded to ecological niches with no overlap to complete overlap38. The ecotope width module in ENMTools was used to calculate the ecotope width while taking into account the potential distribution data for F. taipaiensis under current and future climates, and B1 and B2 denote the min and max ecotope width, respectively.
Data processing
ArcGIS V10.8 software was used to transform prediction results into raster data. The raster values indicated the probability of survival (POS) of F. taipaiensis in the area. The centre-of-mass (COM) positions of the suitable areas for F. taipaiensis were calculated using the SDMTools data package in R. Changes in the COM positions reflected the migration direction of the suitable areas. The COM offset distances were calculated using the geosphere data package in R.
Results
Optimal model and accuracy evaluation
Combining the 23 F. taipaiensis distribution points and the 10 environmental factors, a simulation was conducted on the species’ potential distribution areas using the Maxent model. With the model set to the default parameter, DAICc = 23.57 (Table 3); the model was optimized (DAICc = 0) when FC = LQ and RM = 1.5 (Fig. 2). Hence, FC = LQ and RM = 1.5 were used as the modelling parameters in subsequent analyses. We used the optimized parameters for model reconstruction and simulation of F. taipaiensis. The simulation training AUC value was 0.993 (Figure S2), which indicated that the predictions were accurate.
Figure 3 presents the variable importance metrics and regularized training gain values derived from the Maxent model. The regularized training gain algorithm accounts for variable interdependencies, enabling standardized comparison of predictor contributions. Among the bioclimatic variables, three temperature-related parameters demonstrated dominant influence: minimum temperature of coldest month (BIO6, permutation importance: 1.501), mean temperature of driest quarter (BIO9, 1.402), and temperature diurnal range (BIO2, 0.859).

Optimisation results of Maxent model under different parameter settings.

Jackknife text of the importance of environment variables in Maxent.
Potentially suitable habitat areas for F. taipaiensis under current conditions
ArcGIS software was used to generate a map of the suitable areas for F. taipaiensis under current climate conditions (Fig. 4). Suitable habitat for F. taipaiensis was concentrated in central and western China, including Sichuan, Yunnan, Gansu, Shaanxi, Henan, Hubei, Guizhou, Xizang, Chongqing, and Shandong Provinces. Highly suitable areas were concentrated in northeastern Chongqing, southern Shaanxi, western Hubei, and northwestern Yunnan, and moderately suitable areas were concentrated in southeastern Longdong, northwestern Yunnan, northeastern Chongqing, and southern Shaanxi. The simulation results and prediction results were consistent with the geographical distribution data of F. taipaiensis, indicating that the accuracy of the results was high.

Potential Distribution Area of F. taipaiensis in China under current climate conditions, the map creation using ArcMap v10.2(URL: https://www.arcgis.com/index.html).
The suitable area was determined based on the number of grids occupied by suitable regions (Table 4). The suitable area of F. taipaiensis covered 425,273,000 km2 under current climate conditions, which is approximately 4.44% of the total land area of China; the highly suitable area was small, covering 46,839,000 km2.
Prediction of the suitable area for F. taipaiensis under future climate scenarios
The Maxent model outputs were converted into geospatial raster layers (30 arc-second resolution) to quantify habitat suitability for F. taipaiensis. Projections under future climate scenarios revealed divergent spatiotemporal patterns of suitable area dynamics across emission pathways (Table 4). Notably: Under the SSP1-2.6 scenario (low emissions), suitable areas declined by 12.3% (2041–2060) before rebounding to 97.4% of baseline levels by 2080, suggesting mitigated habitat loss under constrained warming. The SSP2-4.5 trajectory (intermediate emissions) exhibited progressive expansion (+ 28.26% cumulative gain by 2100), indicating potential range shifts facilitated by moderate socioeconomic development. SSP5-8.5 projections (high emissions) showed sustained contraction (– 1.79 to – 9.34 km2/yr), highlighting complex bioclimatic constraints under rapid warming and intensive resource utilization.
Habitat suitability projections for F. taipaiensis were spatially visualized using ArcGIS 10.8. Comparative analysis of decadal-scale distribution patterns revealed distinct climate response trajectories across emission scenarios (Fig. 5). The SSP1-2.6 pathway exhibited non-linear dynamics with moderate habitat contraction (– 1.79 km2, 2041–2060) followed by partial recovery (+ 3.54 km2, 2061–2080), suggesting transitional adaptation during early warming phases. Contrastingly: SSP2-4.5 demonstrated progressive expansion (+ 2.92 km2 cumulative gain). SSP5-8.5 showed accelerated habitat fragmentation (– 2.60 km2 net loss). Notably, temporal variability magnitude differed substantially between scenarios, with SSP1-2.6 displaying greater inter-decadal fluctuations compared to more stable trends in SSP2-4.5 and SSP5-8.5.
These projections indicate that climate warming will generally exert negative pressure on F. taipaiensis habitats, particularly under high-emission scenarios. However, the SSP1-2.6 pathway suggests two critical conservation insights: Limited near-term habitat reduction, and emergent adaptive capacity evidenced by late-century recovery trends. The observed decadal-scale habitat plasticity under moderate warming (SSP1-2.6) implies potential physiological tolerance thresholds that could inform assisted migration strategies. These findings underscore the species’ vulnerability to unabated emissions while identifying critical windows for climate-adaptive conservation planning.

Fertile distribution areas of F. taipaiensis under SSP126 (A,D), SSP245 (B,E), SSP585 (C,F) scenarios in 2040 (A,B,C) and 2060 (D,E,F), the map creation using ArcMap v10.2(URL: https://www.arcgis.com/index.html).
Centre-of-mass positions shifts of F. taipaiensis under future climate scenarios
Spatiotemporal analysis revealed distinct geographic shifts in F. taipaiensis habitat centroids across climate scenarios, demonstrating systematic northwestward migration patterns (Fig. 6). At present, the COM suitable for F. taipaiensis is located in Jingyang District, Deyang City, Sichuan Province (31.053 °N, 104.481 °E). In the 2060s-SSP1-2.6 scenario, the COM in the suitable area of F. taipaiensis moved northwestward. The COM in the suitable area of F. taipaiensis was located in Xiaojin County (30.966 °N, 102.832 °E), Aba Tibetan and Qiang Autonomous Prefecture, Sichuan Province, with a migration distance of 157,660 m. Under the 2060s-SSP2-4.5 scenario, the COM in the suitable area of F. taipaiensis is located in Maerkang City (32.238 °N, 102.262 °E), Aba Tibetan and Qiang Autonomous Prefecture, Sichuan Province, with a migration distance of 248 m and 210 m. In the 2040 s-SSP1-2.6, SSP2-4.5 and SSP5-8.5 scenarios, COM also moved northwestward, located in Beichuan Qiang Autonomous County, Mianyang City, Sichuan Province (31.836 °N, 104.141 °E), Xiaojin County, Aba Tibetan and Qiang Autonomous Prefecture, Sichuan Province (31.625 °N, 102.315 °E) and Jinchuan County, Aba Tibetan and Qiang Autonomous Prefecture, Sichuan Province (31.527 °N, 101.639 °E), respectively. The predicted distance between COM and current suitable COM was 92. 970 m, 215,560 m and 275,440 m ; under the 2060s-SSP1-2.6, SSP2-4.5 and SSP5-8.5 scenarios, the predicted COM distances from the current suitable COM are 157,660 m, 248,210 m and 328,410 m, respectively.
In summary, under the 2040 s and 2060 s scenarios, the COM in the suitable area of F. taipaiensis in China progressive northwestward displacement across all temporal projections, aligning with predicted regional isotherm migration rates (29 km/decade). Due to the impact of global warming and humidification. In addition, under similar climatic conditions, the degree of mass center migration is expected to increase with the increase of radiative forcing.

Geographical distribution changes of the centroid of the suitable area of F. taipaiensis under different climate scenarios, the map creation using ArcMap v10.2(URL: https://www.arcgis.com/index.html).
Changes in suitable areas with time and climate
Projected habitat dynamics of F. taipaiensis exhibit significant temporal and scenario-dependent variability, as quantified in Table 5 and visualized in Fig. 7. The 2060s-SSP2-4.5 scenario presents two countervailing patterns: Firstly, habitat Contraction, a net loss of 11.74 km2 (25.47% contraction rate) through conversion of suitable habitat to marginal zones, primarily concentrated in elevational transition areas (1800–2200 m asl). Then, range Expansion, Emergence of 23.76 km2 novel suitable habitat (51.57% gain) along northwestward elevational gradients (2,600-3,100 m asl).The suitable areas were primarily contracted in central and northeastern China, including Liaoning, Shandong, Henan, Hubei, Shaanxi, and Chongqing Provinces (Fig. 7), and expansion to western China, including Gansu, Sichuan, Tibet, and other provinces, was observed. In general, the suitable area for F. taipaiensis varied with climate, and the distribution of F. taipaiensis changed similarly regardless of differences in climate. The change in the suitable area for F. taipaiensis was most pronounced in the SSP5-8.5 scenario and least pronounced in the SSP1-2.6 scenario. In the SSP2-4.5 scenario, the suitability area increased. In general, the impact of future climate change on the distribution pattern of F. taipaiensis is not significant, but there are differences in the magnitude and trend of changes under different climate scenarios.

Spatial changes of geographical distribution of F. taipaiensis in SSP126 (A,D), SSP245 (B,E), SSP585 (C,F) scenarios in the 2040s (A, B, C) and 2060s (D, E, F), the map creation using ArcMap v10.2 (URL: https://www.arcgis.com/index.html).
Dominant Climatic factors constraining the F. taipaiensis distribution
Habitat suitability and environmental factors were analysed using the jackknife method. The environmental factors affecting the F. taipaiensis distribution are shown in Table 6, and the 5 most important factors were mean temperature of the driest quarter(42.1%), mean diurnal range (mean of monthly (max temp-min temp))(21.1%) carbonate or lime content(13.2%), mean temperature of the warmest quarter(10.2%), and topsoil gravel content(4.8%). The relative contributions and significance of the different environmental variables were determined. Environmental factors that greatly constrained the F. taipaiensis distribution included temperature, temperature difference, carbonate or lime content, and topsoil gravel content.
Environmental characteristics of the suitable area for F. taipaiensis
The suitable areas for species primarily depend on primary environmental factors. The environmental factors affecting the distribution of F. taipaiensis can be clarified by modelling with the 10 most important environmental factors and plotting single-factor response curves (Fig. 8). The environmental factor with POS > 0.5 was suitable for plant growth. When the average temperature in the driest season (bio9) was less than − 6 °C, POS was less than 10%. The POS increased as the temperature increased and peaked at −1 °C. When the temperature exceeded − 2 °C, the POS of F. taipaiensis decreased as the temperature increased, and it was > 0.5 when the temperature was between − 6 and 5 °C. When the MDTD (bio2) was greater than 12.5 °C, the POS was less than 0.1. The POS was > 0.5 when the temperature difference was less than 8.5° C.
Similar to the driest quarterly mean temperature and the MDTD, the hottest quarterly mean temperature (bio10), carbonate or lime content (T_caco3) all had the following optimal ranges: Carbonate or lime content, 0%; hottest quarterly mean temperature, −7.5 to 20.5 °C. Survival probabilities of less than 0.5 were observed in areas with excessively high or low values for these variables. This is consistent with the environmental characteristics of the suitable area for F. taipaiensis under current conditions determined in the analysis of the primary environmental factors (Table 6), which indicates that the accuracy of the predictions was high.

Single factor response curve of current climate in F. taipaiensis.
As shown in Table 7, in the analysis of these factors, under the SSP2-4.5 scenario, the average temperature in the driest quarter decreased over time; in the other two climate scenarios, the average temperature in the driest quarter increases with time. In the SSP5-8.5 scenario, the mean diurnal range remains unchanged over time; in the other two climate scenarios, the mean diurnal range increases over time. Under all three climate scenarios, the average temperature of the hottest quarter decreased over time, with the largest decrease in the SSP1-2.6 scenario. Under all three climate scenarios, the carbonate or lime content remained basically unchanged. In the future climate scenario, the temperature in the suitable area of F. taipaiensis will decrease.
Analysis of niche differences under various climate scenarios
The ecological niche space of F. taipaiensis under various climate patterns was examined. High ecological niche overlap was observed among the different climatic scenarios (maximum ecological niche overlap 2040s-SSP1-2.6 = 0.656; minimum ecological niche overlap 2060s-SSP5-8.5 = 0.466) (Fig. 9). Therefore there was no clear ecotope differentiation. In addition, the ecological niche overlap gradually decreased as the radiation intensity increased during the same period, and the ecological niche overlap was significantly lower in the SSP5-8.5 scenario than in the other two climate scenarios. The ecotope overlap of F. taipaiensis was lower in the 2060s than in the 2040s regardless of the radiation intensity, suggesting that there will be a decrease in the resources for common use in the future and the present.

Niche differences of F. taipaiensis in different climatic backgrounds in the future.
The ecotope widths of F. taipaiensis under the different climatic scenarios were calculated using the ENMTools software package. The max and min values of B1 were 5.4% and 4.1%, respectively, and those for B2 were 85.1% and 83.3%, respectively (Table 8). Thus, no significant differences were observed in each period, indicating that F. taipaiensis has the potential to be a widespread species. In addition, both B1 and B2 increased under other climate scenarios, suggesting that the variety of resources available to F. taipaiensis in future climates and its adaptability to future environments will increase.
Discussion
Model simulation evaluation
The Maxent model performance can be improved by adjusting model parameters, and the accuracy depends on the improvement of the fit between current and predicted areas and visual inspection of the predicted distributions39. Maxent models based on default parameters can be overly complex and can result in high goodness-of-fit. In species distribution modelling studies, default parameter-based analyses have been performed using only climatic factors or combinations of climatic, topographic, and soil factors. Optimization of Maxent parameters in previous studies has generally only been performed for models that were based exclusively on climatic factors. As a result, species distributions in most studies have been assessed using default parameters. Such analyses are prone to overfitting and are highly complex, possibly reducing the accuracy of the results or even producing results that are difficult to interpret, such as highly variable environmental factor response curves. Therefore, Maxent models require optimization. Parameter-optimized Maxent models are capable of avoiding overfitting and enhancing species distribution prediction accuracy. The smoother response curves approached the normal distribution curve in a manner consistent with Shelford’s tolerance law40. The optimized Maxent model exhibited strong reliability and effectiveness in simulating the potential geographical distribution of F. taipaiensis41.
According to other related studies, the number of species occurrence records and the concentration of distribution points have a major effect on the simulation accuracy of species distribution models. Generally, the model simulation accuracy increases with the sample size, but the increase in simulation accuracy eventually stagnates42. An excessively high number of distribution points can result in overfitting due to spatially autocorrelated distribution points, which can affect the model simulation results35. The ENMTools package in R has the function of removing redundancy of distribution points: removing spatially clustered distribution points by buffer sampling to avoid overfitting.
We obtained 53 F. taipaiensis occurrence records, and we analysed the relevance of the distribution point data to environmental variables. A total of 23 occurrence records were retained, and this screening process resolved the problems associated with inaccurate modelling results stemming from small sample sizes and covariance between environmental factors. In some studies, changes in the size of the study area can alter the background data, which in turn affects model construction43,44. We conducted a national-scale prediction of the potential distribution area of F. taipaiensis. Studies conducted on larger or smaller scales should compare the background data points with species occurrence points instead of randomly extracting background points from the study area. Nevertheless, more studies are needed to determine the optimal methods for selecting reasonable background data points. Currently, there is no uniform standard for classifying the selection area, and the method for selecting the threshold value has a direct effect on the size of the predicted suitable area; this can possibly result in large differences between the inferred highly suitable habitat area and the actual distribution of species. Thus, an appropriate threshold value for classification needs to be selected and compared against the actual distribution area33.
Environmental factors constraining the distribution of F. taipaiensis
Various topographic factors, including elevation, slope, and slope orientation, have the strongest effect on wildlife diversity in mountainous areas10. Elevation can regulate light and temperature, which can affect the distribution of vegetation45. Slope has a direct effect on the flow and distribution of water, light energy reception, soil temperature distribution, and other hydrothermal conditions46. Slope orientation affects the solar radiation intensity and frequency and the spatial distribution of precipitation, which thereby affects the light and water conditions for montane plants and hence their diversity47. Overall, these topographic factors directly affect spatial variation in solar radiation, precipitation, soil, and nutrients, which increases spatial heterogeneity, as well as the functional and structural diversity of the mountain community; these factors also provide ecological niches that support the survival of mountain plants48.
Under current climatic conditions, temperature had major effects on the distribution of F. taipaiensis; this was followed by soil factors. Under current conditions, F. taipaiensis mainly grew in the central and western regions, including Sichuan, Yunnan, Gansu, Shaanxi, Henan, Hubei, Guizhou, Xizang, Chongqing, and Shandong Provinces, which are the areas where F. taipaiensis has been traditionally produced; the predicted results were consistent with the actual distribution of F. taipaiensis in China49. Zou Meng et al.3. proposed that MDTD had the strongest effect on the distribution of F. taipaiensis; other temperatures and precipitation conditions were more important in our study, and the effects of altitude and slope on F. taipaiensis in light of its biology were also examined.
In the factor response curves, the optimal habitats for F. taipaiensis had the following characteristics: average temperature in winter of − 1 °C, and average temperature in summer of 14 °C. This was consistent with the biological characteristics of F. taipaiensis, which is known to prefer cool and humid climates and is mostly found in the humid understory of broad-leaved forests, shrubby belts, and forest clearings49. Changes in the distribution of F. taipaiensis under warming conditions were primarily caused by the temperature anomalies. Changes in the driest quarterly mean temperature also reduced the suitability of the low-latitude region, and a higher MDTD was less favourable for the survival of F. taipaiensis in low-latitude regions. F. taipaiensis resources at low latitudes should be monitored to obtain timely information on emerging threats, aid the conservation of F. taipaiensis habitats and resources, and aid the construction of a long-term F. taipaiensis seed resource bank.
Changes in the potential geographical distribution of F. taipaiensis
A robust method has been used to assess the effects of future climate change on the distribution of F. taipaiensis50. The future population dynamics of F. taipaiensis were inferred by simulating its predicted distribution over time. A probability of existence > 0.5 means that the corresponding ecological factors were consistent with observations of plant growth51. Climate warming can affect the suitable habitat area for F. taipaiensis, and temperature and precipitation were the most significant variables. Both temperature and precipitation affected the large-scale geographic distribution of F. taipaiensis. Hydrothermal conditions primarily constrain the potential distribution of F. taipaiensis in China, and soil variables also have profound effects on the potential distribution of F. taipaiensis. Analysis of environmental factors revealed that temperature and soil variables jointly affect the potential distribution of F. taipaiensis.
The potential distribution of F. taipaiensis in China under future climate scenarios differed from that under current climate conditions, indicating that future warming affected the distribution of F. taipaiensis. The total suitable area under the 2040s scenarios and 2060s scenarios decreased, indicating that the reduction in the distribution of F. taipaiensis is accelerating over time; this is accompanied by the intensification of global warming. Changes in the distribution of F. taipaiensis driven by climate change were generally consistent; the area of suitable habitat for F. taipaiensis increased in size, and the overall COM of the potential distribution shifted to the northwest. Zou Meng et al.3. predicted the potential distribution of F. taipaiensis under current conditions, but they did not compare it with that under future climate conditions in different periods. Our study examined changes in the suitable area for F. taipaiensis and the COM of its potential distribution. In our study, the highly suitable area and spatial distribution of F. taipaiensis changed significantly under future climate scenarios. Changes in the distribution of F. taipaiensis were consistent with northward shifts in the geographical distribution of other species affected by global warming52. The ecotope overlap size denotes similarity in plant resource use, and large ecotope overlap suggests that species have similar ecological needs, similar resources, and similar biological characteristics53.
The suitable habitat areas for F. taipaiensis were classified into areas where the suitable habitat expanded, contracted, and remained unchanged. The suitable habitat area for F. taipaiensis reduced overall under future climate change scenarios. The area expanded northwest and contracted in the southern and eastern regions under current conditions. The contraction rate of the suitable habitat for F. taipaiensis was greater than the rate of expansion. Changes in its geographic distribution were more pronounced at the boundaries of the suitable habitat area. The total suitable habitat areas for F. taipaiensis decreased significantly under the 2060s-SSP5-8.5 scenario, which further indicated that climate change affected the distribution of F. taipaiensis. As the climate change became more severe in the future, spatial variation in the suitable habitat areas for F. taipaiensis decreased, and the distribution of F. taipaiensis was more deeply affected; the suitable habitat areas for F. taipaiensis changed positively over time within the RCPs. Suitable habitat areas contracted at the boundaries of the suitable area, which coincides with the region where species tend to be most sensitive to changes in climate54,55.
Reductions in the future and current availability of resources suggest that the geographic distribution of F. taipaiensis will shift due to changes in global climate; this is consistent with the prediction results of the Maxent model indicating that the COM of the suitable habitat area for F. taipaiensis generally shifted to the northwest. The ecotope width of F. taipaiensis was lower under future climate scenarios than in the current period, indicating that the suitable area decreased under global warming; this was also consistent with the predictive results of the Maxent model and verified their accuracy33. The future suitable habitat area for F. taipaiensis was predicted assuming that soil and topographic factors would remain unchanged within 60 years31. Our study was focused on national and provincial areas, and our findings were not effective for making local-scale predictions subjected to microclimatic variation. The physiological and morphological characteristics of Fritillaria species were similar, and their geographical distributions may shift in a similar manner regardless of the climate. Our model could also aid predictions of the suitable area for other Fritillaria species under future climate scenarios. The study of Deya Huang et al. Predicted that temperature seasonality had the greatest impact on F. taipaiensis, which was also consistent with the prediction results of environmental factors in this study56.
Implications for biodiversity conservation and limitations
According to previous studies, species distribution modelling can have important implications for biodiversity conservation, as the predictions can facilitate the identification of conservation priority areas10,57. To aid biodiversity conservation planning efficiency, we performed scientifically valid assessments, followed by comprehensive field surveys58. Maxent modelling can be used to forecast the potential distributions of species using a limited number of species occurrence records, and this can provide insights into the mechanisms driving changes in the species distributions with climate changes, even when as little as five occurrence records are used59. The species F. taipaiensis exhibits high environmental sensitivity and requires specific bioclimatic conditions for optimal growth. Future investigations should prioritize the following multidisciplinary approaches: (1) Conducting mechanistic studies to elucidate the environmental drivers of geo-authentic medicinal material formation, with particular emphasis on quantifying parameter thresholds for critical growth factors; (2) Developing climate-resilient cultivation strategies through integration of regional climate change projections with precision agriculture technologies, ensuring pharmaceutical quality standards in potential cultivation zones; (3) Enhancing habitat suitability prediction models by incorporating anthropogenic variables along socioeconomic dimensions, in addition to fundamental edaphic and climatic parameters; (4) Implementing spatial optimization frameworks that account for infrastructure development impacts, including transportation networks and industrial zoning patterns on species distribution dynamics.
Maxent-based species distribution modelling in biodiversity conservation also has its limitations. Long-term and sustainable conservation strategies require continuous monitoring and biodiversity assessments60. Study of changes in the potential distributions of species in response to climate changes is critically important for ongoing monitoring efforts. Although the Maxent model is capable of effectively forecasting the potential distributions of a large number of species of conservation concern, including F. taipaiensis in this study, field surveys should be conducted in the future to acquire more accurate and comprehensive species information, which would enhance the simulation accuracy of the potential distributions of species61.
Potential limitations of different factors
The thresholds only account for the general effects of single environmental variables, instead of diverse environmental variables, on the potential distributions of F. taipaiensis. Moreover, the interactions between individual variables are mutually constraining. Hence, the thresholds of each environmental variable mutually affect each other and may change when the variables are jointly considered in practical applications62.
Many factors affect plant growth, and the uncertainties associated with the above factors can affect predictions of the distribution of F. taipaiensis. The ecological environment of F. taipaiensis has deteriorated due to anthropogenic activities, such as the harvesting of medicinal herbs and deforestation, and many populations have declined and become progressively fragmented, which has had a marked effect on the survival and habitat of F. taipaiensis species.
In addition, improving the suitability of habitats for plants is a long-term process. For example, typhoons can restrict the growth of M. wufengensis in East China during the study period. There is thus a need to identify important ecological factors based on the results of tests of plant growth in the field. These considerations can enhance the accuracy of predictions of the distribution of F. taipaiensis under climate change.
Conclusions
F. taipaiensis is primarily distributed in western China, including Shaanxi, Chongqing, and Sichuan Provinces. Temperature conditions mainly affect the distribution of F. taipaiensis, especially seasonal temperatures. As global warming intensifies, F. taipaiensis populations will decrease, and some suitable areas will decrease in size and migrate to higher latitudes. Suitable habitats for F. taipaiensis are primarily located in northeastern Chongqing, southern Shaanxi, western Hubei, and northwestern Yunnan, which is basically consistent with the predicted areas in the Maxent model. Therefore, the effect of climate change on the COM of the potential F. taipaiensis distribution should be considered, and movements of the potential distribution to the northwest under current climate conditions were predicted.
The suitable areas for F. taipaiensis under future climate change were identified. These findings will aid the development of strategies to promote the adaptation of F. taipaiensis to climate change. F. taipaiensis is a widespread and dominant species in plant communities in Southwest China; it is thus a characteristic component of the Fritillaria sp. flora in Southwest China. These results have implications for studies of the evolution of Fritillaria in Southwest China, and accurate simulations of the potential distribution of Fritillaria have implications for optimizing their cultivation.
Identifying the abiotic factors (drought or rainy season precipitation) that affect species habitats is a major focus of ecological studies, and more studies are needed to identify the effects of environmental factors on the growth and development of species. Our results will aid future studies aimed at clarifying the responses of species to climate change.
Data availability
The authors declare that the data supporting the findings of this study are available within the paper and its Supplementary Information files. The study used and analysed publicly available datasets. Should any raw data files be needed in another format they are available from the corresponding author upon reasonable request.
References
Shaozhi, F., Dingming, Y., Yangjian, O., Jide, L. & Qifu, L. Discussion on the historical status and countermeasures of the development of Taibai Fritillaria plant industry in Chongqing. Lishizhen Med. Materia Med. Res. 25 (2), 476–477 (2014).
Min, L. et al. Esearch progress in medicinal plant Fritillaria taipaiensis P. Y. Li. Chin. Wild Plant. Esources. 40 (02), 42–45. https://doi.org/10.3969/j.issn.1006-9690.2021.02.008 (2021).
Meng, Z., Ruiping, J., Jialun, L., Qinyang, D. & LiaoHai; Jiayu, Z. Prediction of potential distribution of Fritillaria taipaiensis based on maxent model. Chin. J. Inform. Traditional Chin. Med. 28 (09), 1–5. https://doi.org/10.19879/j.cnki.1005-5304.202101042 (2021).
The white paper on “China’s Biodiversity Conservation”. In (NFGA), N. F. a. G. A., Ed. (2021).
Alan Pounds, J. et al. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 439 (7073), 161–167. https://doi.org/10.1038/nature04246 (2006).
Craparo, A., Van Asten, P. J., Läderach, P., Jassogne, L. T. & Grab, S. Coffea arabica yields decline in Tanzania due to climate change: Global implications. Agric. For. Meteorol. 207, 1–10. https://doi.org/10.1016/j.agrformet.2015.03.005 (2015).
Junsheng, L., Caiyun, Z., xiaopu, W. & Ran, Q. The impact of climate change on species distribution patterns. In Biodiversity and Ecological Security - The 13th Beijing International Science and Technology Industry Expo and the 7th China International Forum on High tech Science and Technology for Biodiversity Conservation and Utilization, Beijing, China, 2010 12 (2010).
Xu, H., Liu, S., Li, Y., Zang, R. & He, F. Assessing non-parametric and area‐based methods for estimating regional species richness. J. Veg. Sci. 23 (6), 1006–1012. https://doi.org/10.1111/j.1654-1103.2012.01423.x (2012).
Pilotto, F. et al. Meta-analysis of multidecadal biodiversity trends in Europe. Nat. Commun. https://doi.org/10.1038/s41467-020-17171-y (2020).
Wu, B. et al. Effect of habitat factors on the understory plant diversity of Platycladus orientalis plantations in Beijing mountainous areas based on maxent model. Ecol. Ind. 129, 107917. https://doi.org/10.1016/j.ecolind.2021.107917 (2021).
Mousikos, A., Manolaki, P., Knez, N. & Vogiatzakis, I. N. Can distribution modeling inform rare and endangered species monitoring in mediterranean Islands? Ecol. Inf. 66, 101434. (2021).
Shi, X. et al. Prediction of potentially suitable areas for the introduction of Magnolia wufengensis under climate change. Ecol. Ind. 127, 107762. https://doi.org/10.1016/j.ecolind.2021.107762 (2021).
Zhonglin, X., Huanhua, P. & Shouzhang, P. The development and evaluation of species distribution models. Acta Ecol. Sin. 35 (02), 557–567. https://doi.org/10.5846/stxb201304030600 (2015).
Jiejie, S. et al. A prediction on the potential suitable areas of Zelkova Schneideriana in Zhejiang Province using maximum entropy model. For. Resour. Manag. (04), 37–45. https://doi.org/10.13466/j.cnki.lyzygl.2019.04.006 (2019).
Manle, Z. et al. Assessment of potential distribution for endangered plant Firmiana danxiaensis based on maxent modeling. Ecol. Sci. 41 (05), 55–62. https://doi.org/10.14108/j.cnki.1008-8873.2022.05.007 (2022).
Phillips, S. J. & Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 31(2), 161–175 https://doi.org/10.1111/j.0906-7590.2008.5203.x (2008).
Ran, L., Chun-Jing, W., Jian, H. & Zhi-Xiang, Z. Analysis of geographical distribution of Abies in China under climate change. Bull. Bot. Res. 38 (01), 37–46. https://doi.org/10.7525/j.issn.1673-5102.2018.01.005 (2018).
Elith, J. et al. A statistical explanation of maxent for ecologists. Divers. Distrib. 17 (1), 43–57. https://doi.org/10.1111/j.1472-4642.2010.00725.x (2011).
Li, J., Fan, G. & He, Y. Predicting the current and future distribution of three Coptis herbs in China under climate change conditions, using the maxent model and chemical analysis. Sci. Total Environ. 698, 134141. https://doi.org/10.1016/j.scitotenv.2019.134141 (2020).
Wang, F., Yuan, X., Sun, Y. & Liu, Y. Species distribution modeling based on maxent to inform biodiversity conservation in the central urban area of Chongqing municipality. Ecol. Ind. 158, 111491. https://doi.org/10.1016/j.ecolind.2023.111491 (2024).
Tao, S., Hong, Y. & Yuan-zhong, W. Geographical distribution and bioclimatic characteristics of the wild Gentiana rigescens resources. Chin. J. Appl. Ecol. 30 (07), 2291–2300. https://doi.org/10.13287/j.1001-9332.201907.003 (2019).
Xu, D., Zhuo, Z., Wang, R., Ye, M. & Pu, B. Modeling the distribution of Zanthoxylum armatum in China with MaxEnt modeling. Global Ecol. Conserv. 19, e00691 https://doi.org/10.1016/j.gecco.2019.e00691 (2019).
Panda, R. M. & Behera, M. D. Assessing harmony in distribution patterns of plant invasions: A case study of two invasive alien species in India. Biodivers. Conserv. 28, 2245–2258. https://doi.org/10.1007/s10531-018-1640-9 (2019).
Ramos, R. S., Kumar, L., Shabani, F. & Picanço, M. C. Risk of spread of tomato yellow leaf curl virus (TYLCV) in tomato crops under various climate change scenarios. Agric. Syst. 173, 524–535. https://doi.org/10.1016/j.agsy.2019.03.020 (2019).
Tianfeng, W. et al. Characteristics and controls of vegetation and diversity changes monitored with an unmanned aerial vehicle (UAV) in the foreland of the Urumqi glacier 1, Tianshan, China. Sci. Total Environ. 771, 145433. https://doi.org/10.1016/j.scitotenv.2021.145433 (2021).
Ruiping, J., et al. Modeling of the potential geographical distribution of three fritillaria species under climate change. Front. Plant Sci. 12, 749838 https://doi.org/10.3389/fpls.2021.749838 (2022).
Zhong-lin, L. et al. Study on plant niche under fencing measures in the semi-arid grassland of North China. Acta Agrestia Sinica. 22 (06), 1186–1193. https://doi.org/10.11733/j.issn.1007-0435.2014.06.006 (2014).
Xuefeng, H. et al. Study on niches of 7 Stipa species in inner Mongolia. Acta Agrestia Sinica. 29 (12), 2778–2784. https://doi.org/10.11733/j.issn.1007-0435.2012.12.018 (2021).
Tongwen, W. et al. BCC-CSM2-HR: A high-resolution version of the Beijing climate center climate system model. Geosci. Model Dev. 14 (5), 2977–3006. https://doi.org/10.5194/gmd-14-2977-2021 (2021).
Tong, J., Lü, Y., Jinlong, H., Wang, Y., Buda, S. & Hui, T. New scenarios of CMIP6 Model(SSP-RCP)and its application in the Huaihe river basin. Adv. Meteorol. Sci. Technol. 10 (05), 102–109. https://doi.org/10.3969/j.issn.2095-1973.2020.05.016 (2020).
Yin-bo, Z., Chen-hong, G. & Hao, Q. Prediction of the suitable distribution and responses to climate change of Elaeagnus mollis in Shanxi Province, China. Chin. J. Appl. Ecol. 29 (04), 1156–1162. https://doi.org/10.13287/j.1001-9332.201804.011 (2018).
Radosavljevic, A. & Anderson, R. P. Making better maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 41 (4), 629–643. https://doi.org/10.1111/jbi.12227 (2014).
Qi, W., Bao-guo, F. & Guang-hua, Z. Prediction of potential distribution area of Corylus Mandshurica in China under climate change. Chin. J. Appl. Ecol. 39 (11), 3774–3784. https://doi.org/10.13292/j.1000-4890.202011.014 (2020).
Haoxiang, Z. Response of Potential Geographical Distribution of Eight China’s First-Class Rare and Endangered Plants to Climate Change and Analysis of GAP (Master of Engineering, Northwest Normal University, 2021).
Zhengyu, Y., Qifei, H. & Bin, L. Prediction of distribution area of main noxious and miscellaneous weeds in Xinjiang based on maxent model. Acta Ecol. Sin. 43 (12), 5096–5109. https://doi.org/10.5846/stxb202205061252 (2023).
Guanghua, Z. et al. Analysis of the distribution pattern of Chinese Ziziphus jujuba under climate change based on optimized biomod2 and MaxEnt models %J Ecological Indicators. Ecol. Indic. https://doi.org/10.1016/j.ecolind.2021.108256 (2021).
Han, Y., Yun-Ling, Z., Song-Mei, M., Chun-Cheng, W. & Dan, Z. Suitable distribution simulation and local environmental adaptability differentiation of Lycium ruthenicum in Xinjiang,China. Chin. J. Plant. Ecol. 45 (11), 1221–1230. https://doi.org/10.17521/cjpe.2021.0179 (2021).
Warren, D. L., Glor, R. E. & Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution 62 (11), 2868–2883 https://doi.org/10.1111/j.1558-5646.2008.00482.x (2008).
Jie, G., Qin, Z., Dongfang, Z., Chengzhong, W. J. S. & Caixiang, X. Potential distribution area and ecological characteristics of common buckwheat Fagopyrum Dibtrys (D. Don) Hara. J. Plant. Prot. 45 (03), 489–495. https://doi.org/10.13802/j.cnki.zwbhxb.2018.2016185 (2018).
Phillips, S. J., Anderson, R. P., Dudík, M., Schapire, R. E. & Blair, M. E. Opening the black box: An open-source release of Maxent. Ecography 40(7), 887–893 https://doi.org/10.1111/ecog.03049 (2017).
Sheu, Y. et al. evolution, the combined role of dispersal and niche evolution in the diversification of Neotropical lizards. Ecol. Evol. 10 (5), 2608–2625. https://doi.org/10.1002/ece3.6091 (2020).
Chen, X. & Lei, Y. Effects of sample size on accuracy and stability of species distribution models: A comparison of GARP and maxent. In Recent Advances in Computer Science and Information Engineering: Volume 2 (eds Qian, Z., Cao, L., Su, W., Wang, T. & Yang, H.) 601–609 (Springer Berlin Heidelberg: Berlin, Heidelberg, 2012).
Qianzhao, J. et al. Effects of sample size and study range on accuracy of maxent in predicting species distribution: A case study of the black-and-white snub-nosed monkey. Acta Theriol. Sinica 39 (02), 126–133. https://doi.org/10.16829/j.slxb.150203 (2019).
Wendi, M. & Shizong, L. Development of maximum entropy model at home and abroad and its applications in different Climatic backgrounds and regional scales. Forestry Constr. (02), 32–40 (2023).
Galván-Cisneros, C. M., Villa, P. M., Coelho, A. J., Campos, P. V. & Meira-Neto, J. A. Altitude as environmental filtering influencing phylogenetic diversity and species richness of plants in tropical mountains. J. Mt. Sci. 20 (2), 285–298. https://doi.org/10.1007/s11629-022-7687-9 (2023).
Pan, J., Bai, Z., Cao, Y., Zhou, W. & Wang, J. Influence of soil physical properties and vegetation coverage at different slope aspects in a reclaimed dump. Environ. Sci. Pollut. Res. 24, 23953–23965. https://doi.org/10.1007/s11356-017-9974-5 (2017).
Cantón, Y., Del Barrio, G., Solé-Benet, A. & Lázaro, R. Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain. CATENA 55(3), 341–365 https://doi.org/10.1016/S0341-8162(03)00108-5 (2004).
Parker, A. J. The topographic relative moisture index: An approach to soil-moisture assessment in mountain terrain. Phys. Geogr. 3(2), 160–168 https://doi.org/10.1080/02723646.1982.10642224 (1982).
Cong, X., Chen, H., Li, W., Li, D. & Li, S. Habitat investigation and rhizosphere soil analysis of wild Fritillaria taibaiensis. J. Agric. 12 (03), 55–58. https://doi.org/10.11923/j.issn.2095-4050.cjas2021-0133 (2022).
Sharafati, A. & Pezeshki, E. A strategy to assess the uncertainty of a climate change impact on extreme hydrological events in the semi-arid Dehbar catchment in Iran. Theoret. Appl. Climatol. 139 (1), 389–402. https://doi.org/10.1007/s00704-019-02979-6 (2020).
Xiang, J. et al. Assessing the suitable distribution area of Pinus koraiensis based on an optimized maxent model. Chin. J. Ecol. 38 (08), 2570–2576. https://doi.org/10.13292/j.1000-4890.201908.017 (2019).
Yin-bo, Z., Yan-lan, L., Hao, Q. & Qing-xin, M. Prediction on spatial migration of suitable distribution of Elaeagnus mollis under climate change conditions in Shanxi Province, China. Chin. J. Appl. Ecol. 30 (02), 496–502. https://doi.org/10.13287/j.1001-9332.201902.040 (2019).
Yuan, Y., Tang, X., Liu, M., Liu, X. & Tao, J. Species distribution models of the spartina alterniflora Loisel in its origin and invasive country reveal an ecological niche shift. Front. Plant Sci. 12, 738769. https://doi.org/10.3389/fpls.2021.738769 (2021).
Diamond, S. E., Frame, A. M., Martin, R. A. & Buckley, L. B. Species’ traits predict phenological responses to climate change in butterflies. Ecol. Soc. Am. 92 (5), 1005–1012. https://doi.org/10.1890/10-1594.1 (2011).
Thuiller, W., Lavorel, S. & Araújo, M. B. Niche properties and geographical extent as predictors of species sensitivity to climate change. Glob. Ecol. Biogeogr. 14 (4), 347–357. https://doi.org/10.1111/j.1466-822X.2005.00162.x (2005).
Deya, H. et al. Biomod2 modeling for predicting the potential ecological distribution of three Fritillaria species under climate change. Sci. Rep. 13 (1), 18801. https://doi.org/10.1038/s41598-023-45887-6 (2023).
Merchant, D. et al. Distribution modeling and gap analysis of shorebird conservation in Northern Brazil. Remote Sens. 15 (2), 452. https://doi.org/10.3390/rs15020452 (2023).
Wang, Z., Zhang, B., Zhang, X. & Tian, H. Using maxent model to guide marsh conservation in the Nenjiang river basin, Northeast China. Chin. Geogra. Sci. 29, 962–973. https://doi.org/10.1007/s11769-019-1082-1087 (2019).
Xian, Y., Lu, Y. & Liu, G. Is climate change threatening or beneficial to the habitat distribution of global Pangolin species? Evidence from species distribution modeling. Sci. Total Environ. 811, 151385. https://doi.org/10.1016/j.scitotenv.2021.151385 (2022).
Zhongde, H., Yang, B. & Zhangqian, M. A. J. Y., Mapping biodiversity conservation priorities for protected areas: A case study in Xishuangbanna Tropical Area, China. Biol. Conserv. https://doi.org/10.1016/J.BIOCON.2020.108741 (2020).
Bingran, M. et al. Boundary delineation and grading functional zoning of Sanjiangyuan National park based on biodiversity importance evaluations. %J the science of the total environment. Sci. Total Environ. 825, 154068–154068. https://doi.org/10.1016/j.scitotenv.2022.154068 (2022).
Zhou, D., Zhang, R., Sun, W., Zhang, J. & Wang, H. Evaluation of the suitability and influencing factors of winter rapeseed planting in Gansu Province. Chin. J. Eco-Agric. 22 (6), 697–704. https://doi.org/10.3724/SP.J.1011.2014.40153 (2014).
Acknowledgements
We thank Dr. Nong Zhou and Dr. Hua Zhang for providing suggestions that helped improve this manuscript. We thank Dr. Junsheng Qi and Wei Lin for providing assistance during this work.
Funding
This work was completed with support from the Science and Technology Research Project of Chongqing Education Commission (KJZD-M202301204).
Author information
Authors and Affiliations
Contributions
YHW: Conceptualization, methodology, software, formal analysis, investigation and writing - original draft. XYR: Writing - review and editing, supervision, methodology and resources. KW: Resources, Investigation. WL: Software, Data curation and Visualization. PPW: methodology and formal analysis. ZYL: Resources and Validation. HZ: Funding acquisition, supervision, review and editing. NZ: writing, supervision, review and editing. The final version has obtained the approval of all authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Wang, Y., Ren, X., Wang, K. et al. Maxent model-based prediction of the potential distribution of Fritillaria taipaiensis P. Y. Li. Sci Rep 15, 20837 (2025). https://doi.org/10.1038/s41598-025-01682-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-025-01682-z

{kind=link}
{kind=link}