
Abstract
Neuropsychiatric symptoms (including anxiety, depression, apathy, impulse–compulsive behaviors and hallucinations) are among the most common non-motor features of Parkinson’s disease. Whether these symptoms should be considered as a direct consequence of the pathophysiologic mechanisms of Parkinson’s disease is controversial. Morphometric similarity network analysis and epicenter mapping approach were performed on T1-weighted images of 505 patients with Parkinson’s disease and 167 age- and sex-matched healthy participants from Parkinson’s Progression Markers Initiative database to reveal the commonalities and specificities of distinct neuropsychiatric symptoms. Abnormal cortical co-alteration pattern in patients with neuropsychiatric symptoms was in somatomotor, vision and frontoparietal regions, with epicenters in somatomotor regions. Apathy, impulse–compulsive behaviors and hallucinations shares structural abnormalities in somatomotor and vision regions, with epicenters in somatomotor regions. In contrast, the cortical abnormalities and epicenters of anxiety and depression were prominent in the default mode network regions. By embedding each symptom within their co-alteration space, we observed a cluster composed of apathy, impulse–compulsive behaviors and hallucinations, while anxiety and depression remained separate. Our findings indicate different structural mechanisms underlie the occurrence and progression of different neuropsychiatric symptoms. Based upon these results, we propose that apathy, impulse-compulsive behaviors and hallucinations are directly related to damage of motor circuit, while anxiety and depression may be the combination effects of primary pathophysiology of Parkinson’s disease and psychosocial causes.
Similar content being viewed by others

Introduction
Motor symptoms remain central to the diagnosis of Parkinson’s disease (PD), but neuropsychiatric symptoms (NPS) are gaining more attention as common and clinically important non-motor features [1], such as PD can now be conceptualized as a complex neuropsychiatric disorder [2]. These NPS can be divided into several categories of affect including anxiety and depression, perception and thinking (i.e., hallucinations and psychosis), and motivation (i.e., impulse–compulsive behaviors (ICBs) and apathy) [2].
These NPS are complex and diverse, making comprehensive study of the neuropathologic mechanisms of NPS in PD difficult. Whether they should be considered a direct consequence of the pathophysiologic mechanisms of PD, or a pseudospecific effect related to PD’s impact on psychosocial functioning and subsequent psychological sequelae [2]. For example, epidemiologically, patients with PD have more distinctive hallucinations and ICBs, which differ from the spectrum of neuropsychiatric disorders found in the general population, suggesting a link to PD pathophysiology. More ambiguous, however, is whether the anxiety and depression observed in PD are distinct from that in formal psychiatric disorders or even the general population [2,3,4]. Presentation of NPS, particularly depression and anxiety, in early PD can have a clear psychological or psychosocial causation, whereas the onset of these symptoms in advanced disease have been associated with widespread PD neuropathology or exposure to dopaminergic therapy [2].
Based on the clinical and epidemiologic characteristics of NPS [2,3,4], as well as previous neuroimaging studies of single NPS [5,6,7,8,9,10,11,12,13,14,15,16,17,18,19], we hypothesized that the various types of NPS will demonstrate different relationships to the known cortical core pathophysiological changes found in PD (e.g., motor cortex deterioration). We predict that select NPS unique to PD patient will be linked to evidence of the network spread of PD damage emanating from motor circuit, whereas depression and anxiety in PD will align more closely with psychological and social factors in ways similar to primary psychiatric disorders, and mainly accompanied by structural and functional damage in brain regions involved in advanced cognitive functions (e.g., the default mode network (DMN) regions such as the prefrontal cortex, the orbitofrontal cortex(OFC) and the anterior cingulate cortex) [5,6,7, 9, 20].
Our approach is to use structural neuroimaging to obtain a spectral characterization of PD-associated NPS, all toward the goal of clarifying if the independence or phenomenological overlap of NPS, mirrors a shared or distinct structural basis and etiology. This strategy has been applied to well-known neurodevelopmental (e.g., attention-deficit/hyperactivity disorder and autism spectrum disorder) and psychiatric disorders (e.g., generalized anxiety disorder, major depressive disorder, schizophrenia spectrum disorders and obsessive-compulsive disorder) [21, 22].
Morphometric similarity network (MSN) analysis combines multiple modalities or features that may provide insights into macroscale cortical organization in a single individual [23]. As a promising approach to characterize inter-regional anatomical similarity by leveraging within-subject variance of different morphometric properties [23], MSN reflects inter-region cytoarchitectonic similarity [24, 25], genetic similarity [23] and white matter connection [24, 26]. PD as a neurodegenerative disease in which pathologic protein aggregates are capable of spreading from focal areas of pathogenesis to nonadjacent areas via transsynaptic or transneuronal spreading along discrete brain networks [27,28,29]. Here we further employ the network diffusion model (i.e., the epicenter model) to speculate on whether certain brain regions could act as sources or epicenters of pathological diffusion through brain networks. Lastly, based upon our findings, we revealed the extent of the relationship of distinct NPS to the core cortical pathophysiologic damage imposed by PD.
Materials and methods
Participants
The data used in this study were obtained on February 2023 from the Parkinson Progression Markers Initiative (PPMI) [30] database via a standard application process. The PPMI study is an international multicentric cohort study designed to identify PD progression markers to improve understanding of disease etiology and course.
The inclusion and exclusion criteria of PD and Healthy participants (HPs) were described in the Supplementary Materials and Methods for details. Each PPMI recruiting center in this study received ethics approval from their local ethics board and obtained written informed consent from all participants in accordance with the declaration of Helsinki.
Assessments
The Movement Disorder Society Unified Parkinson’s Disease Rating Scale (MDS-UPDRS) Part III and Hoehn–Yahr staging was used to assess the motor impairments of PD [31]. Montreal Cognitive Assessment (MoCA) were used to assess cognitive decline.
Anxiety symptoms were assessed with the State-Trait Anxiety Inventory (S-TAI) [32]. Depression symptoms were assessed using the 15-item Geriatric Depression Scale (GDS-15) [33]. The Short version of the Questionnaire for Impulsive-Compulsive Disorders in Parkinson’s Disease (QUIP-S) screened for impulse control and compulsive disorder symptomatology [34]. Lastly, apathy and hallucinations were assessed with single items from the MDS-UPDRS Part I (see Supplementary Materials and Methods for details).
MRI Data pre-processing
MRI acquisition, MRI scanner vendors and scan sequences differed between the 35 PPMI acquisition sites (see Supplementary Materials and Methods for details). The 3D T1-weighted images were pre-processed in surface-based space using FreeSurfer (version 6.0, http://surfer.nmr.mgh.harvard.edu/). Briefly, the cortical surface was reconstructed through skull stripping, segmentation of brain tissue, separation of hemispheres, and construction of the grey/white interfaces and the pial surfaces [23, 35]. To check for differences in motion and image quality between groups, the Euler number was calculated for each T1-weighted image [36] (Fig. S1).
Construction of the MSN
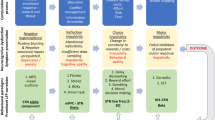
Morphometric features from each T1-weighted images were extracted from the Glasser atlas, 360 cortical regions constructed using multimodal MRI images from the Human Connectome Project [37]. Features were extracted including surface area, cortical thickness, grey matter volume, intrinsic (Gaussian) curvature, mean curvature, curved index and folding index (Fig. 1A) [23, 38]. The morphometric similarity between each possible pair of regions was estimated by Pearson’s correlation between their morphometric feature vectors (Fig. 1B, C) [23, 38,39,40]. To harmonize the center effect, ComBat was used on MSN [41]. W-score produced a normalized MSN change for each patient relative to controls, with normal aging and sex effects accounted for (Fig. 1D) [42]. The detailed process was described in the Supplementary Materials and Methods. Regional MSN change was equivalent to the sum of the W-scores of a given region to all other regions (Fig. 1E) [40, 43].

A Seven cortical morphometric features were extracted from T1-weighted images by FreeSurfer. The morphometric similarity between each possible pair of regions was estimated by Pearson’s correlation between their morphometric feature vectors (B) to produce a 360 × 360 MSN for each participant (C). D W-score produced a normalized MSN change for each patient relative to controls, taking age- and gender-related brain deformation into account. E Regional MSN changes were equivalent to the sum of the W-score of a given region to all other regions. F Group-level analysis base on these regional MSN changes. MSN morphometric similarity network.
Analysis of the MSN
For regional MSN change, by considering nNPS (with no neuropsychiatric symptoms, including anxiety, depression, apathy, impulse-compulsive behaviors and hallucinations) as controls, we obtained Cohen’s d maps reflecting case-control differences for each NPS in regional MSN for 360 Glasser parcels (Fig. 1F). In addition, we performed network-level analysis on the twelve human brain’s macroscopic functional networks across the Glasser parcels [44] (primary visual network (VIS1), secondary visual network (VIS2), somatomotor network (SMN), cingulo-opercular network (CON), dorsal attention network (DAN), language network (LAN), frontoparietal network (FPN), auditory network (AUD), DMN, posterior multimodal network (PMM), ventral multimodal network (VMM), orbito-affective network (ORA)). These analyses were conducted using a mixed-effects model that regressed gender, age, and center as covariates while accounting for within-subjects effects using Surfstat [45] (https://math.mcgill.ca/keith/surfstat).
Specific epicenter identification
To identify the epicenters closely related to the structural alterations, we applied the disease epicenter mapping (ENIGMA toolbox [46] v.1.1.3; https://enigma-toolbox.readthedocs.io/en/latest/)approach using group-wise normalized functional and structural connectivity matrices based on the Glasser atlas from 207 Human Connectome Project healthy adults [22, 47,48,49]. By spatially correlating the healthy functional and structural connectivity characteristics of each region with specific whole-brain patterns of regional MSN alterations, specific epicenters were identified for each NPS condition. This approach was repeated systematically across the whole brain, assessing the statistical significance of the spatial similarity of each region’s functional and structural connectivity profiles to regional MSN Cohen’s d maps with 1000 times spatial permutation tests (spin test) [50]. Brain regions with higher spatial similarity indicated more likely a disease epicenter (at p < 0.05 after spin tests). The twenty most relevant brain regions were selected to represent the epicenters of functional and structural connectivity. At the network level, chi-square tests determined whether the epicenters occurred more frequently in twelve human brain’s macroscopic functional networks, focusing on differences in the occurrence frequency in intra- versus extra-networks.
Embedding of each NPS in shared or distinct MSN alteration space
First, we intended to investigate associations between individual NPS in terms of structural alterations. We examined the spatial similarity of effects within each NPS type by calculating Pearson’s correlation between any pair of Cohen’s d maps. The similarity of structural alterations between NPS may also be affected by co-comorbidity, and here we also evaluate the contribution of NPS comorbidities to MSN similarity by linear regression analysis.
Second, we utilized hierarchical clustering to group the NPS such that those within an obtained cluster have MSN similarity metrics “similar” to each other, but different from NPS in other clusters. This yielded NPS sub-groups with shared or distinct MSN alternations (i.e., abnormalities) relative to our nNPS comparison group. More specifically, hierarchical clustering was applied to the Cohen’s d maps of each NPS.
Lastly, we assessed the overlap between shared and distinct NPS epicenters in terms of number and plotted this with the Cohen’s d maps of NPS spatial similarity to form a two-dimensional space. Together, these analyses revealed how individual symptoms are embedded in relation to each other within a NPS coordinate frame.
Results
Data samples
We studied 505 patients with PD (336 males, mean age ± SD = 62.15 ± 9.75 years) and 167 HPs (106 males, mean age ± SD = 61.29 ± 11.31 years) from 35 centers in the PPMI database. For T1-weighted MRI data, 505 PD patients had images at baseline, 129 of them had images at 1-year follow-up, 97 of them had images at 2-year follow-up, and 84 of them had images at 4-year follow-up. For HPs, only the baseline T1-weighted MRI data were used.
Based upon evaluation of the NPS-related symptom and behavioral assessments, we identified a total of 232 PD patients with criterion NPS forming the following subgroups: PD patients with anxiety (n = 67), depression (n = 76), apathy (n = 106), ICBs (n = 133), and hallucinations (n = 31). The highest comorbidities was observed for depression and anxiety (48.96%). Details of overall demographic and clinical characteristics are shown in Table 1, more details of each NPS condition and the comorbidities rates between them are shown in Tables S1 and S2.
Shared and distinct MSN alterations in NPS
Utilizing nNPS as a comparison group, we obtained Cohen’s d maps reflecting case-control differences for PD with NPS and PD with each NPS (i.e., anxiety, depression, apathy, ICBs and hallucinations).
Individuals with NPS exhibited decreased MSN alterations in the bilateral dorsolateral prefrontal cortex (DLPFC), bilateral precentral gyrus, postcentral gyrus and bilateral occipital cortex compared to nNPS (Fig. 2A, B). Based on network-level analysis, NPS exhibited significantly decreased MSN alterations in the SMN (p < 0.05, FDR corrected) and VIS1 (p < 0.05, no corrected), and significantly increased MSN alterations in the FPN compared to nNPS (p < 0.05, FDR corrected, Fig. 2C).

A NPS and nNPS both showed MSN changes relative to HPs. B Compared with nNPS, MSN of NPS significantly decreased in the DLPFC, somatomotor cortex and occipital cortex, with significantly decrease in SMN and VIS1 and increase in FPN based on network-level analysis (C). Decreased MSN in DMN were observed in anxiety and depression (D, E), while apathy, ICBs and hallucinations were characterized by decreased MSN in SMN and VIS1 (F–H). *p < 0.05, no corrected; **p < 0.05, FDR corrected. ICBs impulse compulsive behaviors, HPs healthy participants, MSN morphometric similarity network; NPS neuropsychiatric symptoms, nNPS with no neuropsychiatric symptoms, DLPFC dorsolateral prefrontal cortex, VIS1 primary visual network, VIS2 secondary visual network, SMN somatomotor network, CON cingulo-opercular network, DAN dorsal attention network, LAN language network, FPN frontoparietal network, AUD auditory network, DMN default mode network, PMM posterior multimodal network, VMM ventral multimodal network, ORA orbito-affective network.
Both anxiety and depression showed decreased MSN in DMN. Although they had similar patterns of reduction, the significance of the reduction was weaker for anxiety (p < 0.05, no corrected, Fig. 2D) relative to depression (p < 0.05, FDR corrected, Fig. 2E). Apathy, ICBs and hallucinations shared significant MSN decrease in SMN (p < 0.05, FDR corrected). In addition, they also shared MSN alterations in vision network, with decreased MSN in VIS1 of apathy, ICBs and hallucinations (p < 0.05, FDR corrected), and increased MSN in VIS2 of ICBs (p < 0.05, FDR corrected) and hallucinations (p < 0.05, no corrected). Simultaneously, apathy exhibited significant increase in FPN, ICBs exhibited significant increase in AUD and PMM, and hallucinations exhibited significant decrease in DMN (p < 0.05, FDR corrected, Fig. 2F–H). See Table S3 for more details.
Noting that different NPS have different level of motor impairments and cognitive decline (Fig. S2), we added MDS-UPDRS-III scores and MoCA as covariates in the mixed-effects model. The results showed that the main brain regions with significant results did not change (Fig. S3), indicating that the specific structural alterations of these NPS were not significantly affected by the level of motor impairments and cognitive decline. In addition, we conducted two additional control experiments to include total intracranial volume in covariates (Fig. S4) and to include center effects in the calculation of W-scores (Fig. S5), and showed that neither of these effects affected the main results.
In addition, we also demonstrated the abnormalities compared to HPs, by averaging MSN W-score in each group, see Fig. 2A and Fig. S6 for details.
Shared and distinct NPS epicenters
Disease epicenters represented regions whose functional and structural connectivity profiles spatially resembled the NPS-related MSN Cohen’s d maps. Systematically investigating the connectivity profiles of 360 cortical seeds revealed somatomotor regions as potential functional and structural epicenters of shared alterations in NPS (Fig. 3A, B). The top twenty functional NPS epicenters were observed in the bilateral precentral and postcentral gyrus, left middle temporal gyrus and right OFC (Fig. 3A). The top twenty structural NPS epicenters were present in the bilateral postcentral and precentral gyrus, and right middle temporal gyrus (Fig. 3B). At the network level, we found that functional and structural epicenters were more likely to appear in intra-SMN regions rather than extra-SMN regions (p < 0.05, FDR corrected, Fig. 3C).

Functional and structural epicenters of shared alterations in NPS were mainly present in somatomotor regions (A, B) with a significantly higher probability of occurrence in SMN (C). In anxiety and depression, DLPFC, PCC, mPFC and OFC were detected as unique epicenters, with a significantly higher probability of occurrence in DMN (D, E). Somatomotor cortex emerged as unique epicenters in apathy, ICBs and hallucinations, with a significantly higher probability of occurrence in SMN (F–H). *p < 0.05, no corrected; **p < 0.05, FDR corrected. ICBs impulse-compulsive behaviors, NPS neuropsychiatric symptoms, DLPFC dorsolateral prefrontal cortex, mPFC medial Prefrontal Cortex, OFC orbitofrontal cortex, PCC posterior cingulate cortex, VIS1 primary visual network, VIS2 secondary visual network; SMN: somatomotor network, CON cingulo-opercular network, DAN dorsal attention network, LAN language network, FPN frontoparietal network, AUD auditory network, DMN default mode network, PMM posterior multimodal network, VMM ventral multimodal network, ORA orbito-affective network.
In anxiety and depression, spatial correlations between MSN maps and seed-based functional and structural connectivity profiles implicated the DLPFC, PCC, mPFC, OFC and right middle temporal gyrus as unique epicenters, with a significantly higher probability of intra-DMN as opposed to extra-DMN regions, while the significance was weaker for depression(p < 0.05, no corrected, Fig. 3E) relative to anxiety(p < 0.05, FDR corrected, Fig. 3D). Conversely, somatomotor regions (postcentral gyrus and precentral gyrus) emerged as unique epicenters in apathy, ICBs, and hallucinations in PD, with a significantly higher probability of occurrence in intra-SMN compared to extra-SMN regions (p < 0.05, FDR corrected, Figs. 3F–H and S7). Results of other networks were shown in Supplementary Results for details (Table S4).
Embedding of each NPS in shared or distinct MSN alteration space
We first conducted spatial similarity analyses on the MSN Cohen’s d maps of various NPS. We observed a clear similarity between anxiety and depression (r = 0.65), apathy and ICBs (r = 0.64) (Fig. 4A). Simultaneously, the comorbidity between each NPS were calculated and a higher comorbidity rate (48.96%) was found to exist between anxiety and depression (Fig. 4B). We used linear regression analysis to evaluate the contribution of NPS comorbidities in MSN similarity, and found that 34.1% of MSN similarity could be attributed to NPS comorbidities, mainly caused by comorbidities of anxiety and depression (Fig. 4C).

A Similarities of MSN Cohen’s d maps between different NPS. B Comorbidity between different NPS. C The linear regression analysis to evaluate contribution of NPS comorbidities in MSN Cohen’s d maps similarities. D Different NPS cluster together in a two-cluster solution by hierarchical clustering analysis. E A similarity framework between each NPS and their shared alterations based on the correlation between shared NPS Cohen’s d maps and each NPS Cohen’s d maps (x-axis) and the overlap between shared NPS and each NPS epicenters (y-axis). MSN morphometric similarity network, NPS neuropsychiatric Symptoms.
Further hierarchical clustering analysis confirmed that a cluster composed of apathy, ICBs and hallucinations, while anxiety and depression stayed separate (Fig. 4D). We finally constructed a similarity framework between shared and distinct alterations in NPS using the epicenters and similarity dimensions. It compared the degree of MSN similarity and overlap of epicenters between shared and distinct MSN alterations, proposing their embedding in the NPS co-alteration space. We found that apathy, ICBs and hallucinations are more similar to the co-alterations in both dimensions, whereas anxiety and depression showed disparate symptom-specific profiles (Fig. 4E).
Discussion
Our study reported coordinated effects of five NPS related to PD (anxiety, depression, apathy, ICBs and hallucinations) on morphometric profiles and the embedding of each NPS within their shared or distinct brain structural alteration space. Specifically, morphometric similarity mapping disclosed the abnormal cortical co-alteration patterns affecting multiple regions in NPS patients. By using abnormal MSN patterns in each NPS, we further applied epicenter mapping to seek out focal “epicenters,” regions whose connectivity patterns—in the healthy brain—most closely mirrored the known disease vulnerability pattern of PD [51]. We found that the epicenters of shared alterations in NPS predominated in SMN regions. The epicenters of apathy, ICBs and hallucinations were also observed in SMN, whereas the epicenters of anxiety and depression were present in DMN regions. Moreover, we observed a MSN-based cluster composed of apathy, ICBs and hallucinations, while anxiety and depression clustered separately. These results indicate different mechanisms of occurrence in different NPS. These data provided evidence that apathy, ICBs and hallucinations may link to the primary pathophysiology of PD in motor circuit, while anxiety and depression are less clear neurobiologic basis, may be the result of a combination of pathology of PD (e.g. related to hypodopaminergic state) and psychological factors.
We found that morphometric similarity was significantly reduced in somatomotor cortex (precentral gyrus, postcentral gyrus), occipital cortex and DLPFC in NPS patients in PD population. Reduced regional MSN implies reduced morphometric similarity (or greater morphometric differentiation) between these areas and the rest of the cortex, which is being possibly attributed to reduced anatomical connectivity to and from the less similar, more differentiated cortical areas [40, 43]. Conversely, MSN increases. This means that these areas are affected by NPS pathology, their structural changes are not as well synchronized with changes elsewhere in the brain network. The epicenter mapping further identified that somatomotor cortex may be the putative source of this damage pattern through functional and structural connectivities. The motor circuit is the major component of basal ganglia-thalamocortical circuits [52], and motor features in PD development are the result of abnormal neuronal activity in the motor circuit [53]. Somatomotor cortex has been found to be the core cortical brain region to be involved in PD in animal and clinical experiments and directly contributes to motor symptoms [52, 54,55,56], and consistent structural [57,58,59,60,61], functional [61,62,63], and metabolicl [64, 65] abnormalities have already been found in neuroimaging studies. PD has long been linked to neuronal networks by the clinical and anatomical progression [53, 66], and the pathological changes in PD typically develop in the nervous system following specific anatomical pattern. The initial α-synuclein misfolding in a small number of cells could progressively lead to the intra-axonal spread of α-synuclein aggregates to multiple brain regions over years or decades following the initial insult [66,67,68,69]. Hence, the current MSN analysis and epicenter mapping provides evidence for the somatomotor cortex being the key sites of onset of the neuropathology of NPS in PD. There is a potential pathologic spreading pathway between somatomotor cortex and structural damage regions.
As a flexible hub for cognitive control, the FPN was considered to play an important role in instantiating and flexibly modulating cognitive control [70]. Similar to DMN, structural changes in the FPN regions are also widely present in many psychiatric disorders such as schizophrenia, bipolar disorder and major depressive disorder [21, 71]. Here, we considered that the increase of MSN in FPN in NPS patients was a compensation for cognitive control disorders, especially in apathy patients. We should note that our cortical-level data are agnostic with regard to specific abnormalities within the “direct” and an “indirect” circuits known to underlie the subcortical pathology of PD [72]. Our evidence of MSN and epicenter abnormalities at network-level of SMN is consistent with the downstream cortical effects of these complex subcortical circuit disruptions.
Specifically exploring various NPS, we found that compared to nNPS, apathy, hallucinations, and ICB exhibited significant alterations primarily in the somatomotor cortex, with MSN alterations involving multiple unimodal networks such as somatomotor, visual, and auditory. These three NPS could be associated with pathophysiology of PD in motor circuit, which has been evidenced by previous epidemiological survey data, as the incidence rates of these NPS in PD were significantly increased compared to normal population [2]. The demographic data in our study also supported the increased incidence rate of apathy and hallucinations in PD. Apathy is a frequent neuropsychiatric disturbance in advanced PD, and its occurrence may be attributed to the structural and functional disruption of different networks in motivational circuits [73]. Hallucinations can occur in early-stage PD and the frequency increases with the progression of PD [74, 75], while the increased risk of ICBs may be related to receiving dopamine therapy, especially the use of dopamine agonists [2, 76]. Epicenter mapping analysis further confirmed our conclusion. Cortical functional and structural epicenters of these three NPS were all located in the somatomotor cortex, indicating that somatomotor cortex alterations in motor circuit may play a crucial role in generating these damage patterns. In contrast, anxiety in PD mainly exhibited decreased MSN in the DLPFC, while depression also showed this trend in spite of no statistically significant difference. Network analysis detected that the MSN alterations were mainly concentrated in DMN. The next epicenter mapping analysis found that the damage patterns of anxiety and depression in PD were related to the DMN. Moreover, brain alterations we observed in anxiety and depression in PD were similar to MSN change in major depressive disorder [40].
We further summarized the above analysis by embedding of each NPS within their co-alterations in MSN space. Quantifying cross-NPS correlations and performing hierarchical clustering analysis further quantified and supported our hypothesis. We speculated that the five NPS in PD had different sources: one mainly manifested as damage to transmodal network and composed of anxiety and depression, and the other was caused by damage to the unimodal network composed of apathy, ICBs and hallucinations. It has been widely confirmed that abnormalities in transmodal regions (i.e., DMN) often occur early and then spread to unimodal regions in common mental disorders [22, 77, 78]. Nonetheless, some mental disorders such as schizophrenia and neuropsychiatric sequelae of stroke may early present abnormalities in unimodal regions [79,80,81,82], and sequentially present a bottom-up spread. These evidences provided that anxiety and depression in PD may be the combination effects of primary pathophysiology of PD(e.g., related to hypodopaminergic state) and social and psychological factors (e.g., low economic status, poor physical health, disability and social isolation) [83]. Considering the similarities with primary psychiatric disorders in terms of altered brain patterns [40, 43, 84], epicenters [22], and high comorbidity [85, 86], it may also be a supporting evidence that social and psychological factors play a partial role in the onset of anxiety and depression in PD. By contrast, apathy, ICBs and hallucinations in PD could belong to another classification due to the similar MSN co-alteration patterns, highly overlapping epicenters and specific abnormalities directionally pointing to SMN, suggesting that they might have a direct link to the primary pathophysiology of PD. In disease management, guided self-help, psychoeducation, advice on exercise and sleep hygiene, cognitive behavioral therapy all show good effects on depression and anxiety in PD [87,88,89], and selective serotonin re-uptake inhibitors are considered first-line treatments [90]. However, antidepressants effective in the management of depression have not been found helpful in the management of apathy in PD [87, 91]. The preliminary evidence indicates that L-dopa, dopaminergic agonists such as pramipexole and ropinirole may improve apathy [92,93,94]. These clinical evidences support our viewpoint to some degree. Our present results provide new insights into the pathogenesis of NPS related to PD.
This study has the following limitations: (i) The sample size (n = 505) is small and larger datasets with neuroimaging data from PD samples are needed. (ii) Our MSN study is limited to the cortical regions and the role of the subcortical region in the MSN damage pattern of PD remains elusive. (iii) We have taken into account the effects of normal aging and sex through the W-score, but the role of chronological age or age at time of PD onset cannot be completely eliminated. (iv) Other factors at work comorbid medical/health problems are not taken into consideration–cerebrovascular/hypertension, diabetes, sleep, and diet/obesity.
Data availability
All data of this study can be found in Parkinson Progression Markers Initiative (PPMI) [30] database (www.ppmi-info.org).
Code availability
For T1-weighted images pre-process, FreeSurfer is available online: http://surfer.nmr.mgh.harvard.edu/. The statistics software of Surfstat is freely available online: https://math.mcgill.ca/keith/surfstat/. Disease epicenter mapping can be found in ENIGMA toolbox [46]: https://enigma-toolbox.readthedocs.io/en/latest/. Other code, and materials are available upon request to the corresponding author Q.Z.
References
Aarsland D, Bronnick K, Alves G, Tysnes OB, Pedersen KF, Ehrt U, et al. The spectrum of neuropsychiatric symptoms in patients with early untreated Parkinson’s disease. J Neurol Neurosurg Psychiatry. 2009;80:928–30.
Weintraub D, Aarsland D, Chaudhuri KR, Dobkin RD, Leentjens AF, Rodriguez-Violante M, et al. The neuropsychiatry of Parkinson’s disease: advances and challenges. Lancet Neurol. 2022;21:89–102.
Gustafsson H, Nordstrom A, Nordstrom P. Depression and subsequent risk of Parkinson disease: a nationwide cohort study. Neurology. 2015;84:2422–9.
Kazmi H, Walker Z, Booij J, Khan F, Shah S, Sudre CH, et al. Late onset depression: dopaminergic deficit and clinical features of prodromal Parkinson’s disease: a cross-sectional study. J Neurol Neurosurg Psychiatry. 2021;92:158–64.
Oosterwijk CS, Vriend C, Berendse HW, van der Werf YD, van den Heuvel OA. Anxiety in Parkinson’s disease is associated with reduced structural covariance of the striatum. J Affect Disord. 2018;240:113–20.
Zhang H, Qiu Y, Luo Y, Xu P, Li Z, Zhu W, et al. The relationship of anxious and depressive symptoms in Parkinson’s disease with voxel-based neuroanatomical and functional connectivity measures. J Affect Disord. 2019;245:580–8.
Cardoso EF, Maia FM, Fregni F, Myczkowski ML, Melo LM, Sato JR, et al. Depression in Parkinson’s disease: convergence from voxel-based morphometry and functional magnetic resonance imaging in the limbic thalamus. Neuroimage. 2009;47:467–72.
Skidmore FM, Yang M, Baxter L, von Deneen K, Collingwood J, He G, et al. Apathy, depression, and motor symptoms have distinct and separable resting activity patterns in idiopathic Parkinson disease. Neuroimage. 2013;81:484–95.
Kostic VS, Agosta F, Petrovic I, Galantucci S, Spica V, Jecmenica-Lukic M, et al. Regional patterns of brain tissue loss associated with depression in Parkinson disease. Neurology. 2010;75:857–63.
Surdhar I, Gee M, Bouchard T, Coupland N, Malykhin N, Camicioli R. Intact limbic-prefrontal connections and reduced amygdala volumes in Parkinson’s disease with mild depressive symptoms. Parkinsonism Relat Disord. 2012;18:809–13.
Reijnders JS, Scholtissen B, Weber WE, Aalten P, Verhey FR, Leentjens AF. Neuroanatomical correlates of apathy in Parkinson’s disease: A magnetic resonance imaging study using voxel-based morphometry. Mov Disord. 2010;25:2318–25.
Carriere N, Besson P, Dujardin K, Duhamel A, Defebvre L, Delmaire C, et al. Apathy in Parkinson’s disease is associated with nucleus accumbens atrophy: a magnetic resonance imaging shape analysis. Mov Disord. 2014;29:897–903.
Baggio HC, Segura B, Garrido-Millan JL, Marti MJ, Compta Y, Valldeoriola F, et al. Resting-state frontostriatal functional connectivity in Parkinson’s disease-related apathy. Mov Disord. 2015;30:671–9.
Biundo R, Weis L, Facchini S, Formento-Dojot P, Vallelunga A, Pilleri M, et al. Patterns of cortical thickness associated with impulse control disorders in Parkinson’s disease. Mov Disord. 2015;30:688–95.
Cerasa A, Salsone M, Nigro S, Chiriaco C, Donzuso G, Bosco D, et al. Cortical volume and folding abnormalities in Parkinson’s disease patients with pathological gambling. Parkinsonism Relat Disord. 2014;20:1209–14.
Pellicano C, Niccolini F, Wu K, O’Sullivan SS, Lawrence AD, Lees AJ, et al. Morphometric changes in the reward system of Parkinson’s disease patients with impulse control disorders. J Neurol. 2015;262:2653–61.
Goldman JG, Stebbins GT, Dinh V, Bernard B, Merkitch D, deToledo-Morrell L, et al. Visuoperceptive region atrophy independent of cognitive status in patients with Parkinson’s disease with hallucinations. Brain. 2014;137:849–59.
Watanabe H, Senda J, Kato S, Ito M, Atsuta N, Hara K, et al. Cortical and subcortical brain atrophy in Parkinson’s disease with visual hallucination. Mov Disord. 2013;28:1732–6.
Pagonabarraga J, Soriano-Mas C, Llebaria G, Lopez-Sola M, Pujol J, Kulisevsky J. Neural correlates of minor hallucinations in non-demented patients with Parkinson’s disease. Parkinsonism Relat Disord. 2014;20:290–6.
Feldmann A, Illes Z, Kosztolanyi P, Illes E, Mike A, Kover F, et al. Morphometric changes of gray matter in Parkinson’s disease with depression: a voxel-based morphometry study. Mov Disord. 2008;23:42–46.
Xie C, Xiang S, Shen C, Peng X, Kang J, Li Y, et al. A shared neural basis underlying psychiatric comorbidity. Nat Med. 2023;29:1232–42.
Hettwer MD, Lariviere S, Park BY, van den Heuvel OA, Schmaal L, Andreassen OA, et al. Coordinated cortical thickness alterations across six neurodevelopmental and psychiatric disorders. Nat Commun. 2022;13:6851.
Seidlitz J, Vasa F, Shinn M, Romero-Garcia R, Whitaker KJ, Vertes PE, et al. Morphometric similarity networks detect microscale cortical organization and predict inter-individual cognitive variation. Neuron. 2018;97:231–47.e237.
Barbas H. General cortical and special prefrontal connections: principles from structure to function. Annu Rev Neurosci. 2015;38:269–89.
Goulas A, Uylings HB, Hilgetag CC. Principles of ipsilateral and contralateral cortico-cortical connectivity in the mouse. Brain Struct Funct. 2017;222:1281–95.
Wei Y, Scholtens LH, Turk E, van den Heuvel MP. Multiscale examination of cytoarchitectonic similarity and human brain connectivity. Netw Neurosci. 2019;3:124–37.
Sterling NW, Lewis MM, Du G, Huang X. Structural imaging and Parkinson’s disease: moving toward quantitative markers of disease progression. J Parkinsons Dis. 2016;6:557–67.
Jucker M, Walker LC. Self-propagation of pathogenic protein aggregates in neurodegenerative diseases. Nature. 2013;501:45–51.
Frost B, Diamond MI. Prion-like mechanisms in neurodegenerative diseases. Nat Rev Neurosci. 2010;11:155–9.
Parkinson Progression Marker I. The Parkinson Progression Marker Initiative (PPMI). Prog Neurobiol. 2011;95:629–35.
Goetz CG, Tilley BC, Shaftman SR, Stebbins GT, Fahn S, Martinez-Martin P, et al. Movement Disorder Society-sponsored revision of the Unified Parkinson’s Disease Rating Scale (MDS-UPDRS): scale presentation and clinimetric testing results. Mov Disord. 2008;23:2129–70.
Spielberger C, Lushene R, Jacobs G. Manual for the State-Trait Anxiety Inven-tory, STAI (Form Y). Palo Alto: Consulting Psychologists Press; 1983.
Yesavage JA, Sheikh JI. 9/Geriatric depression scale (GDS). Clinical Gerontologist. 1986;5:165–73.
Weintraub D, Hoops S, Shea JA, Lyons KE, Pahwa R, Driver-Dunckley ED, et al. Validation of the questionnaire for impulsive-compulsive disorders in Parkinson’s disease. Mov Disord. 2009;24:1461–7.
Dale AM, Fischl B, Sereno MI. Cortical surface-based analysis. I. Segmentation and surface reconstruction. Neuroimage. 1999;9:179–94.
Rosen AFG, Roalf DR, Ruparel K, Blake J, Seelaus K, Villa LP, et al. Quantitative assessment of structural image quality. Neuroimage. 2018;169:407–18.
Glasser MF, Coalson TS, Robinson EC, Hacker CD, Harwell J, Yacoub E, et al. A multi-modal parcellation of human cerebral cortex. Nature. 2016;536:171–8.
Li J, Keller SS, Seidlitz J, Chen H, Li B, Weng Y, et al. Cortical morphometric vulnerability to generalised epilepsy reflects chromosome- and cell type-specific transcriptomic signatures. Neuropathol Appl Neurobiol. 2023;49:e12857.
Seidlitz J, Nadig A, Liu S, Bethlehem RAI, Vertes PE, Morgan SE, et al. Transcriptomic and cellular decoding of regional brain vulnerability to neurogenetic disorders. Nat Commun. 2020;11:3358.
Li J, Seidlitz J, Suckling J, Fan F, Ji GJ, Meng Y, et al. Cortical structural differences in major depressive disorder correlate with cell type-specific transcriptional signatures. Nat Commun. 2021;12:1647.
Orlhac F, Eertink JJ, Cottereau AS, Zijlstra JM, Thieblemont C, Meignan M, et al. A guide to ComBat harmonization of imaging biomarkers in multicenter studies. J Nucl Med. 2022;63:172–9.
Kakimoto A, Ito S, Okada H, Nishizawa S, Minoshima S, Ouchi Y. Age-related sex-specific changes in brain metabolism and morphology. J Nucl Med. 2016;57:221–5.
Morgan SE, Seidlitz J, Whitaker KJ, Romero-Garcia R, Clifton NE, Scarpazza C, et al. Cortical patterning of abnormal morphometric similarity in psychosis is associated with brain expression of schizophrenia-related genes. Proc Natl Acad Sci USA. 2019;116:9604–9.
Ji JL, Spronk M, Kulkarni K, Repovs G, Anticevic A, Cole MW. Mapping the human brain’s cortical-subcortical functional network organization. Neuroimage. 2019;185:35–57.
Lariviere S, Bayrak S, Vos de Wael R, Benkarim O, Herholz P, Rodriguez-Cruces R, et al. BrainStat: a toolbox for brain-wide statistics and multimodal feature associations. Neuroimage. 2023;266:119807.
Lariviere S, Paquola C, Park BY, Royer J, Wang Y, Benkarim O, et al. The ENIGMA Toolbox: multiscale neural contextualization of multisite neuroimaging datasets. Nat Methods. 2021;18:698–700.
Lariviere S, Rodriguez-Cruces R, Royer J, Caligiuri ME, Gambardella A, Concha L, et al. Network-based atrophy modeling in the common epilepsies: a worldwide ENIGMA study. Sci Adv. 2020;6:eabc6457.
Van Essen DC, Ugurbil K, Auerbach E, Barch D, Behrens TE, Bucholz R, et al. The Human Connectome Project: a data acquisition perspective. Neuroimage. 2012;62:2222–31.
Glasser MF, Sotiropoulos SN, Wilson JA, Coalson TS, Fischl B, Andersson JL, et al. The minimal preprocessing pipelines for the Human Connectome Project. Neuroimage. 2013;80:105–24.
Alexander-Bloch AF, Shou H, Liu S, Satterthwaite TD, Glahn DC, Shinohara RT, et al. On testing for spatial correspondence between maps of human brain structure and function. Neuroimage. 2018;178:540–51.
Zhou J, Gennatas ED, Kramer JH, Miller BL, Seeley WW. Predicting regional neurodegeneration from the healthy brain functional connectome. Neuron. 2012;73:1216–27.
Alexander GE, DeLong MR, Strick PL. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci. 1986;9:357–81.
Blesa J, Foffani G, Dehay B, Bezard E, Obeso JA. Motor and non-motor circuit disturbances in early Parkinson disease: which happens first? Nat Rev Neurosci. 2022;23:115–28.
Guo L, Xiong H, Kim JI, Wu YW, Lalchandani RR, Cui Y, et al. Dynamic rewiring of neural circuits in the motor cortex in mouse models of Parkinson’s disease. Nat Neurosci. 2015;18:1299–309.
Pasquereau B, DeLong MR, Turner RS. Primary motor cortex of the parkinsonian monkey: altered encoding of active movement. Brain. 2016;139:127–43.
Boecker H, Ceballos-Baumann A, Bartenstein P, Weindl A, Siebner HR, Fassbender T, et al. Sensory processing in Parkinson’s and Huntington’s disease: investigations with 3D H(2)(15)O-PET. Brain. 1999;122:1651–65.
Tessa C, Lucetti C, Giannelli M, Diciotti S, Poletti M, Danti S, et al. Progression of brain atrophy in the early stages of Parkinson’s disease: a longitudinal tensor-based morphometry study in de novo patients without cognitive impairment. Hum Brain Mapp. 2014;35:3932–44.
Sterling NW, Wang M, Zhang L, Lee EY, Du G, Lewis MM, et al. Stage-dependent loss of cortical gyrification as Parkinson disease “unfolds”. Neurology. 2016;86:1143–51.
Zhang Y, Huang B, Chen Q, Wang L, Zhang L, Nie K, et al. Altered microstructural properties of superficial white matter in patients with Parkinson’s disease. Brain Imaging Behav. 2022;16:476–91.
Taylor KI, Sambataro F, Boess F, Bertolino A, Dukart J. Progressive decline in gray and white matter integrity in de novo Parkinson’s disease: an analysis of longitudinal parkinson progression markers initiative diffusion tensor imaging data. Front Aging Neurosci. 2018;10:318.
Sharman M, Valabregue R, Perlbarg V, Marrakchi-Kacem L, Vidailhet M, Benali H, et al. Parkinson’s disease patients show reduced cortical-subcortical sensorimotor connectivity. Mov Disord. 2013;28:447–54.
Baudrexel S, Witte T, Seifried C, von Wegner F, Beissner F, Klein JC, et al. Resting state fMRI reveals increased subthalamic nucleus-motor cortex connectivity in Parkinson’s disease. Neuroimage. 2011;55:1728–38.
Kurani AS, Seidler RD, Burciu RG, Comella CL, Corcos DM, Okun MS, et al. Subthalamic nucleus-sensorimotor cortex functional connectivity in de novo and moderate Parkinson’s disease. Neurobiol Aging. 2015;36:462–9.
Eckert T, Barnes A, Dhawan V, Frucht S, Gordon MF, Feigin AS, et al. FDG PET in the differential diagnosis of parkinsonian disorders. Neuroimage. 2005;26:912–21.
Sommerauer M, Hansen AK, Parbo P, Fedorova TD, Knudsen K, Frederiksen Y, et al. Decreased noradrenaline transporter density in the motor cortex of Parkinson’s disease patients. Mov Disord. 2018;33:1006–10.
Poewe W, Seppi K, Tanner CM, Halliday GM, Brundin P, Volkmann J, et al. Parkinson disease. Nat Rev Dis Primers. 2017;3:17013.
Angot E, Steiner JA, Hansen C, Li JY, Brundin P. Are synucleinopathies prion-like disorders? Lancet Neurol. 2010;9:1128–38.
Brundin P, Melki R, Kopito R. Prion-like transmission of protein aggregates in neurodegenerative diseases. Nat Rev Mol Cell Biol. 2010;11:301–7.
Braak H, Del Tredici K, Rub U, de Vos RA, Jansen Steur EN, Braak E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol Aging. 2003;24:197–211.
Marek S, Dosenbach NUF. The frontoparietal network: function, electrophysiology, and importance of individual precision mapping. Dialogues Clin Neurosci. 2018;20:133–40.
Matsumoto J, Fukunaga M, Miura K, Nemoto K, Okada N, Hashimoto N, et al. Cerebral cortical structural alteration patterns across four major psychiatric disorders in 5549 individuals. Mol Psychiatry. 2023;28:4915–4923.
McGregor MM, Nelson AB. Circuit mechanisms of Parkinson’s disease. Neuron. 2019;101:1042–56.
Pagonabarraga J, Kulisevsky J, Strafella AP, Krack P. Apathy in Parkinson’s disease: clinical features, neural substrates, diagnosis, and treatment. Lancet Neurol. 2015;14:518–31.
Pagonabarraga J, Martinez-Horta S, Fernandez de Bobadilla R, Perez J, Ribosa-Nogue R, Marin J, et al. Minor hallucinations occur in drug-naive Parkinson’s disease patients, even from the premotor phase. Mov Disord. 2016;31:45–52.
Dlay JK, Duncan GW, Khoo TK, Williams-Gray CH, Breen DP, Barker RA, et al. Progression of neuropsychiatric symptoms over time in an incident Parkinson’s disease cohort (ICICLE-PD). Brain Sci. 2020;10:78.
Weintraub D, Papay K, Siderowf A. Parkinson’s Progression Markers I. Screening for impulse control symptoms in patients with de novo Parkinson disease: a case-control study. Neurology. 2013;80:176–80.
Lui S, Zhou XJ, Sweeney JA, Gong Q. Psychoradiology: the frontier of neuroimaging in psychiatry. Radiology. 2016;281:357–72.
Xu Y, Han S, Wei Y, Zheng R, Cheng J, Zhang Y. Abnormal resting-state effective connectivity in large-scale networks among obsessive-compulsive disorder. Psychol Med. 2024;54:350–358.
Kaufmann T, Skatun KC, Alnaes D, Doan NT, Duff EP, Tonnesen S, et al. Disintegration of sensorimotor brain networks in schizophrenia. Schizophr Bull. 2015;41:1326–35.
Schroder J, Wenz F, Schad LR, Baudendistel K, Knopp MV. Sensorimotor cortex and supplementary motor area changes in schizophrenia. A study with functional magnetic resonance imaging. Br J Psychiatry. 1995;167:197–201.
Ferro JM, Caeiro L, Figueira ML. Neuropsychiatric sequelae of stroke. Nat Rev Neurol. 2016;12:269–80.
Tay J, Morris RG, Markus HS. Apathy after stroke: diagnosis, mechanisms, consequences, and treatment. Int J Stroke. 2021;16:510–8.
Alexopoulos GS. Depression in the elderly. Lancet. 2005;365:1961–70.
Warren SL, Zhang Y, Duberg K, Mistry P, Cai W, Qin S, et al. Anxiety and stress alter decision-making dynamics and causal amygdala-dorsolateral prefrontal cortex circuits during emotion regulation in children. Biol Psychiatry. 2020;88:576–86.
Garber J, Brunwasser SM, Zerr AA, Schwartz KT, Sova K, Weersing VR. Treatment and prevention of depression and anxiety in youth: test of cross-over effects. Depress Anxiety. 2016;33:939–59.
Cummings CM, Caporino NE, Kendall PC. Comorbidity of anxiety and depression in children and adolescents: 20 years after. Psychol Bull. 2014;140:816–45.
Mueller C, Rajkumar AP, Wan YM, Velayudhan L, Ffytche D, Chaudhuri KR, et al. Assessment and management of neuropsychiatric symptoms in Parkinson’s disease. CNS Drugs. 2018;32:621–35.
Pachana NA, Egan SJ, Laidlaw K, Dissanayaka N, Byrne GJ, Brockman S, et al. Clinical issues in the treatment of anxiety and depression in older adults with Parkinson’s disease. Mov Disord. 2013;28:1930–4.
Yang S, Sajatovic M, Walter BL. Psychosocial interventions for depression and anxiety in Parkinson’s disease. J Geriatr Psychiatry Neurol. 2012;25:113–21.
Bomasang-Layno E, Fadlon I, Murray AN, Himelhoch S. Antidepressive treatments for Parkinson’s disease: a systematic review and meta-analysis. Parkinsonism Relat Disord. 2015;21:833–42.
Zahodne LB, Bernal-Pacheco O, Bowers D, Ward H, Oyama G, Limotai N, et al. Are selective serotonin reuptake inhibitors associated with greater apathy in Parkinson’s disease? J Neuropsychiatry Clin Neurosci. 2012;24:326–30.
Czernecki V, Pillon B, Houeto JL, Pochon JB, Levy R, Dubois B. Motivation, reward, and Parkinson’s disease: influence of dopatherapy. Neuropsychologia. 2002;40:2257–67.
Leentjens AF, Koester J, Fruh B, Shephard DT, Barone P, Houben JJ. The effect of pramipexole on mood and motivational symptoms in Parkinson’s disease: a meta-analysis of placebo-controlled studies. Clin Ther. 2009;31:89–98.
Czernecki V, Schupbach M, Yaici S, Levy R, Bardinet E, Yelnik J, et al. Apathy following subthalamic stimulation in Parkinson disease: a dopamine responsive symptom. Mov Disord. 2008;23:964–9.
Acknowledgements
We are grateful to all of the study participants for their patience and cooperation. Data used in the preparation of this article were obtained on February 2023 from the PPMI database (www.ppmi-info.org/access-data-specimens/download-data), RRID:SCR 006431. For up-to-date information on the study, visit www.ppmi-info.org. PPMI – a public-private partnership – is funded by the Michael J. Fox Foundation for Parkinson’s Research and funding partners, including 4D Pharma, Abbvie, AcureX, Allergan, Amathus Therapeutics, Aligning Science Across Parkinson’s, AskBio, Avid Radiopharmaceuticals, BIAL, Biogen, Biohaven, BioLegend, BlueRock Therapeutics, Bristol-Myers Squibb, Calico Labs, Celgene, Cerevel Therapeutics, Coave Therapeutics, DaCapo Brainscience, Denali, Edmond J. Safra Foundation, Eli Lilly, Gain Therapeutics, GE HealthCare, Genentech, GSK, Golub Capital, Handl Therapeutics, Insitro, Janssen Neuroscience, Lundbeck, Merck, Meso Scale Discovery, Mission Therapeutics, Neurocrine Biosciences, Pfizer, Piramal, Prevail Therapeutics, Roche, Sanofi, Servier, Sun Pharma Advanced Research Company, Takeda, Teva, UCB, Vanqua Bio, Verily, Voyager Therapeutics, the Weston Family Foundation and Yumanity Therapeutics.
Funding
This work was supported by the National Natural Science Foundation of China (No. 82271273) and the Jiangsu Social Development Project (BE2022808).
Author information
Authors and Affiliations
Contributions
QL, QZ and KZ concepted and designed research; QL, ZZ, and QZ analyzed and interpreted data; QL, ZZ, HZ, CG, AS, MG, HS, XC, YY acquired data; QL, ZZ, HZ, QZ and KZ drafted the article; QL, ZZ, HZ, JT, QZ and KZ revised the article; QZ and KZ approved the final version to be published. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Lu, Q., Zhu, Z., Zhang, H. et al. Shared and distinct cortical morphometric alterations in five neuropsychiatric symptoms of Parkinson’s disease. Transl Psychiatry 14, 347 (2024). https://doi.org/10.1038/s41398-024-03070-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41398-024-03070-z
