
Abstract
The apparent optimum air temperature for vegetation photosynthesis (Topt) is a key temperature parameter in terrestrial ecosystem models estimating daily photosynthesis or gross primary production (GPP, g C/m2/day). To date, most models use biome-specific Topt (Topt-biome) parameter values. Given vegetation acclimation and adaptation to local climate, site-specific Topt (Topt-site) is needed to reduce uncertainties in estimating daily GPP across the scales from site to region and the globe. Previous studies have demonstrated using the Enhanced Vegetation Index (EVI) derived from Moderate Resolution Imaging Spectroradiometer (MODIS) images and daytime air temperature data to estimate the Topt-site at the eddy covariance tower sites. This study used MODIS-derived EVI and ERA5 climate data to estimate and generate global Topt-site data products from 2000 to 2019. The Topt-site of individual pixels within a biome has large variation, which clearly cannot be represented accurately by the widely used Topt-biome. Therefore, using this global dataset of Topt-site estimates might significantly affect GPP simulation in current ecosystem models.
Similar content being viewed by others

Background & Summary
Among the carbon flux components, Gross Primary Production (GPP) represents the amount of carbon dioxide absorbed by vegetation, playing a pivotal role in the global carbon cycle1,2. Air temperature (Tair, ~2 m above the ground) is one of the key climatic variables influencing the spatiotemporal dynamics of GPP2,3,4. An increase in Tair can boost enzyme activity, leading to a rise in photosynthesis rate, and excessive Tair can result in stomatal closure and enzyme deactivation, causing a decline in photosynthesis5,6. Therefore, the GPP response to Tair follows a parabolic (unimodal) curve4,7, and the apparent optimum air temperature (Topt) is generally defined as the Tair at which vegetation achieves its peak photosynthesis rate.
Almost all terrestrial ecosystem carbon cycle models have temperature response functions for GPP, with Topt being a crucial temperature parameter8,9. Light Use Efficiency (LUE) models also have the Topt parameter in estimating GPP10,11,12. Currently, the Topt parameter values in terrestrial ecosystem models use biome-specific Topt for individual biomes (Topt-biome)13,14. However, recent studies suggest that the Topt-biome may significantly differ from site-specific Topt (Topt-site)15,16. Topt-biome parameter values used in the models came from analyses of the limited number of locations for individual biomes16. Additionally, because of vegetation acclimation and adaption to local climate8,17, the use of biome-specific Topt at the global scale could introduce large errors and uncertainty in global GPP estimation.
GPP and air temperature data from eddy covariance (EC) flux tower sites are widely used to study the response curves of GPP to air temperature and to estimate the Topt-site at individual sites (ecosystems)4,8,16,18,19,20. However, the number of EC towers is limited21, and their footprints only cover a small portion of the global land surface22. Additionally, most EC flux tower observations last a few years or a decade, making it challenging to explore the interannual variations of Topt at the sites. Several studies explored using remote sensing data to estimate Topt-site over large geographical domains and longer periods15,18,23. Since the photosynthesis rate depends on the number of photosynthetic organs (i.e., chloroplasts) in the canopy, it can be directly represented by the total chlorophyll content in the canopy24,25. Many studies have explored the estimation of canopy chlorophyll content from satellite-derived vegetation indices (VIs), such as the Enhanced Vegetation Index (EVI)26,27, Normalized Difference Vegetation Index (NDVI)28, and Near-infrared Reflectance of Vegetation (NIRv)29. These VIs have shown a strong relationship with GPP and are therefore used as predictor variables to represent vegetation structure and function23,25,30,31. Recent studies have demonstrated the feasibility of using VIs-T response curves to estimate Topt-site at site, regional, or global scales7,15,20,23,32. It is worth noting that these publications differ in terms of VIs variables (e.g., NDVI, EVI, and NIRv), air temperature datasets (e.g., daily mean air temperature), and algorithms, leading to noticeable discrepancies in Topt-site estimates across these studies. Accurate quantification of Topt-site and site-year-specific Topt (Topti-site-year) at global scale remains challenging.
In this study, built upon our previous studies that used GPP, EVI, and daytime air temperature data to estimate Topt-site at the eddy covariance tower sites7,20, we first generated the Topt-site-year and Topt-site dataset across the globe from 2000 to 2019, using MODIS-derived EVI data (500-m spatial resolution) and daytime air temperature from ERA5 climate data33,34. We also carried out exploratory data analyses of this dataset, which provides basic information on (1) the spatiotemporal patterns of Topt-site-year and Topt-site across the globe over the past two decades and (2) the variability (frequency distribution) of Topt-site from all the pixels within individual biomes. Terrestrial ecosystem models could use this global Topt-site dataset to increase the accuracy of and reduce the uncertainty of GPP data under historical, current, and future climate scenarios. The Topt-site dataset could also be used for further analyses that offer valuable insights into how vegetation has responded to climate change over the past two decades.
Methods
Datasets and pre-processing
ERA5-Land reanalysis climate data
ERA5 is the fifth generation of atmospheric reanalysis products released by the European Centre for Medium-Range Weather Forecasts (ECMWF)33. This dataset offers comprehensive historical and near-real-time information on global atmospheric, terrestrial, and oceanic variables34. ERA5-Land complements the ERA5 by focusing specifically on terrestrial processes. Compared to ERA5, ERA5-Land boasts a higher spatial resolution (0.1°) and a higher temporal resolution (hourly), enabling a more precise depiction of surface processes and characteristics, including air temperature and precipitation.
In this study, we used the air temperature (Tair) layer from the ERA5-Land reanalysis climate dataset. We provided an example of the air temperature data from the Harvard Forest site (US-Ha1) in 2010 (Fig. 2) and illustrate the discrepancies among the three temperature variables: daily maximum air temperature (Tair-max), daily daytime mean air temperature (Tair-DT), and daily mean air temperature (Tair-mean); and we assessed the appropriateness of these three temperature variables for the Topt estimation algorithm. Tair-DT is defined by averaging Tair-max with Tair-mean10:
We aggregated these three air temperature variables from daily data into 8-day intervals by averaging the observations over the 8-day period to align with the temporal resolution of MOD09A1.
MODIS land surface temperature dataset
The Moderate Resolution Imaging Spectroradiometer (MODIS) is a key instrument for observing changes on the Earth’s surface35. The MYD11A2 dataset provides an 8-day and 1-km composite of global land surface temperatures (LST) and emissivity, sourced from the MODIS instrument aboard NASA’s Aqua satellite. This dataset provides LST at 1:30 a.m. and 1:30 p.m. In this study, we constrained the analyses of EVI and Tair data and Topt algorithm within the thermal growing season36, as defined by nighttime LST37, which helps to mitigate uncertainties from outliers beyond this period (e.g., high EVI caused by snow). For each pixel (1-km spatial resolution), we reconstructed the LST time series using a moving window of three consecutive 8-day periods, calculating the average across them (Fig. 1a). Adopting the threshold assumption that the LST for the thermal growing season is > = 5 °C38, the start of the season (SOSLST) was marked by the LST first surpassing this threshold in winter/spring, while its end of the season (EOSLST) corresponds to the LST first descending below this threshold in fall/winter (Fig. 1a). Note this method was applied only to those pixels where the annual peak LST surpasses 5 °C. For areas with an annual peak LST not reaching 5 °C (e.g., high-altitude plateau or boreal forests), we substituted LST with Tair-DT.

The workflow for estimating site-specific apparent optimum temperature (Topt-site) for vegetation photosynthesis at a deciduous broadleaf forest site in the USA (Harvard Forest, US-Ha1, DBF, 42.5378°N, 72.1715°W). The US-Ha1 site was selected because it is a mature forest subject to long-term observation and research, showing the typical seasonal dynamics of vegetation in mid-latitude areas. The altitude is 340 meters. In contrast, we also selected the Amazon forest site BR-Sa3 to show the seasonal dynamics of vegetation in tropical regions (Fig. S1). (a) raw LST (LSTraw) and reconstructed LST (LSTre) time series. The thermal growing season is defined as the period LSTre greater than 5 °C (light blue filled area). SOSLST and EOSLST represent the start and end of the thermal growing season, respectively; (b) raw EVI (EVIraw, only after cloud removal), gap-filled EVI (EVIgapfill, linear interpolation), and smoothed EVI (EVIsmooth, S-G filter); (c) The curve of EVI-Tair. The solid green curve is fitting curves using cubic regression splines. (c) also shows a boxplot with points every 2 °C bin; (d) interannual variations of Topt-site-year from 2000 to 2019, Topt-site is calculated as the median value from 20 years of Topt-site-year data.
MODIS surface reflectance and vegetation indices
The MODIS surface reflectance products are widely used in monitoring global vegetation39. In this study, we utilized the MOD09A1 Surface Reflectance V6.1 product, a dataset of seven spectral bands ranging from 0.459μm to 2.155μm with a 500-m spatial and an 8-day temporal resolution40.
We have processed and analyzed the global MOD09A1 surface reflectance time series data from 2000 to 2019. Three vegetation indices - NDVI, EVI, and NIRv - were calculated. NDVI, utilizing near-infrared and red bands, offers a measure of vegetation greenness and is hence used to identify green vegetation areas41. EVI, while similar to NDVI, is specifically designed to correct for atmospheric disturbances and background soil effects42. NIRv is derived from NDVI, yet it exhibits a stronger correlation with observed GPP than NDVI43. Among these vegetation indices, EVI has been reported to be most closely related to leaf and canopy-level chlorophyll content26,27,28,31,44. Additionally, several biophysical studies have shown that EVI is nearly unsaturated and sensitive to canopy changes26,31. The three VIs are calculated by the formulas (2–4):
where ρnir, ρred, and ρblue are the surface reflectance bands of near infrared, red, and blue, respectively. In order to generate a high-quality and continuous surface reflectance time series, the QA-band quality detection approach was employed to retrieve pixels affected by clouds and cloud shadows. Additionally, the Best Index Slope Extraction (BISE) method was used to detect overlooked low-quality observation pixels45. After two rounds of filtration processes, the low-quality observations were then marked as a mask for the row image (Fig. 1b). A linear interpolation approach, coupled with high-quality observations, was used to fill the gaps46,47 (Fig. 1b). Finally, to further minimize the temporal variation within the time series data, we utilized the Savitzky-Golay (S-G) method for curve smoothing48. Note that the smoothing window is set based on the varying lengths of the thermal growing season; for instance, if the thermal growing season of a pixel is 240 days, a sliding window of 31 8-day observations was employed for curve smoothing.
MODIS land cover product
The MODIS Land Cover Type (MCD12Q1) Version 6.1 data product offers annual global land cover classifications from 2001 to the present49. In this study, we utilized the International Geosphere Biosphere Program (IGBP) layer, which identifies 17 distinct land cover types. Our focus was on the following 14 categories: Evergreen Needleleaf Forests (ENF), Evergreen Broadleaf Forests (EBF), Deciduous Needleleaf Forests (DNF), Deciduous Broadleaf Forests (DBF), Mixed Forests (MF), Closed Shrublands (CSH), Open Shrublands (OSH), Woody Savannas (WSA), Savannas (SAV), Grasslands (GRA), Permanent Wetlands (WET), Croplands (CRO), Urban and Built-up Lands (URB), and Cropland/Natural Vegetation Mosaics (CNV). The spatial delineation of these categories is illustrated in Fig. S2. In this paper, we explored the variation of Topt-site within individual biomes for the above-mentioned 14 biomes.
Methods to estimate Topt-site-year and Topt-site
Air temperature variables input
We investigated the seasonal dynamics of Tair-max, Tair-DT, and Tair-mean within one year (Fig. 2a, the case at Harvard Forest site). The discrepancy between Tair-max and Tair-DT (or between Tair-DT and Tair-mean) can be as high as 3 °C throughout the whole year.

Temporal dynamics of air temperature and GPPEC at the Harvard Forest eddy flux tower site (US-Ha1) in 2010. (a) Seasonal dynamics of daily maximum air temperature (Tair-max), daily daytime mean air temperature (Tair-DT), and daily mean air temperature (Tair-mean). This figure also shows the difference between Tair-max and Tair-DT (Tdiff); (b) diurnal dynamics of GPP and Tair; (c) 2-d scatter plot of GPP and Tair.
A further examination of the diurnal dynamics of GPP and Tair is presented in Fig. 2b. Interestingly, the diurnal peaks of GPP and Tair were not consistent. GPP reached its peak between 9:00 a.m. and 11:00 a.m., whereas Tair peaked later, from 2:00 p.m. to 4:00 p.m. Of particular note is that among the three temperature variables, Tair-DT most closely approximated the temperature of Tair at the moment GPP peaked (Fig. 2c). Given these observations, Tair-DT is regarded as the most appropriate air temperature variable for the algorithm input.
Algorithm of Topt-site-year and Topt-site
Topt-site-year is derived from the annual response curve of EVI to Tair-DT16,20. For each year, we plotted the response curve of EVI versus Tair-DT with all observations within the thermal growing season (Fig. 1c). Then, the curve fitting of EVI against Tair-DT was performed using cubic regression splines. The optimum Tair-DT range for vegetation photosynthesis is defined as when EVI exceeds 95% of its maximum value, and Topt-site-year is determined from the average of all Tair-DT within that range (Fig. 1c). Topt-site was calculated as the median value of Topt-site-year across all 20 years (Fig. 1d).
Data Records
The datasets include Topt-site-year with 500-meter spatial resolution across the globe from 2000 to 2019, labeled as “Global_Topt_site_year_“ followed by the specific year, and Topt-site, labeled as “Global_Topt_site”. The units are in °C with a scalar factor of 0.01. The maps only show the areas where the annual mean NDVI is greater than 0.1 (vegetated pixel). The datasets have been submitted to the figshare data repository portal: https://doi.org/10.6084/m9.figshare.24514459.v650.
Fig. 3a shows the global distribution of Topt-site-year for the year 2000. From a global perspective, 91% of the vegetation has their Topt-site-year within the 10 to 30 °C range, while 3% and 6% have their Topt-site-year below 10 °C and above 30 °C, respectively (Fig. 3a). Across the latitudinal gradient, Topt-site-year ranges between 4 to 35 °C (Fig. 3b). The highest Topt-site-year is observed in the tropics. The Topt-site-year value declines with a northward or southward progression in latitude, decreasing by an average of 0.34 °C for each 1° higher latitude, reaching its minimum near the poles. In high-altitude regions, such as the Qinghai-Tibet Plateau and Andes Mountains, the Topt-site-year is significantly low.

Global maps of apparent optimum air temperature and interannual variation. (a) global map of Topt-site-year in 2000; (c) global map of Topt-site, derived from the median value of Topt-site-year from 2000 to 2019; (e) Topt-std is calculated as the standard deviation of Topt-site-year during the period. The global map only shows the area where the annual NDVI is larger than 0.1 (The blank areas are pixels where the annual NDVI is less than 0.1). The histogram represents the proportion of pixels within the different intervals, as shown on the x-axis. The red line represents the Kernel Density Estimation (KDE). (b,c,f) correspond to the average values across the latitude gradient in (a,c,e), respectively. The solid black lines are calculated as the average value for all pixels within 1° of latitude, and the shadow indicates the standard deviation.
Fig. 3c shows the global map of Topt-site, derived from the median value of two decades of Topt-site-year. The spatial distribution and its trend across the latitude gradient mirror the patterns observed for Topt-site-year in 2000 (Fig. 3c,d). Comparing the histograms in Fig. 3c and Fig. 3a, the Topt-site shows a slight increase relative to the Topt-site-year from 2000. Notably, between 25 °C and 30 °C, the proportion of vegetation with this Topt-site has marginally increased (~approximately 5% pixels higher). We further investigate the trends at five-year intervals (Fig. S3). The Topt-site-year exhibited a slight global increasing trend, predominantly observed in arid regions such as central-northern Australia, North Africa, and southern North America. In these areas, the Topt-site-year showed an increase exceeding 5 °C.
Fig. 3e presents the standard deviation of the global Topt-site-year (Topt-std) over the last two decades. Between 2000 and 2019, the histogram shows that Topt-std primarily fluctuated between 1 °C and 3 °C for 71% of the vegetation pixels in the globe. Meanwhile, 14% of the vegetation exhibited relative stability with interannual variations of less than 1 °C, and 15% of vegetation experienced fluctuations greater than 3 °C. Across the latitude gradient (Fig. 3f), the largest Topt-std predominantly occurs in the northern boreal and southern temperate zones. Within the northern boreal zone, the Topt-std increases with latitude, up to 3 °C. In the southern temperate zone, Australia manifests the largest Topt-std, up to 5 °C. Tropical regions exhibit the smallest Topt-std with an interannual variability of around 1 °C, followed by the northern temperate zone, which shows a moderate fluctuation ranging from 2 to 3 °C.
The histogram in Fig. 4 shows the range (frequency distribution) of Topt-site across 14 biomes, revealing significant variation of Topt-site due to geographical and climatic differences. For instance, the GRA exhibits a wide Topt-site range from 4 °C to 35 °C, reflecting their extensive global distribution. In contrast, the DNF, primarily found in colder boreal zones, has a narrower range of 14 °C to 21 °C.

Technical Validation
Comparison against Topt-site derived from eddy covariance flux tower (\({{\rm{T}}}_{{\rm{opt}}-{\rm{site}}}^{{\rm{GPP}}}\))
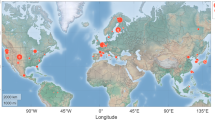
We compared the Topt-site (derived from EVI) with the \({{\rm{T}}}_{\mathrm{opt} \mbox{-} \mathrm{site}}^{\mathrm{GPP}}\) (derived from GPP, see Supplementary methods 1) at 137 eddy flux tower sites across 11 out of 14 biomes (DNF, URB, and CNV were excluded due to their very limited number of sites) in the globe (Fig. 5a). We employed a simple linear regression analysis (see Supplementary methods 2) to investigate the relationship between Topt-site and \({T}_{\mathrm{opt} \mbox{-} \mathrm{site}}^{\mathrm{GPP}}\). The Topt-site generated in this study showed high consistency with \({T}_{\mathrm{opt} \mbox{-} \mathrm{site}}^{\mathrm{GPP}}\) across all 11 biomes (r2 between 0.80 and 0.95) (Fig. 5b). The black solid line represents the overall linear fit for all biomes, exhibiting an r² of 0.83 (By comparison, the number is 0.52 in Huang et al.15 across 125 sites), indicating that the Topt-site estimated in this study is reliable at the ecosystem scale.

The comparison with Topt-site and \({T}_{\mathrm{opt} \mbox{-} \mathrm{site}}^{\mathrm{GPP}}\) across the globe. (a) The 137 eddy flux tower sites across the 11 biomes in the globe; (b) Relationship between Topt-site and \({{\rm{T}}}_{{\rm{opt}} \mbox{-} {\rm{site}}}^{{\rm{GPP}}}\). Error bars indicate ± S.D. from Topt-site-year and \({{\rm{T}}}_{{\rm{opt}} \mbox{-} {\rm{site}} \mbox{-} {\rm{year}}}^{{\rm{GPP}}}\), respectively. Different colors indicate different biomes, and the black solid line indicates all biomes.
Comparison of materials and methods used to estimate Topt-site
We compiled previous studies that utilized GPP/NEE/VIs-Tair response curves for estimating Topt-site (Table 1). These studies varied in terms of VIs variables, air temperature variables, and algorithms, leading to noticeable discrepancies in global Topt-site estimations. For instance, two studies estimated the mean global Topt at 18.8 ± 7.1 °C18 and 23 ± 7.8 °C15, respectively. In contrast, this study estimates it to be 22 ± 6 °C. The discrepancies highlight the necessity and challenges associated with reducing uncertainties in variables, data sources, and algorithms.
In terms of vegetation index variables used in estimating Topt-site, NDVI, as one of the earliest and most commonly used vegetation indices51,52, might introduce biases when estimating Topt-site based on the NDVI- temperature response curve. This bias is attributed to the characteristics of NDVI dynamics in the plant growing season. In those sites of dense forest with high leaf area index (> = 4 m2/m2), NDVI may saturate and remain high throughout the summer, which leads to a shift in the Topt window from early summer to mid-summer, causing an overestimation of Topt (Fig. S4). In the later summer, although NDVI remains high, GPP has already begun to decline23, The histograms of NDVI-derived Topt-site map show that pixels with Topt-site over 20 °C make up 64% (Fig. S5a), whereas this proportion is 55% in EVI-derived Topt-site map (Fig. 3b). Recent studies employed NIRv to generate regional and global Topt-site maps15,32. Given that EVI exhibits a higher correlation with GPP than NDVI in most ecosystems like forests26,27, crops53,54, and grasslands55, it can be regarded as an ideal proxy to study vegetation response to temperature variations. Our findings also show that the Topt-site map derived from NIRv has similar global patterns and magnitudes with those derived from EVI (Fig. S5c). This consistency can be attributed to the close correlation of EVI and NIRv, and both of them also have strong correlation with solar-induced chlorophyll fluorescence (SIF), which is the amount of light emitted by plants15,43.
In terms of air temperature variables used in estimating Topt-site, most studies employ two types of air temperature variables: Tair-max15,32 and Tair-mean8,18,19. However, the peak of vegetation photosynthesis does not occur at the hottest moment of the day (as shown in Fig. 2). The maximum GPP typically occurs in the later morning, while Tair-max tends to occur in the earlier afternoon. This renders Tair-max an inappropriate choice for air temperature variables in the analyses of air temperature and GPP relationship, as it tends to overestimate Topt-site at the higher end, i.e., a bias of higher Topt-site values. Since photosynthesis only happens during daytime and Tair-mean factors in nighttime temperatures, the use of Tair-mean will introduce a bias of lower Topt-site values. Previous studies have already demonstrated the potential and feasibility of estimating Topt based on the EVI-Tair-DT response curve at a limited number of sites7,20, and to our knowledge, this study is the first time that Tair-DT was used as variable inputs to estimate global Topt-site.
In terms of algorithms, most studies defined Topt as the air temperature corresponding to the highest point of the VI-Tair or GPP-Tair response curve with multi-year data4,8,15,18,19,32; for example, the recent global Topt-site estimates by Huang ey al.15. Different with those studies, this study first calculated annual Topt-site-year and then estimated Topt-site as the median of Topt-site-year in 20 years. The advantage of this method is that Topt-site-year provides the variation information of Topt-site across years, especially considering the substantial inter-annual variations in climate and land cover in the past two decades in the globe. Topt-site-year data would help expand the understanding of how vegetation responds to environmental changes over time. In addition, abnormal vegetation growth caused by extreme climate events in specific years may also introduce uncertainty to the estimation. We use the median value of Topt-site-year over multiple years, as the median value is primarily used to eliminate potential outliers and will provide a more balanced perspective of the Topt-site over time. Additionally, we improved the Topt-site estimation window based on the thermal growing season defined by LST, which significantly reduced the uncertainty introduced by snow in the algorithm, particularly in high-latitude regions.
Spatial variation of Topt-site across biomes from this study and its comparison with Topt-biome
The range of Topt-site highlights the limitations of using a fixed Topt-biome parameter. Topt-biome shows a substantial discrepancy when compared with the Topt-site across biomes. This discrepancy is especially pronounced in such biomes as ENF, OSH, SAV, and CRO, where the Topt-site is always lower than the Topt-biome. This bias is mainly due to the Topt-biome being based on limited site data, further introducing significant uncertainty into the GPP estimation7,20,56,57. Our previous research suggests that replacing the Topt-biome with Topt-site can significantly improve the accuracy of GPP estimates from the model at site scale by approximately 1% to 34%, especially in the GRA, CRO, and OSH ecosystems16. This highlights the importance of using the Topt-site rather than the Topt-biome for simulations of GPP models across the scales from site to region and the globe.
Usage Notes
The Topt-site dataset produced in this study can be used as an input for most ecosystem models that include temperature parameters. Note that we have masked areas with an annual average NDVI of less than 0.1 (such as deserts, glaciers, or areas with high-density impervious surfaces). Furthermore, Topt-site is highly related to plant physiology, ecology, and climatology. The Topt-site-year dataset could be used to explore the significant importance of understanding plant growth’s environmental dependence, ecosystems’ functioning, and their responses to environmental changes.
Code availability
The algorithm is run on Google Earth Engine (GEE) and the code can be found at this link: https://doi.org/10.6084/m9.figshare.2458725958.
References
Frankenberg, C. et al. New global observations of the terrestrial carbon cycle from GOSAT: Patterns of plant fluorescence with gross primary productivity. Geophysical Research Letters. 38 (2011).
Anav, A. et al. Spatiotemporal patterns of terrestrial gross primary production: A review. Reviews of Geophysics. 53, 785–818 (2015).
Jung, M. et al. Global patterns of land‐atmosphere fluxes of carbon dioxide, latent heat, and sensible heat derived from eddy covariance, satellite, and meteorological observations. Journal of Geophysical Research: Biogeosciences. 116 (2011).
Bennett, A. C. et al. Thermal optima of gross primary productivity are closely aligned with mean air temperatures across Australian wooded ecosystems. Global Change Biology. 27, 4727–4744 (2021).
Lloyd, J. & Farquhar, G. D. Effects of rising temperatures and [CO2] on the physiology of tropical forest trees. Philosophical Transactions of the Royal Society B: Biological Sciences. 363, 1811–1817 (2008).
Medlyn, B. et al. Temperature response of parameters of a biochemically based model of photosynthesis. II. A review of experimental data. Plant, Cell & Environment. 25, 1167–1179 (2002).
Chang, Q. et al. Assessing variability of optimum air temperature for photosynthesis across site-years, sites and biomes and their effects on photosynthesis estimation. Agricultural and Forest Meteorology. 298, 108277%@ 100168–101923 (2021).
Niu, S. et al. Thermal optimumity of net ecosystem exchange of carbon dioxide and underlying mechanisms. New Phytologist. 194, 775–783 (2012).
Friedlingstein, P. et al. Climate–carbon cycle feedback analysis: results from the C4MIP model intercomparison. Journal of climate. 19, 3337–3353 (2006).
Zhang, Y. et al. A global moderate resolution dataset of gross primary production of vegetation for 2000–2016. Scientific data. 4, 1–13 (2017).
Running, S. W. & Zhao, M. Daily GPP and annual NPP (MOD17A2/A3) products NASA Earth Observing System MODIS land algorithm. MOD17 User’s Guide. 2015, 1–28 (2015).
Yuan, W. et al. Deriving a light use efficiency model from eddy covariance flux data for predicting daily gross primary production across biomes. Agricultural and Forest Meteorology. 143, 189–207 (2007).
Raich, J. et al. Potential net primary productivity in South America: application of a global model. Ecological applications. 1, 399–429 (1991).
Melillo, J. M. et al. Global climate change and terrestrial net primary production. Nature. 363, 234–240 (1993).
Huang, M. et al. Air temperature optima of vegetation productivity across global biomes. Nature ecology & evolution. 3, 772–779 (2019).
Chang, Q. et al. Assessing variability of optimum air temperature for photosynthesis across site-years, sites and biomes and their effects on photosynthesis estimation. Agricultural and Forest Meteorology. 298, 108277 (2021).
Mooney, H. A., Björkman, O. & Collatz, G. J. Photosynthetic acclimation to temperature in the desert shrub, Larrea divaricata: I. Carbon dioxide exchange characteristics of intact leaves. Plant Physiology. 61, 406–410 (1978).
Wang, B. et al. Dryness limits vegetation pace to cope with temperature change in warm regions. Global Change Biology. 29, 4750–4757 (2023).
Yuan, W. et al. Thermal adaptation of net ecosystem exchange. Biogeosciences. 8, 1453–1463 (2011).
Chang, Q. et al. Estimating site-specific optimum air temperature and assessing its effect on the photosynthesis of grasslands in mid-to high-latitudes. Environmental Research Letters. 15, 034064 (2020).
Baldocchi, D. Measuring fluxes of trace gases and energy between ecosystems and the atmosphere–the state and future of the eddy covariance method. Global change biology. 20, 3600–3609 (2014).
Papale, D. et al. Towards a standardized processing of Net Ecosystem Exchange measured with eddy covariance technique: algorithms and uncertainty estimation. Biogeosciences. 3, 571–583 (2006).
Chen, Y., Feng, X., Fu, B., Wu, X. & Gao, Z. Improved global maps of the optimum growth temperature, maximum light use efficiency, and gross primary production for vegetation. Journal of Geophysical Research: Biogeosciences. 126, e2020JG005651 (2021).
Schlemmer, M. et al. Remote estimation of nitrogen and chlorophyll contents in maize at leaf and canopy levels. International journal of applied earth observation and geoinformation. 25, 47–54 (2013).
Peng, Y., Gitelson, A. A. & Sakamoto, T. Remote estimation of gross primary productivity in crops using MODIS 250 m data. Remote Sensing of Environment. 128, 186–196 (2013).
Xiao, X. et al. Satellite-based modeling of gross primary production in an evergreen needleleaf forest. Remote sensing of environment. 89, 519–534 (2004).
Xiao, X. et al. Modeling gross primary production of temperate deciduous broadleaf forest using satellite images and climate data. Remote sensing of environment. 91, 256–270 (2004).
Xin, F. et al. Modeling gross primary production of paddy rice cropland through analyses of data from CO2 eddy flux tower sites and MODIS images. Remote sensing of environment. 190, 42–55 (2017).
Wu, G. et al. Radiance-based NIRv as a proxy for GPP of corn and soybean. Environmental Research Letters. 15, 034009 (2020).
Hunt, E. R. Jr et al. A visible band index for remote sensing leaf chlorophyll content at the canopy scale. International journal of applied earth observation and Geoinformation. 21, 103–112 (2013).
Gao, Y. et al. A MODIS-based Photosynthetic Capacity Model to estimate gross primary production in Northern China and the Tibetan Plateau. Remote Sensing of Environment. 148, 108–118 (2014).
Chen, A., Huang, L., Liu, Q. & Piao, S. Optimum temperature of vegetation productivity and its linkage with climate and elevation on the Tibetan Plateau. Global Change Biology. 27, 1942–1951 (2021).
Hersbach, H. et al. The ERA5 global reanalysis. Quarterly Journal of the Royal Meteorological Society. 146, 1999–2049 (2020).
Muñoz-Sabater, J. et al. ERA5-Land: A state-of-the-art global reanalysis dataset for land applications. Earth System Science Data. 13, 4349–4383 (2021).
Justice, C. O. et al. An overview of MODIS Land data processing and product status. Remote sensing of Environment. 83, 0034–4257 (2002).
Linderholm, H. W., Walther, A. & Chen, D. Twentieth-century trends in the thermal growing season in the Greater Baltic Area. Climatic Change. 87, 405–419 (2008).
Zhang, Y. et al. Consistency between sun-induced chlorophyll fluorescence and gross primary production of vegetation in North America. Remote Sensing of Environment. 183, 154–169 (2016).
Dong, J. et al. Tracking the dynamics of paddy rice planting area in 1986–2010 through time series Landsat images and phenology-based algorithms. Remote Sensing of Environment. 160, 99–113 (2015).
Justice, C. O. et al. The Moderate Resolution Imaging Spectroradiometer (MODIS): Land remote sensing for global change research. IEEE transactions on geoscience and remote sensing. 36, 1228–1249 (1998).
Didan, K., Munoz, A. B., Solano, R. & Huete, A. MODIS vegetation index user’s guide (MOD13 series). University of Arizona: Vegetation Index and Phenology Lab. (2015).
Rouse, J. W., Haas, R. H., Schell, J. A. & Deering, D. W. Monitoring vegetation systems in the Great Plains with ERTS. NASA Spec. Publ. 351, 309 (1974).
Huete, A. et al. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote sensing of environment. 83, 195–213 (2002).
Badgley, G., Field, C. B. & Berry, J. A. Canopy near-infrared reflectance and terrestrial photosynthesis. Science advances. 3, e1602244 (2017).
Pan, L. et al. Interannual variations and trends of gross primary production and transpiration of four mature deciduous broadleaf forest sites during 2000–2020. Remote Sensing of Environment. 304, 114042 (2024).
Viovy, N., Arino, O. & Belward, A. The Best Index Slope Extraction (BISE): A method for reducing noise in NDVI time-series. International Journal of remote sensing. 13, 1585–1590 (1992).
Liu, L. et al. Mapping cropping intensity in China using time series Landsat and Sentinel-2 images and Google Earth Engine. Remote Sensing of Environment. 239, 111624 (2020).
Wang, J. et al. Mapping sugarcane plantation dynamics in Guangxi, China, by time series Sentinel-1, Sentinel-2 and Landsat images. Remote sensing of environment. 247, 111951 (2020).
Savitzky, A. & Golay, M. J. Smoothing and differentiation of data by simplified least squares procedures. Analytical chemistry. 36, 1627–1639 (1964).
Sulla-Menashe, D. & Friedl, M. A. User guide to collection 6 MODIS land cover (MCD12Q1 and MCD12C1) product. Usgs: Reston, Va, Usa. 1, 18 (2018).
Pan, L. & Xiao, X. Site-specfic optimum air temperature for vegetation photosynthesis across the globe from 2000 to 2019. Figshare. https://doi.org/10.6084/m9.figshare.24514459.v6 (2024).
Potter, C. S. et al. Terrestrial ecosystem production: a process model based on global satellite and surface data. Global biogeochemical cycles. 7, 811–841 (1993).
Cui, Y. Preliminary estimation of the realistic optimum temperature for vegetation growth in China. Environmental management. 52, 151–162 (2013).
Wagle, P., Xiao, X. & Suyker, A. E. Estimation and analysis of gross primary production of soybean under various management practices and drought conditions. ISPRS Journal of Photogrammetry and Remote Sensing. 99, 70–83 (2015).
Jin, C. et al. Effects of in-situ and reanalysis climate data on estimation of cropland gross primary production using the Vegetation Photosynthesis Model. Agricultural and Forest Meteorology. 213, 240–250 (2015).
Li, Z. et al. Modeling gross primary production of alpine ecosystems in the Tibetan Plateau using MODIS images and climate data. Remote Sensing of Environment. 107, 510–519 (2007).
Heinsch, F. A. et al. Evaluation of remote sensing based terrestrial productivity from MODIS using regional tower eddy flux network observations. IEEE transactions on geoscience and remote sensing. 44, 1908–1925 (2006).
Yan, H. et al. Improved global simulations of gross primary product based on a new definition of water stress factor and a separate treatment of C3 and C4 plants. Ecological Modelling. 297, 42–59 (2015).
Pan, L. & Xiao, X. Source code for: Site-specific optimum air temperature for vegetation photosynthesis across the globe. Figshare. https://doi.org/10.6084/m9.figshare.24587259 (2024).
Acknowledgements
This study was supported in part by research grants from the U.S. National Science Foundation (OIA-1946093). This study was also supported by the NASA Earth Science Research from Operational Geostationary Satellite Systems (ESROGSS, 80NSSC24K0118). We thank the reviewers for their comments and suggestions on this manuscript.
Author information
Authors and Affiliations
Contributions
X.X. and L.P. designed the study. L.P. and X.X. analyzed the data, interpreted the results, and drafted the manuscript. All authors reviewed and edited the manuscript and figures.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
41597_2024_3603_MOESM1_ESM.docx
Supplementary Online materials for “Site-specific apparent optimum air temperature for vegetation photosynthesis across the globe”
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pan, L., Xiao, X., Yao, Y. et al. Site-specific apparent optimum air temperature for vegetation photosynthesis across the globe. Sci Data 11, 758 (2024). https://doi.org/10.1038/s41597-024-03603-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41597-024-03603-7
