
Abstract
Xiaozhou mustard (Brassica napiformis) root tuber, a traditional fermented vegetable, has a long history in Rongan County, Guangxi Province. However, the frequent occurrence of root tuber sour rot by Geotrichum candidum (G. candidum) has seriously reduced Xiaozhou mustard production and quality in recent years. The objective of the present study is to investigate the antifungal efficacy of 2-chloro-5-trifluoromethoxybenzeneboronic acid (Cl-F-BBA) against G. candidum and its possible mechanisms. The results revealed that a concentration of 0.25 mg/mL Cl-F-BBA completely halted mycelial growth and spore germination. Furthermore, a slightly lower concentration of 0.20 mg/mL was sufficient to compromise the integrity of the plasma membrane in mycelia and mitochondria, leading to a reduction in respiratory rate, activities of malate dehydrogenase (MDH), and succinate dehydrogenase (SDH), ATP content, and energy charge. This concentration also significantly disordered antioxidant metabolism, resulting in the accumulation of reactive oxygen species (ROS) and malondialdehyde (MDA), and caused intracellular leakage in mycelia. In vivo experiments further demonstrated that Xiaozhou mustard root tubers treated with Cl-F-BBA exhibited markedly lower decay rates and lesion diameters compared to the control group. In summary, Cl-F-BBA presents a promising solution for controlling root tuber sour rot in Xiaozhou mustard caused by G. candidum.
Similar content being viewed by others


Fruits and vegetables undergo rapid spoilage as a result of infection with many fungal pathogens, which cause decay before and after harvest and reduce the quality1,2. Xiaozhou mustard root tuber (Brassica napiformis), a traditional fermented vegetable, has a long history in Rongan County, Guangxi Province, China. Xiaozhou mustard is usually planted in autumn and harvested in next spring, and root tubers are susceptible to pathogen infection causing soft and decay. Many synthetic fungicides were applied to control fruit and vegetable rot3. Nevertheless, their side effects on human health and environmental safety should not be ignored4. Therefore, many fungicides have been excluded from the market, which raises a growing interest about the alternatives to traditionally synthesized fungicides5. A growing number of scientists are interested in boronic acids (BA) because boron is an essential micronutient element required to plants, and BA is considered to be safe and environmentally friendly in application if used sensibly. Several studies have demonstrated that BA has a wide range of antibacterial and antifungal properties and has been confirmed to be effective against several species of human-pathogenic fungi6. Moreover, several studies have demonstrated that BA can control postharvest diseases caused by Penicillium digitatum7 and grapevine fruit disease caused by Botrytis cinerea8. Studies have also shown that BA also reduces the development of Monilinia laxa on peaches9. Recently, the antifungal activity of boron-containing small molecule compounds, e.g. phenylboronic acid (PBA), has attracted much attention of researchers because of their remarkable antifungal effect with low concentration as compared with BA, as well as their abundant biological activities, and environmently friendly features10,11,12. For instance, data from Martinko et al.12 demonstrated that PBA was more efficient in suppressing the impact of Alternaria alternata infection than BA on tomato plants. Furthermore, it was found that boron-containing small molecule compounds had high efficiency and broad-spectrum antifungal activities against plant pathogens13. For example, 2-chloro-5-trifluoromethoxybenzeneboronic acid (Cl-F-BBA) showed better activity against Botrytis cinerea than fungicide boscalid with low cytotoxicity to mammal cells13. However, there have been no reports on the application of Cl-F-BBA in controlling the fungal disease of Xiaozhou mustard root tuber sour rot. Herein, the objective of our research are to investigate the efficacy and mechanisms of Cl-F-BBA against fungal, and to provide a new agent for controlling Xiaozhou mustard root tuber sour rot.
Results
Isolation and identification of fungi
Xiaozhou mustard root tubers rotted when infected by the pathogen before harvest (Fig. 1A) and during drying after harvest (Fig. 1B). The common symptoms of the diseased root tuber included a brown, watery, and soft decay with a thin layer of white mycelia grown on root tubers surface, subsequently, the middle part of the lesion became dark in later stages. The pathogen from a typical rotten root tuber was separated and purified by growth on potato dextrose agar (PDA). After 5 days culture at 28 °C in darkness, two purified fungal isolates on PDA were white, creamy, spreading, flat and smooth (Fig. 1D, F) with a pale yellow reverse side (Fig. 1E, G). A round bulge was also observed in the center of the colony. Both isolates showed similar colony, mycelium, and conidia morphologies. The end of the mycelium was broken into single spores or spore chains when mature. Moreover, cylindrical, barrel-shaped, or ellipsoidal single-cell arthrosporous were found, and the conidia size ranged from approximately 3.2–27.8 μm × 2.8–3.2 μm (n = 50) (Fig. 1H–J). Morphological features were consistent with Geotrichum candidum (G. candidum) as described by Thomidis et al.

Symptoms of naturally rotted root tubers and morphological aspects of the species G. candidum on potato dextrose agar (PDA). (A) Naturally rotted root tuber. (B) Rotted root tuber during drying. (C) The control root tuber (Red arrow) and the root tuber 8 days after inoculation with fungi in the field (Yellow arrow). (D) Colony of G. candidum R01 on PDA after seven days of growth. (E) Reverse colony of G. candidum R01 on potato dextrose agar. (F) Colony of G. candidum R02 on PDA after 7 days of growth. (G) Reverse colony G. candidum R02 on potato dextrose agar. (H) Fungal spores of G. candidum R01 (two types of spore, one was long tube-shaped, another was ellipsoidal oval and hyphae). (I) Long tube-shaped spore of G. candidum R01. (J) Long tube-shaped spore of G. candidum R02. Scale bars: (H–J) = 20 μm.
The DNA fragment was successfully amplified by using total genomic DNA as the template. A DNA-ITS sequence measuring approximately 400 base pairs was acquired from the two isolated DNA samples (Fig. 2A). Subsequently, sequenced the ITS of the two genes and aligned with entries in the National Center for Biotechnology Information (nih.gov) (http://www.ncbi.nlm.nih.gov). The BLASTn results showed that the obtained ITS region sequences from G. candidum R01 and G. candidum R02 all showed 100% identity with G. candidum strain KT336519.1. Thus, the two strains isolated are the same strain, which corroborated the morphological identification. Phylogenetic analysis results indicated that the isolated colonies were clustered with the G. candidum clade (Fig. 2B). Based on morphological and phylogenetic analyses of the ITS ___domain, the two isolates were identified as G. candidum. The EF-1α gene sequences were also amplified by PCR with EF1-728 F and EF1-986R primers (Fig. 2A). The representative sequences were submitted to GenBank and results suggested that the EF-1α sequence was 98% similar to a strain of Geotrichum candidum (accession numbers MT346370.1 and MT346368.1). Phylogenetic analysis results indicated that the isolated colonies were clustered with the G. candidum clade (Fig. 2C). Therefore, the isolate was identified as G. candidum R01.

Molecular identification of the fungal strain. (A) PCR results of ITS and EF-1α. (B) Phylogenetic tree based on nuclear ribosomal DNA-ITS region. (C) Phylogenetic tree based on the EF-1α region; M Marker DL 2000. (1, 2) Two replicates of ITS sequence. (3, 4) Two replicates of EF-1α sequence.
In Koch’s postulate experiments, healthy Xiaozhou mustard root tubers were inoculated with G. candidum in the field. The root tubers inoculated by G. candidum developed white rot symptoms and formed a layer of white mycelium after 8 days inoculation (Fig. 1C, yellow arrow), while no similar symptoms were observed in Xiaozhou mustard root tubers uninoculated with G. candidum (Fig. 1C, red arrow). All pathogens were re-isolated from artificially inoculated root tubers and exhibited the same morphological characteristics as the original isolate, as well as ITS gene sequences, fulfilling Koch’s postulates. The two fungal strains (G. candidum R01 and G. candidum R02) caused root tubers disease but no profound differences were observed between two isolate occurrences in root tuber tissue. Therefore, G. candidum R01 strain was selected for subsequent experiments in this work.
Antifungal activity of Cl-F-BBA against G. Candidum R01
After 5 days incubation in Cl-F-BBA, inhibition rate of mycelial growth in proportion to Cl-F-BBA concentration was increased significantly (Fig. 3A). The application of 0.10, 0.15, 0.20 and 0.25 mg/mL Cl-F-BBA induced mycelia growth inhibition rate by 73.93%, 84.31%, 96.27% and 100% of the control (p ˂ 0.05), respectively (Fig. 3B). Thus, 0.25 mg/mL Cl-F-BBA completely inhibited the colony expansion, with an EC50 value of 87.71 µg/mL. However, no significant difference in mycelial discs diameter was observed between 0.05 mg/mL Cl-F-BBA treated group and the control (Fig. 3A). Furthermore, 0.10, 0.15, 0.2, and 0.25 mg/mL Cl-F-BBA increased relative germination inhibition rates of spore significantly and recorded as 78.33%, 89.03%, 97.11% and 100% (p ˂ 0.05), respectively (Fig. 3C). In addition, Cl-F-BBA also demonstrated an effective inhibitory activity against Monilinia fructicola (M. fructicola), Aspergillus ochraceus (A. ochraceus), and Penicillinum citrinum (P. citrinum), with EC50 values of 32.98, 55.83, and 61.56 µg/mL, respectively (Additional file 1: Fig. S1, Table S1).

Effect of Cl-F-BBA on the mycelial growth inhibition rate (A, B) and relative germination inhibition rate of spore (C) of G. candidum R01. Different lowercase letters indicate significant differences (p < 0.05). Data are mean ± SD (n = 3).
Cl-F-BBA destroys the plasma membrane integrity and mycelial structure of G. Candidum R01
Propidium iodide (PI), a fluorescent molecule, is membrane impermeable and can bind to DNA, therefore, cell stained by PI can be used to quantify cell membrane damage. In the present study, significantly increased red signal intensity in mycelia and spores after treated with 0.20 mg/mL Cl-F-BBA were observed compared to the control (Fig. 4), indicating that Cl-F-BBA treatment damaged the cell membrane integrity of G. candidum R01 mycelia and spores.

Spores and mycelia of G. candidum R01 stained by PI. (A, E) Spores in control. (B, F) Mycelia in control. (C, G) Spores treated with 0.20 mg/mL Cl-F-BBA. (D, H) Mycelia treated with 0.20 mg/mL Cl-F-BBA. Scale bar = 20 μm.
The effect of Cl-F-BBA on G. candidum R01 mycelia morphology was also investigated by Scanning electron microscopy (SEM). The mycelial morphology of the control group showed a tubular, regular, homogeneous and smooth surface (Fig. 5A), whereas the groups treated with 0.20 mg/mL Cl-F-BBA showed irregular, twisted, and collapsed mycelia with a rough surface (Fig. 5B).

SEM images of G. candidum R01 mycelial morphology. (A) Mycelia in control. (B) Mycelia treated with 0.2 mg/mL Cl-F-BBA. Scale bar = 5 μm.
The membrane integrity of G. candidum R01 spores stained by PI was also determined by flow cytometry and was presented in two-dimensional dot plots (Fig. 6). It was showed that homogenous population of undamaged cells dominated in control group, while variations in cell sizes (forward scatter, [FSC]) and complexity (side scatter, [SSC]) were observed on scatter gram of G. candidum R01 spores after 12 h incubation in 0.2 mg/mL Cl-F-BBA. Moreover, PI-stained spores accounted for 6.98% of the total spores in the control (Fig. 6B, b), while the percentage of PI-stained spores treated by 0.20 mg/mL Cl-F-BBA increased to 90.42% (p < 0.05) (Fig. 6C and D), indicating that Cl-F-BBA can destroy the cell membrane integrity of G. candidum R01 spore.

The membrane integrity of G. candidum R01 spores determined by flow cytometry after treated with Cl-F-BBA. (A–C) Cell complexity (SSC) vs. cell size (FSC) showed by scatter gram. (a–c) PI-stained spores percentage of the control. (A, a) Autofluorescence of normal cells. (B, b) Fluorescence of normal cells for PI staining. (C, c) Spores treated with Cl-F-BBA. (D) PI-stained spores percentage after Cl-F-BBA treatment.
Cl-F-BBA damages the mitochondrial activity of G. Candidum R01
Compared to the spores (Figs. 5E and 7A) and mycelia (Fig. 7B, F) in the control group of G. candidum R01, the spores (Fig. 7C, G) and mycelia (Fig. 7D, H) treated with 0.20 mg/mL Cl-F-BBA showed reduced mitochondrial red fluorescence staining intensity, indicating the destruction of the active mitochondria.
In addition, the green fluorescence intensity of Rhodamine 123 was higher in the spores (Fig. 7I, M) and mycelia (Fig. 7J, N) in the control group, however, the green fluorescence intensity of Rhodamine 123 were significantly reduced both in the spores (Fig. 7K, O) and mycelia (Fig. 7L, P) after treated with 0.20 mg/mL Cl-F-BBA, suggesting that Cl-F-BBA can destroy the mitochondrial membrane structure and cause the mitochondrial dysfunction in G. candidum R01 cells.

G. candidum R01 stained with the mitochondrial red fluorescence probe and Rhodamine 123. (A, E) Spores in control. (B, F) Mycelia in control. (C, G) Spores treated with 0.20 mg/mL Cl-F-BBA. (D, H) Mycelia treated with 0.2 mg/mL Cl-F-BBA. (I, M) spores in control. (J, N) mycelia in control. (K, O) spores treated with 0.2 mg/mL Cl-F-BBA. (L, P) mycelia treated with 0.2 mg/mL Cl-F-BBA. Scale bar = 20 μm.
Cl-F-BBA decreases fungal respiration and the activities of MDH and SDH
Cl-F-BBA incubation significantly decreased the respiratory rate of G. candidum R01 (Fig. 8A). Thus, oxygen consumption rate was reduced by 18.56%, 41.61% and 63.17% of the control after Cl-F-BBA treatment for 12, 24 and 36 h. Meanwhile, Cl-F-BBA treatment caused 21.23%, 46.68% and 60.07% reduction of the control in the activity of malate dehydrogenase (MDH), and 38.12%, 60.84% and 73.96% reduction in succinate dehydrogenase (SDH) activity after 12, 24 and 36 h treatment (Fig. 8B, C). The decreases in the activities of two enzymes suggested that mitochondrial function was destroyed by Cl-F-BBA treatments.
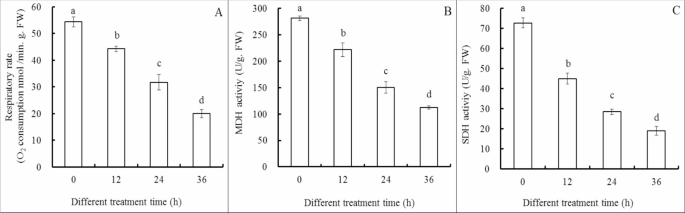
Cl-F-BBA decreases respiration, activities of MDH and SDH in G. candidum R01 cells. (A) Respiratory rate, (B) MDH activity, (C) SDH activity. Different letters above the bars indicate significant differences according to one-way analysis of variance (ANOVA), followed by Tukey’s test (P < 0.05).
Cl-F-BBA changes the contents of ATP, ADP, AMP and energy charge
Compared to the control, 0.20 mg/mL Cl-F-BBA decreased ATP content by 40.75% (p < 0.05) but increased ADP and AMP content by 23.47% and 273.72%, respectively (p < 0.05). Moreover, the energy charge decreased significantly in G. candidum R01 cells treated by 0.2 mg/mL Cl-F-BBA and was recorded as 25.64% compared to the control (p < 0.05) (Fig. 9).
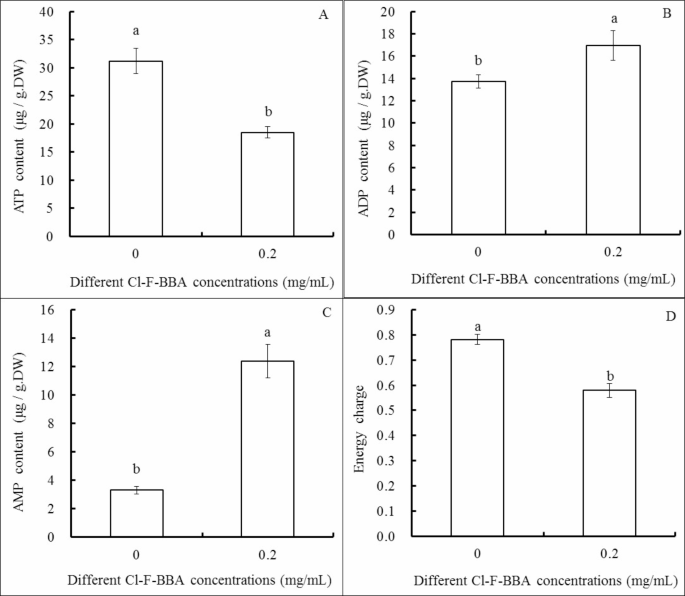
Effects of Cl-F-BBA on the contents of ATP (A), ADP (B), AMP (C) and energy charge (D) in G. candidum R01 cells. Different letters above the bars indicate significant differences according to one-way analysis of variance (ANOVA), followed by Tukey’s test (P < 0.05).
Cl-F-BBA disorder the antioxidant metabolism
The antioxidant enzymes activities of superoxide dismutase (SOD) (Fig. 10A) and catalase (CAT) (Fig. 10B) were significantly decreased in a time-dependent manner during 36 h Cl-F-BBA incubation. For instance, SOD activity was reduced by 20.74%, 42.01% and 73.07% of the control after Cl-F-BBA treatment for 12, 24 and 36 h. However, peroxidase (POD) activity was increased by 41.65% of the control after 12 h Cl-F-BBA exposure, then decreased by 42.31% and 72.84% of the control after 24 and 36 h exposure, respectively (Fig. 10C). Moreover, malondialdehyde (MDA) content was increased by 1.35-, 2.41-, and 2.83-times of the control group after 12, 24, and 36 h of Cl-F-BBA exposure, respectively (p < 0.05) (Fig. 10D). The decreased antioxidant enzymes and the increased MDA content suggested that Cl-F-BBA exposure reduced the antioxidant activity, which reduced the rate of ROS scavenging, caused the accumulation more reactive oxygen species (ROS) in the spore and mycelia (Fig. 11B, D), and further caused oxidative stress and led to cell membranes damage.
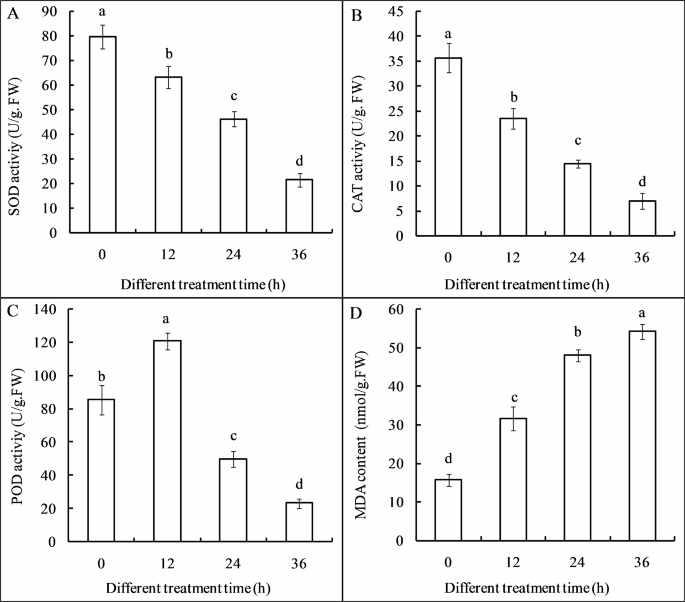
Effects of Cl-F-BBA on antioxidant enzymes activities and MDA content in G. candidum R01 mycelia. (A) SOD activity, (B) CAT activity, (C) POD activity, (D) MDA content. Different letters above the bars indicate significant differences according to one-way analysis of variance (ANOVA), followed by Tukey’s test (P < 0.05).
A specific fluorescent probe, H2DCFDA, was used to detect endogenous ROS in fungal cell. As showed in (Fig. 11), the fluorescence intensity of H2DCFDA was very weak in the cell of G. candidum R01 (Fig. 11A, C) in the control group. However, the cell of Cl-F-BBA treated group (Fig. 11B, D) showed extensive green fluorescence. In the meanwhile, ROS accumulation was also found in cell membranes of mycelia and spores, suggesting the cell membranes may be the target of ROS, and thus leads to more easy attack lipids in the membrane and induces lipids oxidation.

Effects of Cl-F-BBA on ROS accumulation in the spore and mycelia of G. candidum R01. (A, C) Control group. (B, D) 0.2 mg/mL Cl-F-BBA treated group for 36 h. Scale bar = 50 μm.
Cl-F-BBA increases intracellular content leakage in mycelia
The electrical conductivity increased by 1.58-, 2.09-, and 2.21 times of the control group after 4, 8, and 12 h Cl-F-BBA exposure (Fig. 12A), respectively (p < 0.05). The leakage of soluble sugar (Fig. 12B) and protein (Fig. 12C) in mycelia were both increased significantly after Cl-F-BBA treatment as compared with the control.
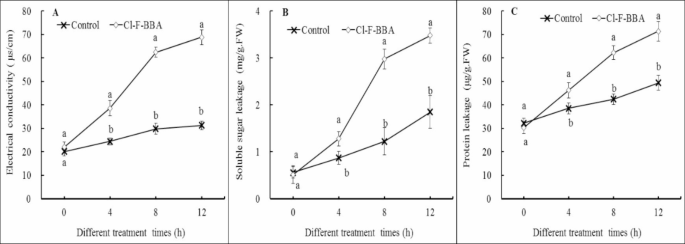
Effects of Cl-F-BBA on intracellular content leakage and the contents of soluble sugar and protein in G. candidum R01 mycelia. (A) Electrical conductivity. (B) Soluble sugar. (C) Protein. Different letters above the bars indicate significant differences according to one-way analysis of variance (ANOVA), followed by Tukey’s test (P < 0.05).
Cl-F-BBA against Xiaozhou mustard root tuber sour rot caused by G. Candidum R01
Rot lesions were decreased significantly in Xiaozhou mustard root tubers treated with Cl-F-BBA after 8 days inoculation with G. candidum R01 spores. The rot lesion diameter in the root tubers treated with 0.25, 0.50, 1.0 and 1.50 mg/mL Cl-F-BBA was decreased by 46.78%, 70.05%, 86.63%, and 100% (p < 0.05) of the control, respectively (Fig. 13A). In addition, the decay rate of root tubers was decreased significantly and disease occurrence was eliminated completely under 1.5 mg/mL Cl-F-BBA treatment (Fig. 13B). In the meanwhile, Cl-F-BBA also showed an excellent antifungal-efficacy on root tuber slices after inculated with G. candidum R01 (Additional file 2: Fig. S2). These data suggest that Cl-F-BBA is effective in against G. candidum R01 caused sour rotten in Xiaozhou mustard root tuber.

Cl-F-BBA treatment inhibited G. candidum R01 virulence on harvested Xiaozhou mustard root tuber. (A) Lesion diameter. (B) Decay rate. Different letters above the bars indicate significant differences according to one-way analysis of variance (ANOVA), followed by Tukey’s test (P < 0.05).
Discussion
G. candidum is a common fungus that causes sour rot in vegetables and fruits14,15. Previous reports revealed that chemical fungicides, e.g. propiconazole, showed high efficiency in preventing sour rot in tomatoes (Solanum lycopersicum)16, peaches (Prunus persica (L.) Batsch) and nectarine (Prunus persica var. Nectarina) fruits15. However, because of the increasing concern of consumers about pesticide residues in food, fungicides have been prevented from being the first choice for controlling fungal diseases. Recently, researchers have been increasingly interested in boron-containing compounds because of their potential antifungal properties. It has been proved that Benzaboronic acid exhibits remarkable antifungal properties, which may be due to the introduction of halogens to its benzene ring. Moreover, a study has shown that an electron-withdrawing substituent significantly enhanced antifungal activity compared with an electron-donating substituent, and the antifungal activity of PBA increased with the number of electron-withdrawing groups attached13.
The present study demonstrates that Cl-F-BBA has excellent antifungal activities against G. candidum R01 and three other plant pathogenic fungi (Additional file 1: Fig. S1, Table S1). At a concentration of 0.10 mg/mL, Cl-F-BBA significantly inhibited the colony expansion. Moreover, at 0.25 mg/mL, it completely stopped spore germination and mycelial growth. The work also explores the potential antifungal mechanisms of Cl-F-BBA against G. candidum R01. The results indicated that Cl-F-BBA treatment disrupted the antioxidant metabolism by decreasing antioxidant enzymes activities, while increasing ROS accumulation and MDA content, which improves oxidative stress in G. candidum R01. Similar results were reported by Ong et al.17, who revealed that Pyochelin inhibits Enterococcus faecalis growth by increasing ROS accumulation, leading to membrane lipid peroxidation and degradation, subsequently increasing MDA content, and accordingly leading to cell death by disrupting plasma membrane integrity. In addition, Cl-F-BBA disrupted the plasma membrane integrity, which induced increases in extracellular conductivity, total soluble sugar and protein contents, and eventually resulting in the destruction of osmotic balance as well as a loss of small molecules. These results are consistent with Du et al.13, who found one kind of phenylboronic acid derivatives treatment disturbed cell membrane integrity of Botrytis cinerea. Similarly, Qin et al.8 reported that BA inhibited Botrytis cinerea growth through the loss of cytoplasmic materials from hyphae. In addition, Cl-F-BBA treatment also destroyed the integrity of the mitochondrial membrane (Fig. 7), subsequently leading to mitochondrial degradation and decreased mitochondrial function7,18. Meanwhile, coupled with mitochondrial membrane structural damage, Cl-F-BBA also caused a reduction in the intracellular ATP content and energy charge. A decrease in energy levels coinciding with a decrease in membrane stability caused by fibroin treatment to inhibit chilling injury in banana fruit was also reported by Liu et al.19. Based on these findings, it is suggested that the destruction of membrane integrity in cells and mitochondria, and the disturbance of energy metabolism might be related to the antifungal mechanism of Cl-F-BBA against G. candidum R01.
In addition, Cl-F-BBA also decreased the activities of key enzymes of MDH and SDH involved in tricarboxylic acid (TCA) cycle, and reduced the respiratory rate, the intracellular ATP content and energy charge. MDH catalyzes the interconversion of malate to oxaloacetate, which is essential for energy balance20, while SDH catalyzes the oxidation of succinate to fumarate in the mitochondrial matrix21, which connects the TCA cycle to the electron transport chain22,23. Therefore, the dysfunction of MDH and SDH can impair the mitochondrial activity, the respiratory rate, and ATP generation in cells. Peng et al.24 showed that pinocembrin decreased the respiration rate of Penicillium italium, thus the oxygen consumption rate was much lower than that of the control. Similar findings were also reported by Stanic et al.25, which indicated that anion channel inhibitors inhibited Phycomyces blakesleeanus growth by decreasing respiration and energy metabolism. Our findings also align with those of He et al.26, who demonstrated that linalool suppresses the respiratory activity of Pseudomonas lundensis and inhibits key enzymes in the TCA cycle, including SDH, MDH, and citrate synthase, thereby disturbing the energy supply. Overall, we speculate that the sharp decline in key enzyme activities involved in the TCA cycle coupled with the loss of ATP in G. candidum, leads to the lack of energy and eventually results in growth inhibition and cell death.
Furthermore, our work also showed the effect of Cl-F-BBA on disease in harvested Xiaozhou mustard root tubers after artificial inoculation with pathogen spores, which was in accordance with the results obtained on PDA plates. In addition, we also found that root tuber slices diseased after 3 days of inoculation with G. candidum, while it took longer for lesions to appear in the full-root tubers in the field, suggesting that the epidermis of the root tubers has a protective effect. In addition, Cl-F-BBA had a good antifungal effect on root slices after they were incubated with G. candidum (Additional file Figs. 2 and 3). Therefore, Cl-F-BBA treatment is a highly efficient and environmentally safe strategy to replacing conventional fungicides for controlling sour rot of Xiaozhou mustard root tuber caused by G. candidum infection.
Conclusions
Cl-F-BBA exhibits excellent antifungal activity by inducing oxidative stress and excessive ROS accumulation, damaging cellular macromolecules. It also reduces the activities of MDH and SDH, ATP content, and energy charge, indicating a blockade of the TCA cycle pathway in fungal cells. The antifungal effects of Cl-F-BBA may stem from the disruption of cell membrane integrity and energy metabolism. Thus, Cl-F-BBA presents a promising fungicide alternative for managing G. candidum-induced sour rot in Xiaozhou mustard root tubers.
Materials and methods
Isolation and identification of fungi
Rotten Xiaozhou mustard root tubers showing a depression and with black lesion were taken from vegetable gardens in Changan Town, Rongan County, Guangxi Province in December 2022. The pathogens were separated from naturally infected Xiaozhou mustard root tubers tissues when they decayed with typical diseases. Briefly, to isolate the pathogen, three symptomatic partially rotted root tubers from three gardens were collected. The root tubers were rinsed twice with distilled water and then surface-disinfected by wiping them with 75% (v/v) ethanol. The root tuber tissues from the margin of lesions was cutted into small pieces (4 × 4 × 3 mm). Six pieces were disinfested with 75% ethanol for 2 min and 2% NaClO for 5 min, rinsed 3 times with sterile water. The tissues were incubated on PDA medium at 28 °C for 5 days in darkness until sporulation. Colonies were transferred to fresh PDA dishes for single-spore isolation until two uniform isolates were obtained. The morphology of the colony and spore was observed under a microscope.
Molecular identification for two uniform isolates was performed as reported by Guo et al.27. Briefly, the ITS region gene was amplified using primers ITS1/ITS4, and the EF-1α gene was amplified with EF1-728 F and EF1-986R primers28. Total genomic DNA of the fungal isolates were extracted by using a DNeasy Kit. PCR were performed using the method described by Guo et al.27. The PCR amplified products were sequenced by Tsingke Biotech (Hangzhou, China). ITS and EF-1α sequences were used for BLAST searches in GenBank database (http://www.ncbi.nlm.nih.gov/BLAST), and obtained results were compared with similar sequences in NCBI database (http://www.ncbi.nlm.nih.gov). A phylogenetic tree was constructed by using MEGA 7.0 software (https://www.megasoftware.net/). Pathogenicity analysis was conducted with two isolates on healthy Xiaozhou mustard root tubers. Xiaozhou mustard root tubers were surface-disinfected with 75% (v/v) alcohol. Wounds were created in the middle of the branches using a sterilized hole punch (4 mm diameter) and ten healthy Xiaozhou mustard root tubers (20 µL per root tuber) were immediately inoculated with spore suspensions (1 × 106 spores/mL) of two isolates, respectively. Ten root tubers were used for each isolate. The control root tubers were inoculated with 20 µL sterilized water. All treated root tubers were kept under general agronomic conditions for 8 days. When symptoms similar to natural disease in the field were observed in the inoculated root tube, the pathogen was re-isolated on PDA, and the morphological features and ITS gene sequences of isolated fungi were compared with the original ones used for inoculation to confirm Koch’s postulates. This experiment was repeated three times.
Determination of minimal concentration of Cl-F-BBA effect on G. Candidum R01
2-chloro-5-trifluoromethoxybenzeneboronic acid (Cl-F-BBA) was purchased from Shanghai Bide Pharmaceutical Technology Co., Ltd. (China). Minimal concentration (MIC) determination of Cl-F-BBA was assayed according to Du et al.13 with some modification. Briefly, Cl-F-BBA was dissolved in 3.0 mL DMSO firstly and then in sterilized distilled H2O to obtain a 20 mg/mL stock solution. An appropriate volume Cl-F-BBA stock solution was pipetted into 60 ml melted PDA to obtain the desired Cl-F-BBA concentrations of 0.0, 0.05, 0.1, 0.15, 0.2, 0.25, 0.5, 0.75, 1.0 and 2.0 mg/mL. Agar plugs (5 mm) with actively growing mycelia of G. candidum R01 from 6-day-old colonies grown on PDA, and then transferred to upside down at the centre of a new PDA plates amended with a varying Cl-F-BBA concentrations(0.0, 0.05, 0.1, 0.15, 0.2, 0.25, 0.5, 0.75, 1.0 and 2.0 mg/mL), three plates per concentration, and with two replicates each. The colony diameter was measured after culture for five days at 28 °C. The percentage of inhibition rate of G. candidum R01 mycelial growth was assayed according to the formula reported by Song et al.16 : MGI (%) = 100%×(d0 − dt)/d0. Where d0 and dt are the average diameters of the fungal colonies of the blank control group and treated group, respectively. The lowest concentration that completely restrained the growth of G. candidum R01 was regarded as minimum inhibitory concentration (MIC). In the meanwhile, for calculation of the effective concentrations of Cl-F-BBA causing a 50% reduction (EC50) in mycelial growth, concentrations of Cl-F-BBA were logarithmically transformed and percent inhibitions were converted to probit values29. EC50 values were estimated with the Probit procedure in the statistical software Data Processing Syste (DPS, ver. 7.05, Hangzhou Ruifeng Information Technology Co. Ltd.) according to Li et al.30.
Determination of the antifungal efficacy of Cl-F-BBA against spore germination
The effects of Cl-F-BBA on the spore germination of G. candidum R01 were evaluated according to Jiao et al.31. Briefly, six concentrations of Cl-F-BBA at 0.0, 0.05, 0.1, 0.15, 0.2 and 0.25 mg/mL were prepared in sterile 10 mL of potato dextrose broth mediums. G. candidum R01 spores were suspended in PDB to get spore suspension (1 × 106 spores/ml). Then, the samples in cotton-plugged glass tubes were cultivated at 28 °C on a rotary shaker (90 r/min). After incubation for 2.5 h, the spore were washed twice with sterile water and re-suspended in sterile water. Afterwards, 20 µL spore suspension was dropped onto a sterilized concave slide, the number of spores germinated was recorded by a 40-fold optical microscope. From each concentration of Cl-F-BBA treatment, more than 200 spores were counted, a spore was considered to have germinated when the length of the germinal tubule reached twice that of the total spore diameter. The relative inhibition percentage of spore germination was calculated. relative germination inhibition rate (%) = 100%× (G0-Gt)/G0, where G0 and Gt are the average values of the spore germination rates of the blank control group and treated group, respectively. Each concentration was replicated in three times.
Determination of plasma membrane integrity
Our previous results demonstrated that treatments with 0.20 mg/mL Cl-F-BBA can compromise the integrity of fungal cell membranes. Mycelia and spores from 4-day-old cultures of G. candidum R01, grown in PDA medium treated with 0.2 mg/mL Cl-F-BBA and were stained with 10.0 µg/mL Propidium Iodide (PI) for 25 min, following the method described by Kong et al.32. The mycelia and spores were observed under fluorescence microscope.
To more accurately assess spore membrane integrity, G. candidum R01 spores were stained with PI (10.0 µg/mL) for 25 min. Subsequently, a cyto-flex flow cytometer was used for sorting, analyzing, and calculating, as outlined by Guo et al.27.
Analysis of mycelial morphology by scanning electron microscopy
For scanning electron microscopy (SEM), G. candidum R01 mycelia grown after 4 days of culture with 0 mg/L and 2.0 mg/mL Cl-F-BBA were collected. Samples were subjected to series dehydration steps using the method described by Xu et al.33. The mycelia were then freeze-dried by sublimation in a freeze dryer for 48 h and gold-coated for 60 s using a sputter coater (E1010, Hitachi, Japan). Mycelia were observed by SEM (JSM-6360LV, JEOL, Japan) using the method introduced by Li et al.34.
Determination of mitochondrial activities
G. candidum R01 treated with 0.0 (control) and 0.20 mg/mL Cl-F-BBA were grown on PDA for 4 days. Subsequently, mycelia and spores were stained with 50 µg/mL Rhodamine 123 for 30 min and 10 µg/mL mitochondrial red fluorescence probe for 25 min, respectively. The mitochondrial activities were assessed by observing the differences in staining intensity under a fluorescence microscope14. Each treatment was replicated three times, and the entire experiment was conducted in triplicate.
Determination of pathogen respiration
Respiration was measured according to the method described by Peng et al.24. The mycelia were washed with ddH2O and suspended in 20 mmol/l HEPES-TRIS buffer (containing 250 mmol/L sucrose, 10.0 mmol/L Tris-HCl, 5.0 mmol/L KH2PO4, 1.0 mmol/l MgCl2, pH 7.2), then, Cl-F-BBA was added into the mycelial suspensions. Oxygen consumption assay was carried out at 28 °C with a Clark-type oxygen electrode. Respiratory rate is expressed as the oxygen consumption rate.
Determination of the activities of SDH and MDH
The mitochondria preparation from mycelia was conducted according to the methods described by Guo et al.27. The activities of SDH and MDH in mitochondria were determined following the detection Kit manufacturer’s instructions (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
Determination of ATP-, ADP-, and AMP content, as well as energy charge in mycelia
Adenosine triphosphate (ATP), adenosine diphosphate (ADP), and adenosine monophosphate (AMP) were extracted from mycelia cultured for 5 days on PDA after treatment with 0 g/L and 0.20 mg/mL Cl-F-BBA. The content of ATP, ADP and AMP were determined by HPLC according to the methods described by Guo et al.27. Standard ATP, ADP, and AMP (Sigma, USA) were used for quantitative detection, and their concentrations were determined using the external standard method. ATP, ADP, and AMP content was used to calculate the energy charge using the following equation: energy charge = (ATP + 0.5 × ADP)/ (ATP + ADP + AMP) according to previous methods35. Three replicates were performed in each treatment, and the experiment was performed in triplicate.
Determination of antioxidant enzyme activity, MDA content and reactive oxygen species
6.0 g of fresh weight mycelia was suspended in 50 mL sterile ddH2O supplemented with 0.20 mg/mL Cl-F-BBA. Sterile ddH2O without Cl-F-BBA was used for the control. The suspension was placed on a rotary shaker incubator (160 r/m at 28 °C) for 0, 12, 24, and 36 h, the collected mycelia (1.0 g) was used to measure antioxidant enzyme activities and MDA content. The activities of SOD, CAT, and POD were measured according to the methods described by Mo et al.36, while MDA content was measured as described by Elsherbiny et al.37. To evaluate ROS accumulated in cell of pathogen, the spore and mycelia, treated with 0.20 mg/ml Cl-F-BBA for 36 h, were collected and stained by 10.0 µmol/L 2, 7-dichlorodihydro-fluorescein diacetate (H2DCFDA) for 30 min. Subsquently, the endogenous ROS were labeled with green fluorescence and observed by a fluorescence microscope.
Determination of the intracellular content leakage in mycelia
0.5 mL of spore suspensions (1 × 106 spores/mL) were added to flasks containing 100 mL PDB, and the flasks were placed on a rotary shaker for 3 days (150 r/m at 28 °C). Then, the mycelia were collected and washed three times with ddH2O. Mycelia were filtered in a vacuum using a 20 μm sterile filter for 25 min. Intracellular content leakage was determined as described by Gao et al.38 with slight modifications. Here, 2.0 g of fresh weight mycelia were transferred to 150 mL flasks containing 50 mL sterile ddH2O with 0.20 mg/mL Cl-F-BBA. Sterile ddH2O without Cl-F-BBA was used for the control. The flasks were placed on a rotary shaker (130 r/m at 28 °C) for 0, 4, 8, and 12 h, the electrical conductivity was measured with a conductivity meter, and expressed as the amount of extracellular conductivity (µs/cm). Aliquots of mycelia suspensions were sampled after 0, 4, 8, and 12 h of incubation at 130 r/m at 28 °C. The suspension was filtered through 0.2-µm spore size membrane. After then, the filtrate was used to measure the total soluble proteins and total soluble sugar. The soluble sugar content was determined using the phenol-sulfuric acid method39, and soluble protein content was determined according to Bradford40. Three replicates were performed in each treatment.
Measurement of Cl-F-BBA against Xiaozhou mustard root tuber rot in vivo
The efficacy of Cl-F-BBA in managing postharvest disease in Xiaozhou mustard root tubers was evaluated by using methodologies reported by Wang et al.4 and He et al.41. The root tubers were first wounded with a sterilized punch, followed by the injection of 15 µL spore suspension containing 1 × 106 spores/mL into each wound. Then, after root tubers were air-dried for 1.0 h, 40 µL Cl-F-BBA (0.0, 0.25, 0.50, 1.0, and 1.5 mg/mL) of was added to the same wound. Sterile ddH2O treated root tubers were used as the control. The treated root tubers were packed in different polyethylene plastic bags and stored in a growth chamber maintained at 28 °C with 95% relative humidity under darkness. The lesion diameter (mm) and decay rate of root tubers were measured after 8 days of storage. Three replicates were performed in each treatment.
Statistical analyses
All experiments were repeated at least three replicates. Data were analyzed using one-way ANOVA in SPSS 23 Software package (SPSS Inc, Chicago, IL, USA). Statistical significance was followed by Tukey’s HSD test at p < 0.05 to examine differences between treatments.
Data availability
The data that support the findings of this study are contained within the manuscript and supplementary information files.
Change history
03 December 2024
A Correction to this paper has been published: https://doi.org/10.1038/s41598-024-81268-3
References
Xu, Y. et al. Effects of 1-methylcyclopropene treatment on quality and anthocyanin biosynthesis in plum (Prunus salicina Cv. Taoxingli) fruit during storage at a non-chilling temperature. Postharvest Biol. Technol.169, 111291 (2020).
Huang, Y., Fan, Z., Cai, Y., Jin, L. & Yu, T. The influence of N-acetyl glucosamine: inducing Rhodosporidium paludigenum to enhance the inhibition of Penicillium expansum on pears. Postharvest Biol. Technol.176, 111486 (2021).
Roberto, S. R., Youssef, K., Hashim, A. F. & Ippolito, A. Nanomaterials as alternative control means against postharvest diseases in fruit crops. Nanomaterials 9, 1752 (2019).
Wang, W., Ling, Y., Deng, L., Yao, S. & Zeng, K. Effect of L-cysteine treatment to induce postharvest disease resistance of Monilinia fructicola in plum fruits and the possible mechanisms involved. Pest Biochem. Phys.191, 105367 (2023).
Li, Y. et al. Tea tree oil exhibits antifungal activity against Botrytis cinerea by affecting mitochondria. Food Chem.234, 62–67 (2017).
Gavilanes-Martínez, M. A., Coral-Garzón, A., Cáceres, D. H. & García, A. M. Antifungal activity of boric acid, triclosan and zinc oxide against different clinically relevant Candida Species. Mycoses 64, 1045–1052 (2021).
Lai, T. et al. Dissecting inhibitory effect of boric acid on virulence and patulin production of Penicillium expansumPostharvest Biol. Technol.17, 187–196 (2016).
Qin, G., Zong, Y., Chen, Q., Hua, D. & Tian, S. Inhibitory effect of boron against Botrytis cinerea on table grapes and its possible mechanisms of action. Int. J. Food Microbiol.138, 145–150 (2010).
Thomidis, T. & Exadaktylou, E. Effect of boron on the development of brown rot (Monilinia laxa) on peaches. Crop Prot.6, 572–576 (2010).
Adamczyk-Woźniak, A., Komarovska-Porokhnyavets, O. & Misterkiewicz, B. Biological activity of selected boronic acids and their derivatives. Appl. Organomet. Chem.26, 390–393 (2012).
Adamczyk-Woźniak, A. et al. Trifluoromethoxy) phenylboronic acids: structures, properties, and antibacterial activity. Molecules 26, 2007 (2021).
Martinko, K., Ivanković, S., Lazarević, B., Ðermić, E. & Ðermić, D. Control of early blight fungus (Alternaria alternata) in tomato by boric and phenylboronic acid. Antibiotics 11, 320 (2022).
Du, S. S. et al. Exploring boron applications in modern agriculture: antifungal activities and mechanisms of phenylboronic acid derivatives. Pest Manag. Sci.79, 2748–2761 (2023).
Song, S. et al. First report of a new potato disease caused by Galactomyces candidum F12 in China. J. Integr. Agric.19, 2470–2476 (2020).
Thomidis, T., Prodromou, I., Farmaki, A. & Zambounis, A. Effect of temperature on the growth of Geotrichum candidum and chemical control of sour rot on tomatoes. Trop. Plant Pathol.46, 545–552 (2021).
Yaghmour, M. A., Bostock, R. M., Adaskaveg, J. E. & Michailides, T. J. Propiconazole sensitivity in populations of Geotrichum candidum, the cause of sour rot of peach and nectarine, in California. Plant Dis.96, 752–758 (2012).
Ong, K. S., Cheow, Y. L. & Lee, S. M. The role of reactive oxygen species in the antimicrobial activity of pyochelin. J. Adv. Res.8, 393–398 (2017).
Shi, X. Q., Li, B. Q., Qin, G. Z. & Tian, S. P. Mechanism of antifungal action of borate against Colletotrichum gloeosporioides related to mitochondrial degradation in spores. Postharvest Biol. Technol.67, 138–143 (2012).
Liu, P. et al. Fibroin treatment inhibits chilling injury of banana fruit via energy regulation. Sci. Hortic.248, 8–13 (2019).
Moriyama, S., Nishio, K. & Mizushima, T. Structure of glyoxysomal malate dehydrogenase (MDH3) from Saccharomyces cerevisiae. Acta Crystallogr. Sect. F Struct. Biol. Commun.74, 617–624 (2018).
Zhou, M. et al. The self-association of cytoplasmic malate dehydrogenase 1 promotes malate biosynthesis and confers disease resistance in cassava. Plant. Physiol. Biochem.201, 107814 (2023).
Sun, F. et al. Crystal structure of mitochondrial respiratory membrane protein complex II. Cell 121, 1043–1057 (2005).
Fu, W. et al. Reversible regulation of succinate dehydrogenase by tools of photopharmacology. J. Agric. Food Chem.70, 4279–4290 (2022).
Peng, L. et al. Antifungal activity and action mode of pinocembrin from Propolis against Penicillium italicumFood Sci. Biotechnol.21, 1533–1539 (2012).
Stanić, M. et al. Growth inhibition of fungus Phycomyces Blakesleeanus by anion channel inhibitors anthracene-9-carboxylic and niflumic acid attained through decrease in cellular respiration and energy metabolites. Microbiology 163, 364–372 (2017).
He, R. et al. Respiratory depression driven by membrane damage as a mechanism for linalool to inhibit Pseudomonas lundensis and its preservation potential for beef. J. Appl. Microbiol.134, 1093 (2023).
Guo, T. R. et al. Isolation, identification, biological characteristics, and antifungal efficacy of sodium bicarbonate combined with natamycin on Aspergillus Niger from Shengzhou nane (Prunus salicina var. Taoxingli) fruit. Front. Microbiol.13, 1075033 (2023).
Carbone, I. & Kohn, L. M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 91, 553–556 (1999).
Hu, S., Zhang, J., Zhang, Y., He, S. & Zhu, F. Baseline sensitivity and toxic actions of boscalid against Sclerotinia sclerotiorum. Crop Prot. 110, 83–90 (2018).
Li, J. L. et al. Baseline sensitivity and control efficacy of DMI fungicide epoxiconazole against Sclerotinia sclerotiorumEur. J. Plant. Pathol.141, 237–246 (2015).
Jiao, W. et al. Epsilon-poly-L-lysine (ε-PL) exhibits antifungal activity in vivo and in vitro against Botrytis cinerea and mechanism involved. Postharvest Biol. Technol.168, 111270 (2020).
Kong, J. et al. Antifungal effects of thymol and salicylic acid on cell membrane and mitochondria of Rhizopus stolonifer and their application in postharvest preservation of tomatoes. Food Chem.285, 380–388 (2019).
Xu, Y. et al. Tea tree oil controls brown rot in peaches by damaging the cell membrane of Monilinia fructicolaPostharvest Biol. Technol.175, 111474 (2021).
Li, W. et al. Antifungal activity and mechanism of tetramycin against Alternaria alternata, the soft rot causing fungi in Kiwi fruit. Pest Biochem. Phys.192, 105409 (2023).
Hu, W. et al. Yarrowia Lipolytica reduces the disease incidence of asparagus infected by Fusarium proliferatum by affecting respiratory metabolism and energy status. Biol. Control159, 104625 (2021).
Mo, Y. et al. Enhanced preservation effects of sugar apple fruits by salicylic acid treatment during post-harvest storage. J. Sci. Food Agric.88, 2693–2699 (2008).
Elsherbiny, E. A., Dawood, D. H. & Safwat, N. A. Antifungal action and induction of resistance by β-aminobutyric acid against Penicillium digitatum to control green mold in orange fruit. Pesticide Biochem. Physiol.171, 104721 (2021).
Gao, Y. et al. Toxicity and biochemical action of the antibiotic fungicide tetramycin on Colletotrichum scovillei. Pest Biochem. Phys.147, 51–58 (2018).
Hodges, D. M., DeLong, J. M., Forney, C. F. & Prange, R. K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 207, 604–611 (1999).
Bradford, M. N. A rapid and sensitive method for the quantitation of microgram quantities of protein using the principle of protein-dye binding. Anal. Biochem.6, 3177–3188 (1976).
He, C., Zhang, Z., Li, B., Xu, Y. & Tian, S. Effect of natamycin on Botrytis cinerea and Penicillium expansum-postharvest pathogens of grape berries and jujube fruit. Postharvest Biol. Technol.151, 134–141 (2019).
Funding
This work was supported by public welfare projects of Zhejiang Province in China (No. LGN22C150002).
Author information
Authors and Affiliations
Contributions
Authors have contributed to this work in following roles: Y.W. andT.R. designed the study. Q.Y., Z.H, and F.X. performed the experiments. G. and K.Y. analysed data. Y.W. wrote the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this Article was revised: The original version of this Article contained an error in an equation in the Materials and methods section, under the subheading ‘Determination of ATP-, ADP-, and AMP content, as well as energy charge in mycelia’. Full information regarding the correction made can be found in the correction for this Article.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Mo, Q., Xiao, Z., Ou, K. et al. Antifungal activity of 2-chloro-5-trifluoromethoxybenzeneboronic acid and inhibitory mechanisms on Geotrichum candidum from sour rot Xiaozhou mustard root tuber. Sci Rep 14, 22802 (2024). https://doi.org/10.1038/s41598-024-74211-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-74211-z
