
Abstract
Post-translational SUMO modification is a widespread mechanism for regulating protein function within cells. In humans, SUMO-conjugated proteins are partially regulated by the deconjugating activity of six SENP family members. The proteolytic activity of these enzymes resides within a conserved catalytic ___domain that exhibits specificity for the two primary SUMO isoforms: SUMO1 and SUMO2/3. SENP5, along with SENP3, are nucleolar proteins involved in ribosome biogenesis and preferentially target SUMO2/3 isoforms. Here, we present the crystal structures of human SENP5 in complex with both SUMO1 and SUMO2 isoforms. These structures reveal a minimal complex interface and elucidate the molecular basis for SENP5’s preference for the SUMO2 isoform. This preference can be attributed to a basic patch surrounding SENP5 Arg624 at the interface. Swapping mutagenesis and structural analysis demonstrate that Arg624 is favorably oriented to interact with Asp63 in SUMO2/3, while its interaction with the equivalent Glu67 in SUMO1 is less favorable. These results suggest that subtle structural differences within SUMO isoforms can significantly influence their deconjugation by SENP enzymes, opening new avenues for exploring the regulation of SUMOylation in various cellular processes and for developing therapeutic agents targeting SUMOylation pathways.
Similar content being viewed by others

Introduction
SUMOylation is a post-translational modification wherein the Small Ubiquitin-like Modifier (SUMO) is covalently attached to lysine residues of target proteins. Like ubiquitin, SUMO is conjugated via its C-terminal tail through a multi-step enzymatic cascade involving E1 activation, E2 conjugation, and E3 ligation1. In humans, SUMOylation plays a pivotal role in diverse cellular processes, including DNA replication, nuclear transport, and DNA damage response2,3,4. SUMOylation typically modulates protein-protein interactions by binding to specific SUMO interaction motifs (SIMs)5,6,7. This modification can be reversed by deSUMOylating proteases that cleave the isopeptidic bond between SUMO and the target lysine8,9,10,11. SUMO is a ubiquitous modification found across eukaryotes. While humans encode four SUMO isoforms (SUMO1, SUMO2, SUMO3, and SUMO4), yeast and invertebrates have only one (Smt3), and plants have multiple12,13. Notably, SUMO2 and SUMO3 share a high degree of sequence similarity (97%, collectively referred to as SUMO2/3 after maturation), while SUMO1 exhibits lower similarity (47%). Interestingly, the inability of SUMO4 to be processed prevents it from forming SUMO conjugates14.
The SENP/ULP protease family constitute the major group of deSUMOylating enzymes within human cells8,9,15,16,17. They are composed by long unstructured regions followed by a conserved catalytic ___domain that belongs to the CE-clan of cysteine proteases, containing the characteristic active site catalytic triad (His-Asp-Cys)18. Similar to ULP1, the founding member discovered in Saccharomyces cerevisiae19,20, the SENP/ULP family in humans is constituted by six members that can be divided into three subfamilies based on phylogenesis and sequence homology: SENP1 and SENP2; SENP3 and SENP5; and SENP6 and SENP79. However, in the recent years, two novel classes of SUMO proteases have been discovered in human cells non-related to the SENP/ULP family: the deSUMOylating peptidase 1 and 2 (DESI1 and DESI2)21; and the deubiquitinating enzyme USPL1, which is active for SUMO22,23. The SENP/ULP family members are responsible of a dual proteolytic function: processing of SUMO precursors, by cleaving off the α-peptide bond in the C-terminus of SUMO precursors (preSUMO1, preSUMO2, and preSUMO3) to expose the diglycine motif (mature SUMO); and the deconjugating activity of SUMO targets, by cleaving off the isopeptide bond between SUMO and the lysine residue of substrates24,25,26,27. Whereas all six members of the SENP/ULP family possess isopeptide or deconjugating activity, only the SENP1 and SENP2 subfamily are able to efficiently process the immature SUMO precursors, displaying different activities depending on the type of SUMO isoforms26,28,29,30.
Substrate specificity in the SENP/ULP family is mostly regulated by localization of the different members within the cell, which is mostly determined by the unstructured regions outside the conserved catalytic ___domain8,9. In some cases, alternative splicing or post-translational modification contribute to the spatial control of the deconjugation activity24,31,32,33,34. For example, SENP1 and SENP2 are enriched at the nuclear pore complex and in the PML nuclear bodies during the interphase, but they accumulate at the kinetochore during mitosis27,35,36,37. SENP3 and SENP5 are mostly compartmentalized within the granular component of the nucleolus, although it can also be found at the mitochondria38,39,40,41,42. SENP6 and SENP7 are mainly located at the nucleoplasm, where they interact with chromatin regulating gene expression25,43. Alternatively, short splicing variants of SENP2, SENP5, and SENP7 have also been found at the cytoplasm35,44,45,46.
Regarding SENP3 and SENP5 known functions, knockdown of SENP5 inhibits cell proliferation, having important defects in nuclear morphology and in cell division47,48. The unstructured N-terminal region of SENP5 has a major role in the restriction of SUMO deconjugation activity to the nucleolus47, where together with SENP3, have been involved in ribosome and pre-rRNA biogenesis processes49,50. SENP3 and SENP5 were initially found to directly interact with B23/nucleophosmin40. More recently, a proteomics analysis have identified different subsets of SUMOylated trans-acting factors involved in ribosome biogenesis controlled by either SENP3 or SENP550. Their data suggest that SENP5 predominantly participates in the 40S particle assembly, whereas SENP3 is connected to both 40S and 60S maturation steps50. SENP5 has also been involved in the regulation of the mitochondrial fission during mitosis by regulating the activity of the GTPase Drp141,42. A short splicing variant of SENP5 lacking the catalytic ___domain has been shown to competitively regulate the SUMO conjugation of Drp146. Recently, SENP5 and SENP3 have also been reported to regulate the SUMOylation levels of Aurora A kinase and thus control its activity during mitosis51.
The structural and functional characterization of SENP/ULP family members has revealed that their specificity towards SUMO isoforms (SUMO1 vs. SUMO2/3) is primarily determined by their conserved catalytic domains25,28,29,30,52,53,54. These domains exhibit distinct binding affinities for each SUMO isoform. Within the three SENP subfamilies, only SENP1 and SENP2 demonstrate efficient deconjugation of both SUMO1 and SUMO2/3 isoforms28,29,30. SENP6 and SENP7 possess a unique loop insertion sequence within their catalytic domains, which expands the protein-protein contact interface and confers specific activity towards SUMO2/3 conjugates25,52,53,54. Interestingly, the SENP3 and SENP5 subfamily also exhibits a preference for deconjugating SUMO2/3 over SUMO1 in vitro, despite lacking the loop insertion characteristic of SENP738,40,55.
Here, we present the crystal structures of SENP5 in complex with SUMO1, SUMO1 E67D mutant, and SUMO2 isoforms at resolutions of 2.1, 1.7, and 2.3 Å, respectively. The SENP5 complex interface is notably compact, representing the smallest among known SENP/ULP member complex structures. Intriguingly, a specific basic patch surrounding Arg624 in SENP5 appears to be crucial for its preference for the SUMO2 isoform, forming a favorable ion pair as evidenced by the structure of a complex with an engineered SUMO1 E67D mutant. Our findings hold potential for the development of more selective inhibitors and probes targeting individual members of the human SENP family, such as SENP5, which play essential roles in various cellular processes.
Results
SENP5 prefers SUMO2 deconjugation
The full-length sequence of human SENP5 comprises 755 residues, and based on sequence alignments with SENP/ULP family members and on AlphaFold-2 prediction, the structure of SENP5 is composed by a N-terminal unstructured region followed by the canonical catalytic ___domain (Supplementary Fig. 1). Several constructs of the SENP5 catalytic ___domain with different N-terminal truncations have been analyzed by recombinant expression, including a construct with a 3-helix bundle ___domain predicted by AlphaFold-2 located N-terminal to the catalytic ___domain (from residue 493 to 755), which might participate in substrate binding (Supplementary Fig. 2). However, only the shortest SENP5 catalytic ___domain construct (from residue 568 to 755), yielded soluble protein in Escherichia coli and ultimately SUMO proteolytic activity in in vitro assays. This shorter SENP5 construct lacks some N-terminal secondary structure elements present in the catalytic ___domain of other members of the human SENP family, like a N-terminal α-helix and a short β-structure. These elements are relevant for SUMO binding in other family members, such as SENP1, SENP2, and SENP7, in the latter playing a major role in the SUMO2 isoform specificity28,29,30,52.
In vitro activity assays using the catalytic ___domain of SENP5 (named SENP5CD from now on) revealed limited proteolytic processing of the three SUMO isoform precursors. Only at high SENP5CD concentrations (150 nM) processing activity is observed for preSUMO2, while preSUMO1 and preSUMO3 remain largely unprocessed (Supplementary Fig. 3). In contrast, SENP5CD exhibited robust isopeptidase activity toward SUMO2/3-conjugated substrates, evidenced when comparing time course reactions with model substrates RanGAP1-SUMO1 and RanGAP1-SUMO2/3, or with SUMO1 and SUMO2-AMC substrates at 5 nM or 50 nM SENP5CD concentrations respectively (Fig. 1a, b). Similar isopeptidase activity is observed for a diSUMO2 substrate, indicating its potential to process SUMO2 chains (Supplementary Fig. 3). Therefore, as reported before40,55, our in vitro assays confirm the preference for SUMO2/3 over SUMO1, with significantly slower reaction rates for SUMO1. This proteolytic behavior aligns with the SENP6 and SENP7 subfamily but diverges from the SENP1 and SENP2 subfamily, which are active toward both SUMO1 and SUMO2/3 isoforms in deconjugation reactions25,28,29,30,52.

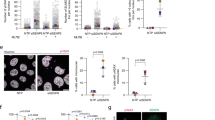
a SDS-PAGE of the endpoint assays of SENP5 using RanGAP1-SUMO1 and RanGAP1-SUMO2 substrates. (Right) Plot of the fraction of the RanGAP1-SUMO2 substrate after 30 min reaction. Data values represent the mean ± SEM, n = 3 technical replicates. Significance was measured by a two-tailed unpaired t test relative to wild-type. All data were analyzed with a 95% confidence interval. *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P ≤ 0.0001. Exact P value: 0.0003. b Plot of the proteolytic cleavage of SUMO1-AMC and SUMO2-AMC to test the activity of SENP5. SUMO2-AMC was incubated with SENP5, and released AMC was identified by fluorescence. c Plot of a competition assay by measuring the interference of the SENP5 activity for SUMO2-Rhodamine substrate by competition with either RanGAP1-SUMO1 and RanGAP1-SUMO2 at different concentrations. (Right) Inhibitory constant values for RanGAP1-SUMO1 and RanGAP-SUMO2 substrates on the SENP5 activity on SUMO2-Rho. Data values represent the mean ± SEM, n = 3 technical replicates. Source data are provided as a Source Data file.
To compare SENP5’s deconjugation activities between SUMO1 and SUMO2/3, we conducted a competitive in vitro fluorogenic kinetic assay (Fig. 1c). By measuring the decay of SENP5CD activity against SUMO2-Rhodamine upon incubation with increasing amounts of RanGAP1-SUMO2/3 or RanGAP1-SUMO1 substrates, we found that only RanGAP1-SUMO2/3 effectively competed with the fluorescent substrate (Fig. 1c). This competition resulted in an IC50 (half maximal inhibitory concentration) of approximately 3.7 µM. In contrast, RanGAP1-SUMO1 showed no competition with SUMO2-Rho deconjugation up to 30 µM concentration. These results further confirm SENP5’s significant preference for SUMO2/3 over SUMO1 in our in vitro assays.
Crystal structure of the human SENP5CD in complex with SUMO1 and SUMO2
To elucidate the determinants of SENP5’s preference for SUMO2, we engineered SUMO1 and SUMO2 with a propargylated C-terminal group (SUMO-PA). This modification allowed for covalent crosslinking of the SUMO C-terminus to the active site cysteine (Cys713) of SENP5CD, facilitating complex crystallization. SUMO1-PA was prepared using a synthetic peptide approach56, while SUMO2-PA was synthesized via an intein-based fusion protocol followed by propargylamine modification57,58.
SENP5CD complexes with SUMO1-PA and SUMO2-PA were formed via covalent crosslinking between the SUMO C-terminal alkyne group and the SENP5CD active site cysteine. After cationic exchange purification, both complexes were concentrated and prepared for crystallization experiments. Diffraction quality crystals were obtained in a buffer of 2.1 M DL-Malic acid pH 7.0 for SUMO1 complex and 0.1 M HEPES 7.0, 15% PEG 20,000 for SUMO2 complex. The SENP5CD-SUMO2 crystals belonged to P212121 space group and diffracted up to 2.36 Å resolution, whereas the SENP5CD-SUMO1 crystals belonged to P6522 space group and diffracted up to 2.18 Å resolution (Table 1). Electron density maps for both complexes revealed a continuous SENP5CD chain from Met568 to Asp756 and confirmed the covalent bond between SUMO Gly93 and SENP5CD Cys713, validating the specific reaction between SENP5CD and the SUMO-PA activity-based probes (Fig. 2).

a Cartoon representations of the SENP5CD-SUMO1 complex structure. SENP5 catalytic ___domain and SUMO1 are shown in orange and blue, respectively. The catalytic residues are labelled and depicted in stick representation. Secondary structure elements are labelled. N-terminal and C-terminal are marked. b Cartoon representations of the SENP5CD-SUMO2 complex structure. SENP5 catalytic ___domain and SUMO1 are shown in blue and green, respectively. The catalytic residues are labelled and depicted in stick representation. Secondary structure elements are labelled. N-terminal and C-terminal are marked. c Structural superimposition between SENP5CD-SUMO1 (blue) and SENP5CD-SUMO2 (green) complexes. SENP5CD is depicted as an electrostatic surface representation. d Structural alignment of sequences corresponding to the catalytic domains for human SENP2, SENP5 and SENP7. Red asterisks indicate interface residues with SUMO2. Catalytic triad residues are shown in red. Secondary structure cartoon is depicted above for SENP5 (green), SENP2 (violet) and SENP7 (brown). e Superposition of the SENP5CD structures (blue surface) in complex with SUMO1 (red) or SUMO2 (blue). Side chain interface residues are depicted in a stick representation.
The SENP5 catalytic ___domain adopts the canonical fold characteristic of the SENP/ULP family, albeit lacking certain N-terminal secondary structure elements found in other family members (Fig. 2d). Comparison of the bound and unbound SENP5 structures (AlphaFold-2 model) revealed minimal structural rearrangements upon SUMO-PA binding (rmsd values of 0.71 Å over 190 aligned residues). Structural overlapping shows that the active site catalytic triad might already be well arranged in the absence of SUMO substrate, as compared with the AlphaFold-2 structure of the unbound SENP5CD (Supplementary Fig. 2). The SENP5CD-SUMO2 interface is smaller than those observed in other SENP/ULP family members. PDBePISA59 analysis revealed an interface area of 833 Å2 for SENP5CD-SUMO2, compared to 982 Å2 and 1170 Å2 for SENP7CD (PDB ID 7R2E)52 and SENP2CD (PDB ID 2IO0)29, respectively (Supplementary Fig. 4).
Structural comparison indicates a closer evolutionary relationship between SENP5CD and the SENP1/SENP2 subgroup than the SENP6/SENP7 subgroup, as reflected by rmsd values of 1.22 Å over 175 aligned residues (with a 37% sequence identity with SENP2) and 1.77 Å over 167 aligned residues (with a 27% sequence identity with SENP7). Despite lacking the Loop1 insertion characteristic of SENP7, which is associated with SUMO2/3 specificity, SENP5CD exhibits a clear preference for SUMO2/3 over SUMO1. This suggests that distinct structural determinants within the SENP5CD interface may underlie its stronger deconjugation activity toward SUMO2/3 isoforms.
Structural comparison of SUMO1 and SUMO2 bound to SENP5CD revealed minimal differences in the docking interface (Fig. 2c). However, to understand the underlying reasons for SENP5’s lower isopeptidase activity toward SUMO1, a detailed analysis of specific interface contacts is necessary. While most contacts are conserved between SUMO1 and SUMO2, including key SENP/ULP family interface residues like Asp585, Phe609, and Trp625, a basic patch in the SENP5CD interface, consisting of Arg624 and Lys627, adopts different rotamer orientations (Fig. 2e). This patch participates in an electrostatic interaction only in the SUMO2 complex structure and resulted crucial for SENP5’s SUMO2 isopeptidase activity.
SENP5CD interface with SUMO1 and SUMO2
The C-terminal tails of SUMO1 and SUMO2 are buried within a SENP5CD cleft containing the catalytic triad, which cleaves the isopeptide bond following the C-terminal diGly motif (Figs. 2 and 3a). The geometry and contacts of the C-terminal tail in both complexes are similar to those observed in other SENP/ULP family members (Fig. 3b). The interface is primarily formed by a mainchain hydrogen bond network between the SUMO C-terminal tail (Q/E89-Q90-T91-G92-G93) and SENP5CD mainchain residues, along with interactions involving His642 (asparagine in SENP6 and 7) and the C-terminal SUMO diGly motif sandwiched between Trp582 (phenylalanine in SENP6 and 7) and Trp647. These C-terminal contacts are basically conserved in SENP2 and SENP7 interfaces with SUMO2 (PDB IDs 7R2E and 2IO0)29,52 (Fig. 3b).

a Close-up view of the C-terminal tail of human SUMO2 (yellow) in complex with SENP5CD (blue). Binding side-chain residues in the contact area of substrate and enzyme are shown in stick representation and labelled. Hydrogen bonds are represented by dashed red lines. b Similar close-up views of the C-terminal of SUMO2 (gray) in complex with SENP2 (upper) and SENP7 (below). c (Upper) Close-up view of the ribbon representation of SENP5CD-SUMO2 interface in two orthogonal views. Binding side-chain residues in the contact area are labelled and shown in stick representation. Hydrogen bonds and charged interactions are represented by dashed red lines. (Below) Similar close-up view of the ribbon representation of SENP5CD-SUMO1 interface in two orthogonal views.
SENP5’s preference for SUMO2 likely stems from interactions within the extensive interface between its catalytic ___domain and the SUMO globular ___domain. Highly conserved residues across the SENP/ULP family, including Asp585 (electrostatic with Arg59), Phe609 (stacking with Phe87), and Trp625 (contacting Gly64) (SENP5 and SUMO2 nomenclature), are crucial for both SUMO binding and catalytic activity (Fig. 3c). Interestingly, SENP8/NEDP1, the only SENP family member specific for Nedd8, deviates from this pattern with glutamate and proline substitutions for the conserved phenylalanine and tryptophan (Supplementary Fig. 1)60. While these contacts are observed in all SENP/ULP family members and are essential for the SUMO specificity over other UbLs, they do not alone determine SENP5’s SUMO2 preference25,28,29,30,52,53.
Among all SENP5CD interface contacts with SUMO1 and SUMO2, Asn607 and Arg624 stand out as potential determinants of SENP5’s SUMO2 preference. Asn607 forms a hydrogen bond with the carbonyl oxygen of Gly64 (SUMO2) or Gly68 (SUMO1), and in SUMO2 also with the guanidinium group of Arg61 (Figs. 3c and 4a). In SENP2 and SENP7, Asn607 is replaced by serine, preventing its involvement in the interface (PDB IDs 7R2E and 2IO0, respectively)29,52 (Fig. 2e).

a Close-up view of the ribbon representation of interfaces of SENP5CD-SUMO2 (green) and SENP5CD-SUMO1 (blue) around Asn607 and Arg624. Interface residues are labelled and shown in stick representation. b Plot of the proteolytic cleavage of SUMO1-AMC and SUMO2-AMC to test the activity of SENP5CD wild type and SENP5CD N607A, R624A, K627A, and R624/K627A mutants. SUMO2-AMC was incubated with SENP5CD, and released AMC was identified by fluorescence. The plot is an average of a triplicate experiment. c SDS-PAGE of the endpoint assays of SENP5CD wild type and SENP5CD N607A, R624A K627A, and R624/K627A mutants using RanGAP1-SUMO1 and RanGAP1-SUMO2 substrates. (Below) Plot of the fraction of the RanGAP1-SUMO2 and RanGAP1-SUMO1 substrate after 30 min reaction. Data values represent the mean ± SEM, n = 3 technical replicates. Significance was measured by a two-tailed unpaired t test relative to wild-type. All data were analyzed with a 95% confidence interval. *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P ≤ 0.0001. Exact P values from left to right: 0.0005, <0.0001, <0.0001, and <0.0001. d Western Blot analysis of HEK293F cells co-transfected with Flag-PML and either the empty pcDNA3 (−) or the plasmids expressing full-length SENP5, SENP5-R624A, SENP5-C713A or SENP5-R624A/K627A. (left) Immunoblot showing the levels of endogenous SUMO1 and SUMO2 in HEK293F cells. Anti-SUMO1 and antiSUMO2 antibodies were used to detect both endogenous SUMO isoforms. Anti-tubulin antibody was used as a loading control. (right) Plot of the Western Blot triplicate showing the fraction of polySUMO conjugates for each experiment regarding to SENP5 active site mutant (C713A). Data values represent the mean ± SEM, n = 3 technical replicates. Significance was measured by a two-tailed unpaired t test relative to C713A mutant. All data were analyzed with a 95% confidence interval. *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P ≤ 0.0001. Exact P values from left to right: <0.0001 and 0.0032. Source data are provided as a Source Data file.
Arg624, located within the α-helix α4, forms electrostatic interactions with Asp63 and Asp82 of SUMO2 in the SENP5CD-SUMO2 complex (bond distances: 2.8 Å and 3.2 Å, respectively) (Figs. 3, 4a and Supplementary Fig. 5). These interactions are absent in the SENP5CD-SUMO1 complex (Figs. 3, 4a and Supplementary Fig. 5). Arg624 is conserved in SENP1 and SENP2 but replaced by threonine in SENP6 and SENP7 (Fig. 2e and Supplementary Fig. 1). Interestingly, the presence of glutamic acid in SUMO1 (Glu67) or aspartic acid in SUMO2 (Asp63) at the same interface position may influence Arg624’s orientation, facilitating the electrostatic interaction in SUMO2. This difference could contribute significantly to SENP5’s preference for SUMO2.
Major role of Arg624 in the SENP5CD selectivity for SUMO2 substrates
In vitro activity assays using SENP5CD N607A, R624A, and K627A point mutants with both fluorogenic substrates (SUMO1-AMC and SUMO2-AMC) and model conjugated substrates (RanGAP1-SUMO1 and RanGAP1-SUMO2) revealed distinct effects on isopeptidase activity (Fig. 4b, c). While the N607A and K627A mutants impair activity for both SUMO1 and SUMO2 substrates, the R624A mutation only decreases the deconjugation activity for SUMO2 substrates. SENP5CD R624A exhibits a significant decrease in activity with all tested SUMO2 substrates but maintains comparable activity toward SUMO1 substrates compared to wild-type SENP5CD.
To further investigate SENP5’s deconjugation activity in a cellular context, we analyzed SUMO1 and SUMO2 deconjugation assays in HEK293F and COS-7 cells (Fig. 4d and Supplementary Fig. 6). HEK293F suspension cells were co-transfected with full-length SENP5 wild-type, C713A active site mutant, R624A single and R624A/K627A double point mutant, together with the Fl-PML substrate (Fig. 4d) and endogenous SUMO isoform conjugates were checked with either SUMO1 and SUMO2 antibodies. Western blot analysis of cellular extracts 4 days after transfection display a significant decrease of endogenous SUMO2 conjugates in SENP5 WT compared to SENP5 C713A, R624A, and R624A/K627A point mutants, supporting the role of Arg624 in the deconjugation activity. On the other hand, immunoblot analysis of similar extracts with SUMO1 antibody does not show significant differences between WT and the point mutants, probably indicating a poor activity of SENP5 for SUMO1-conjugates in HEK293F cells. Analogous results are observed in COS-7 cells co-transfected with HA-SUMO1 or HA-SUMO2, but in this instance we could only compared the different levels of unbound HA-SUMO2 (Supplementary Fig. 6).
All these activities suggest that the electrostatic interactions mediated by Arg624, particularly with Asp63 of SUMO2, plays a crucial role in enhancing SENP5’s preference for SUMO2 over SUMO1 conjugates.
Swapping the SUMO isoforms activities in SENP5CD
To further investigate the role of electrostatic interactions in SUMO isoform preference, we generated point mutations (SUMO1 E67D and SUMO2 D63E) that swapped the countercharge partner for Arg624 in SUMO1 and SUMO2 (Fig. 5). These mutations introduce a key chemical difference: the presence of an additional methylene group in the glutamate side chain compared to aspartate. We hypothesized that this alteration might contribute to the lower deconjugation activity observed with all tested SUMO1 substrates.

a SDS-PAGE of the endpoint assays of SENP5CD using the RanGAP1-SUMO1 and RanGAP1-SUMO2 wild types, and RanGAP1-SUMO1 E67D and RanGAP1-SUMO2 D63E point mutants as substrates. (Right) Plot of the fraction of the RanGAP1-SUMO1 wild type and point mutants after 30 min reaction. Data values represent the mean ± SEM, n = 3 technical replicates. Significance was measured by a two-tailed unpaired t test relative to wild-type. All data were analyzed with a 95% confidence interval. *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P ≤ 0.0001. Exact P values from left to right: 0.0001 and 0.0173. b Plot of the proteolytic cleavage of SUMO1-Rho, SUMO2-Rho wild types and SUMO1-Rho E67D and SUMO2-Rho D63E point mutants to test the activity of SENP5CD. The plot is an average of a triplicate experiment. c Plot of a competition assay by measuring the interference of the SENP5CD activity for SUMO2-Rhodamine substrate by competition with RanGAP1-SUMO1 and RanGAP1-SUMO2 wild types, and RanGAP1-SUMO1 E67D and RanGAP1-SUMO2 D63E point mutants at different concentrations. (Right) Inhibitory constant values (IC50) for RanGAP1-SUMO1 and RanGAP1-SUMO2 wild types, and RanGAP1-SUMO1 E67D and RanGAP1-SUMO2 D63E point mutants substrates on the SENP5CD wild type activity on SUMO2-Rhodamime fluorogenic substrate. Data values represent the mean ± SEM, n = 3 technical replicates. d Immunoblot analysis of the labeling of endogenous full length human SENP3 in HEK293T lysates with activity-based probes of either SUMO1 or SUMO2 wild type and point mutants. Anti-actin antibody was used as a loading control. n = 3 technical replicates. e Close-up view of the ribbon representation of interfaces of SENP5CD-SUMO1 wild type depicting the basic patch region around Arg624. Interface residues are labelled and shown in stick representation. f Close-up view of the ribbon representation of interfaces of SENP5 CD-SUMO1 E67D point mutant depicting the basic patch region around Arg624. g Superposition of the SENP5 structures (blue surface) in complex with SUMO1 wild type (blue ribbon) or SUMO1 E67D (orange ribbon). Source data are provided as a Source Data file.
Strikingly, the SUMO1 E67D mutation enhances its deconjugation activity toward RanGAP1 conjugates compared to wild-type SUMO1, which is nearly inactive. Conversely, the SUMO2 D63E mutation significantly reduces RanGAP1 deconjugation activity (Fig. 5a). Fluorogenic SUMO-Rhodamine confirmed these results, displaying similar activities for SUMO2-Rho D63E and SUMO1-Rho E67D, with a notable gain of activity for SUMO1 mutant compared to the loss for SUMO2 mutant (Fig. 5b).
Following our initial competitive kinetics assay (Fig. 1c), we investigated the impact of these swapping mutations using a similar approach. We measured the decrease in SUMO2-Rho activity upon competition with RanGAP1 conjugated to either SUMO1 E67D or SUMO2 D63E (Fig. 5c). The IC50 for RG-SUMO2 D63E significantly increased from 3.7 µM to 56.4 µM compared to wild-type SUMO2, indicating a roughly 15-fold decrease in its inhibitory activity. Conversely, the IC50 for RG-SUMO1 E67D, which was undetectable for wild-type SUMO1, became measurable at around 87.2 µM, quite similar to RG-SUMO2 D63E.
Human SENP3, the closest homolog of SENP5, has been suggested to share functional overlap50,59. Consistent with SENP5 results, full length SENP3 preferentially binds SUMO2-derived activity-based probe (ABP) (Fig. 5d). While SENP3 showed a slight binding increase against SUMO1 E67D, it exhibited decreased binding against SUMO2 D63E compared to their wild-type counterparts. These findings reinforce the importance of Arg624 in mediating SUMO2/3 selectivity for both SENP3 and SENP5, thus broadening the significance of our results.
The crystal structure of SUMO1E67D resembles the SUMO2 interface to SENP5CD
Moreover, to further investigate the molecular mechanism behind the SUMO1 E67D enhancement, we successfully crystallized and solved the structure of SENP5CD in complex with the SUMO1 E67D mutant at 1.73 Å resolution (Table 1) (Fig. 5e, f). Electron density maps of the complex interface validated our hypothesis regarding the role of Arg624, revealing the presence of electrostatic contacts between Arg624 and Lys627 with Asp67 in the SUMO1 mutant, resembling those observed in the SENP5CD-SUMO2 complex (Suppementary Figs. 5 and 7). However, these interactions were absent in wild-type SUMO1, where Glu67 is present. Furthermore, the overall structural alignment of wild-type and E67D SUMO1 revealed a significant backbone shift, positioning the mutant closer to the SUMO2 interface (Fig. 5g).
Our findings suggest that the electrostatic interaction between Arg624 in SENP5CD and Asp63 in SUMO2 contributes to SENP5’s higher proteolytic activity toward SUMO2 conjugates. The shorter side chain of aspartate compared to glutamate in SUMO1, may allow for the optimal positioning and distance required for a stronger electrostatic interaction with arginine, potentially enhancing SUMO2 binding and subsequent proteolytic processing by SENP5CD.
Discussion
The human SENP/ULP family of SUMO isopeptidases can be classified into three subgroups based on structural and sequence analysis: SENP1-2, SENP3-5, and SENP6-7. Excluding the divergent SENP8/NEDP1, which is specific for Nedd861, all other SENP members exhibit SUMO isopeptidase activity29,30,52,54. While SENP1 and SENP2 exhibit activity toward both SUMO isoforms, the remaining subgroups, SENP3-5 and SENP6-7, demonstrate a strong preference for the SUMO2 isoform as determined by in vitro deconjugation assays. Within the SENP6-7 subgroup, long insertions are present in the middle of the catalytic ___domain, including the “Loop1” insertion, which we identified as a key determinant of SUMO2/3 substrate specificity25,52,53,54. In contrast, the specificity of SENP3 and SENP5 for SUMO2/3 isoforms likely arises from interactions other than “Loop1”, which they lack. To elucidate the structural basis for SUMO2 isoform preference in SENP5, we have compared the crystal structures of SENP5CD in complex with either SUMO1 or SUMO2.
Full-length SENP/ULP family members consist of a long, structurally disordered N-terminal extension followed by a conserved catalytic ___domain. The catalytic ___domain adopts a characteristic CE-clan cysteine protease fold18. While the precise function of the disordered N-terminal extensions remains to be fully elucidated, available evidence suggests that they may play roles in several aspects of SENP/ULP biology24,31,32,33,34. These extensions could potentially contribute to the subcellular localization of SENP/ULP members or facilitate the binding of specific protein targets within the cell.
Cellular functions attributed to the non-catalytic disordered regions in substrate recruitment have been reported for SENP6 and SENP7. For instance, the presence of seven SIM (SUMO-Interacting motif) domains within the disordered region of SENP7 has been shown to play a crucial role in the recruitment of polySUMOylated KAP1 (Karyopherin 1) during homologous recombination62. The disordered region of SENP6 also contains multiple SIM motifs, enabling it to recruit polySUMOylated targets for deconjugation24,63,64. Additionally, the yeast ULP2, a SENP6 homolog, possesses multiple SIM motifs in its N-terminal disordered region24. This suggests a conserved role for SIM motifs in the recruitment of polySUMOylated substrates by SENP/ULP family members65. In the SENP3 and SENP5 subfamily, the non-structured N-terminal region has primarily been implicated in subcellular localization40,46,51,55. Despite their length and structural disorder, the N-terminal extensions of all SENP/ULP family members appear to be dispensable for catalytic activity, including the ability to dismantle polySUMO2 chains. The catalytic activity of these enzymes is solely confined to their conserved CE-clan catalytic domains.
Structural analysis of SENP5CD complexes with SUMO1 and SUMO2 reveals a smaller interface with SUMO compared to SENP1CD, SENP2CD, and SENP7CD (Supplementary Fig. 4). Despite its reduced size, this interface is sufficient to confer SUMO2/3 specificity over SUMO1. In addition to the common SUMO contacts observed in all members of the SENP/ULP family, such as the conserved C-terminal tail and essential residues in the globular ___domain (Asp585, Phe609, and Trp625), the SENP5CD interface features unique contacts that may contribute to its preference SUMO2/3. One such feature is a basic patch centered around Arg624.
The relatively smaller interface between SENP5CD and SUMO2 compared to SENP1CD, SENP2CD, or SENP7CD may amplify the effects of single point mutations on deconjugation activity, as observed in our analysis of the SENP5CD N607A, K627A, and R624A point mutants. These positions were chosen, particularly N607A and R624A, because they engage in unique interactions with SUMO2, compared to SUMO1. Surprisingly, whereas disruption of Asn607 decreases the SENP5CD activity for both SUMO isoforms, Arg624 has a strong decrease of deconjugating activity only for SUMO2 substrates. Arg624 in SENP5CD forms a well-oriented salt bridge interaction with Asp63 and Glu77 in SUMO2. However, in SUMO1, the equivalent Asp63 position is replaced by a glutamate residue. Due to its longer side chain, glutamate is unable to properly accommodate Arg624, preventing the formation of the electrostatic interaction observed in complex with SUMO2, but absent in the SUMO1 complex.
Interestingly, a similar electrostatic interaction between Arg624 and SUMO2 residues can be observed in the SENP2CD-SUMO2 complex structure (PDB code 2IO0)29. However, this interaction is absent in the SENP2CD-SUMO1 complex (PDB code 1TGZ)28, where the arginine side chain is oriented outwards, resembling the conformation seen in our SENP5CD-SUMO1 complex. Despite the presence of this electrostatic interaction in SENP2CD-SUMO2, SENP2 does not exhibit a strong preference for either SUMO1 or SUMO2. This suggests that the larger number of interface contacts in SENP2CD-SUMO complexes may compensate for the absence of the arginine-SUMO1 interaction, mitigating its effect on isoform specificity.
In addition to their well-known roles in eukaryotic systems, CE-clan proteases, such as SENP5, have been identified in infectious bacteria and viruses, raising intriguing questions about their evolution and function. Given that most prokaryotes lack conventional ubiquitin or UBLs, the presence of bacterial CE-clan proteases targeting ubiquitin conjugates is particularly notable66,67,68. This adaptation expands the substrate specificity of the canonical catalytic ___domain, enabling bacteria to manipulate the host’s ubiquitin and UBL signaling pathways. Such modifications likely enhance bacterial survival and persistence within the host. In bacterial CE-clan proteases, specificity for ubiquitin and UBLs is mainly regulated by changes in three connecting loops (VR1-3) at the interface with the ubiquitin or UBL moiety67. In contrast, SENP5’s specificity for SUMO2 is primarily determined by a basic patch surrounding Arg624, located in the VR-2 loop, while SENP7 recognizes SUMO2 through the Loop1 insertion in the VR-1 loop. These differences illustrate the remarkable adaptability of CE-clan proteases and their diverse strategies for achieving substrate specificity.
The human SENP family plays a pivotal role in regulating a variety of cellular processes, and their dysregulation is implicated in numerous diseases, including cancer, neurodegenerative disorders, and cardiovascular diseases. Recent reviews have emphasized the critical involvement of SENP proteases in tumorigenesis, highlighting their role in the disruption of cellular signaling pathways8,69. Given the importance of SENPs in cellular regulation, understanding the molecular mechanisms behind their substrate specificity, particularly the preference for SUMO isoforms, is essential. This insight could facilitate the development of highly specific inhibitors and probes targeting individual SENP family members, offering new therapeutic opportunities with minimal off-target effects.
Methods
Plasmids, cloning and point mutation
SENP5CD catalytic ___domain (aa 568–755) was amplified from human cDNA library and cloned into a pET28a vector. Restriction Enzyme Free PCR was used to clone the PTXB1-Δ14-humanSUMO2-intein-chitin-binding ___domain (CBD) (14 residues N-terminal deletion)70. Point mutants of SENP5, SUMO1, and SUMO2 were performed by site-directed mutagenesis71. Full-length RGS-SENP5 was a gift from Edward Yeh (Addgene plasmid #18053; http://n2t.net/addgene:18053; RRID:Addgene_18053), pLPC-Flag-PML-IV was a gift from Gerardo Ferbeyre (Addgene plasmid #62804; http://n2t.net/addgene:62804; RRID:Addgene_62804), pcDNA3-HA-SUMO1 was a gift from Junying Yuan (Addgene plasmid #21154; http://n2t.net/addgene:21154; RRID:Addgene_21154) and pcDNA3 HA-SUMO2 WT was a gift from Guy Salvesen (Addgene plasmid #48967; http://n2t.net/addgene:48967; RRID:Addgene_48967)38,72,73,74. The primers used for these experiments are shown in Supplementary Table 1. Final constructs were verified by DNA sequencing.
Protein expression and purification
SENP5CD (aa 568 to 755) and ∆14SUMO2 expression constructs were expressed in E. coli Rosetta (DE3) cells (Novagen) for 5 h at 30 °C after induction with 0.5 mM IPTG. SENP5CD suspension was equilibrated in 350 mM NaCl, 20 mM Tris-HCl (pH 8.0), 10 mM imidazole, 20% sucrose, 1 mM BME (β-mercaptoethanol), and 0.1% IGEPAL CA-630, and cells were sonicated to break. Nickel affinity chromatography was used to purify SENP5CD via the N-terminal HIS-tag from the pET28 vector. Proteins were further separated by gel filtration chromatography (Superdex 75 26/60; Cytiva) with buffer 250 mM NaCl, 20 mM Tris 8.0, 1 mM BME.
For ∆14-SUMO2 PA protein purification, 60 mL chitin beads were prepared (New England Biolabs, UK), previously washed with 5CV (column volume) (1CV = 200 mL) cold lysis buffer (50 mM HEPES 8.0, 50 mM NaCl) in 4 degrees. Four liters of Δ14SUMO2-intein-CBD pellet are resuspended in 200 mL lysis buffer. After being sonicated, the 200 mL supernatant is filtered through a 0.45 μm nylon membrane filter, placed into the chitin column and gently shaken overnight at 4 °C. To remove unbound proteins, the column was washed at 4 °C with 5CV lysis buffer. Then 200 mL of lysis buffer containing 150 mM MesNa was added and gently shaken for 48 h at room temperature. The remaining product was collected by adding 15 mL lysis buffer to the 200 mL flow-through. Eluted sample was concentrated to 30 mL with a 3 kDa filter and dialyzed overnight at 4 °C. After dialysis, 150 mM Propargylamine was added to the 30 mL product for 4 and 5 h at room temperature. The product was dialyzed overnight at 4 °C, concentrated to remove precipitate and applied to an anion exchange resin (Resource Q 6 mL; GE Healthcare). Elution was performed with a 0–1 M NaCl gradient from 0 to 50% in 20 mM Tris-HCl (pH 8.0) and 1 mM BME.
Synthesis of Δ18 N SUMO1 wild-type and E67D PA
SUMO1 E67D (ΔN; 18 to C-term) was obtained by standard 9-fluorenylmethoxycarbonyl (Fmoc) based solid phase peptide chemistry on trityl-resin as described in literature on a 20 mmol scale56. The N-terminus was protected by treating the resin with Di-tert-butyl dicarbonate (21.8 mg, 100 µmol, 5.0 eq.) and DIPEA (34.84 µl, 200 µmol, 10.0 eq.) in DMF (2 ml) for 3 h. Sidechain protected SUMO was released from the resin by treatment with 1,1,1,3,3,3-hexafluoropropan-2-ol (HFIP)/DCM (2.5 ml, 1/4, v/v) for 3 × 20 min, after which it was filtered and the solvent was removed by rotary evaporation. Residual HFIP was removed by co-evaporation 2–3 times with DCE (to prevent formation of HFIP ester in next steps) and dried to use in the next step. The SUMO protein was dissolved in DCM (5 mL) and to this were added propargyl amine (13 μL, 200 μmol, 10 eq), PyBOP (52 mg, 100 μmol, 5.0 eq) and TEA (14 μL, 100 μmol, 5.0 eq), upon which the mixture was stirred at rt for 16 h. The mixture was concentrated under reduced pressure and the SUMO polypeptide sequence was dissolved in a mixture of TFA/H2O/TIPS/phenol (90.5/5/2/2.5 v/v/v/v; 5 mL) and stirred for 4 h at rt towards global deprotection. The protein was precipitated with ice-cold Et2O/n-pentane (3/1; v/v; 40 mL). The precipitated protein was isolated by centrifugation and washed by three cycles of resuspension in ice-cold diethyl ether and centrifugation. The pellet was dissolved in H2O/MeCN/FA (75/24/1; v/v/v; 20 mL) and lyophilized. The crude product was purified using RP-HPLC and relevant fractions were pooled and lyophilized to afford ΔN SUMO1 E67D PA as a white solid (Supplementary Fig. 8).
Synthesis of SUMO-Rhodamine-Morpholine (RM) reagents
SUMO1/2 wild-type or mutant (SUMO1 E67D and SUMO2 D63E) were obtained by standard 9-fluorenylmethoxycarbonyl (Fmoc) based solid phase peptide chemistry on trityl-resin as described in literature on a 20 mmol scale56. The N-terminus was protected by treating the resin with Di-tert-butyl dicarbonate (21.8 mg, 100 µmol, 5.0 eq.) and DIPEA (34.84 µl, 200 µmol, 10.0 eq.) in DMF (2 ml) for 3 h. Sidechain protected SUMO was released from the resin by treatment with 1,1,1,3,3,3-hexafluoropropan-2-ol (HFIP)/DCM (2.5 ml, 1/4, v/v) for 3 × 20 min, after which it was filtered and the solvent removed by rotary evaporation. Residual HFIP was removed by co-evaporation 2–3 times with DCE (to prevent formation of HFIP ester in next steps) and dried to use in the next step. The SUMO protein was dissolved in DMF (5 mL) and to this were added EDC HCl (18 mg, 10 μmol, 5.0 eq), HOBt monohydrate (15 mg, 100 μmol, 5.0 eq) and GlyRM75 (20 mg, 40 μmol, 2.0 eq), upon which the mixture was stirred at rt for 16 h. The mixture was concentrated under reduced pressure and the SUMO polypeptide sequence was dissolved in a mixture of TFA/H2O/TIPS/phenol (90.5/5/2/2.5 v/v/v/v; 5 mL) and stirred for 4 h at rt towards global deprotection. The protein was precipitated with ice-cold Et2O/n-pentane (3/1; v/v; 40 mL). The precipitated protein was isolated by centrifugation and washed by three cycles of resuspension in ice-cold diethyl ether and centrifugation. The pellet was dissolved in H2O/MeCN/FA (75/24/1; v/v/v; 20 mL) and lyophilized. The purity of the peptides was determined by LC-MS analysis. The crude product was purified using RP-HPLC and relevant fractions were pooled and lyophilized to afford the SUMO peptides as white solids (Supplementary Figs. 9 and 10).
Synthesis of SUMO propargylamide activity-based probes
SUMO1/2 wild type or mutant (S1 E67D, S2 D63E) were obtained by standard 9-fluorenylmethoxycarbonyl (Fmoc) based solid phase peptide chemistry on trityl-resin as described in literature on a 20 mmol scale. A solution of Fmoc-PEG2-COOH (39 mg, 100 μmol, 5.0 eq), PyBOP (52 mg, 100 μmol, 5.0 eq), and DIPEA (36 μL, 200 μmol, 10 eq) in NMP was shaken for 10 min and added to the SUMO on resin for 3 h at room temperature. After completion, the resin was washed alternately with NMP (3 × 5 mL) and DCM (3 × 5 mL). The resin was treated with piperidine/NMP (4 mL, 1/4, v/v) for 2 × 20 min. A solution of Fmoc-Lys(biotinyl)-OH (59 mg, 100 μmol, 5.0 eq), PyBOP (52 mg, 100 μmol, 5.0 eq) and DIPEA (36 μL, 200 μmol, 10 eq) in NMP was shaken for 10 min and added to the SUMO on resin for 3 h at room temperature. After completion, the resin was washed alternately with NMP (3 × 5 mL) and DCM (3 × 5 mL). The resin was treated with piperidine/NMP (4 mL, 1/4, v/v) for 2 × 20 min. A solution of Boc2Rho (34 mg, 60 μmol, 3.0 eq), PyBOP (52 mg, 100 μmol, 5.0 eq), and DIPEA (36 μL, 200 μmol, 10 eq) in NMP was shaken for 10 min and added to the SUMO on resin for 3 h at room temperature. After completion, sidechain protected Rho/Bio-SUMO dG was released from the resin by treatment with 1,1,1,3,3,3-hexafluoropropan-2-ol (HFIP)/DCM (2.5 ml, 1/4, v/v) for 3 × 20 min, after which it was filtered and the solvent was removed by rotary evaporation. Residual HFIP was removed by co-evaporation 2–3 times with DCE (to prevent formation of HFIP ester in next steps) and dried to use in the next step. The SUMO protein was dissolved in DMF (5 mL) and to this were added propargyl amine (6.4 μL, 100 μmol, 5.0 eq), EDC HCl (19.2 mg, 100 μmol, 5.0 eq) and HOBt (15.2 mg, 100 μmol, 5.0 eq), upon which the mixture was stirred at rt for 16 h. The mixture was concentrated under reduced pressure and the SUMO polypeptide sequence was dissolved in a mixture of TFA/H2O/TIPS/phenol (90.5/5/2/2.5 v/v/v/v; 5 mL) and stirred for 4 h at rt towards global deprotection. The protein was precipitated with ice-cold Et2O/n-pentane (3/1; v/v; 40 mL The precipitated protein was isolated by centrifugation and washed by three cycles of resuspension in ice-cold diethyl ether and centrifugation. The pellet was dissolved in H2O/MeCN/FA (75/24/1; v/v/v; 20 mL) and lyophilized. The protein was purified using RP-HPLC to afford the Rho/Bio-SUMO PA probes as pink solids (Supplementary Figs. 11 and 12).
Generation of SENP5-SUMO complexes
SENP5 Δ568 protein was incubated with ∆14SUMO2-PA, ∆18SUMO1-PA or ∆18SUMO1E67D-PA (1:3 ratio) in a buffer containing 250 mM NaCl, 20 mM Tris 8.0, 3 mM DTT for 2 h at 37 °C. The protein buffer was exchanged to 50 mM NaCl, 20 mM Tris 8.0, 1 mM DTT and applied to an cation exchange resin (Resource S 6 mL; GE Healthcare).
Crystallization, data collection and refinement
The complexes of SENP5CD with ∆14SUMO2 PA, ∆18SUMO1 PA or ∆18SUMO1E67D PA were finally concentrated to 9.5 mg/mL, 3.5 mg/mL and 6.6 mg/mL, respectively for crystallization in a buffer composed by 60 mM NaCl, 8 mM Tris 8.0, 1 mM BME. Crystallization was performed at 18 °C by sitting drop vapor diffusion method by mixing protein with an equal volume of a screen condition solution containing 0.1 M Sodium HEPES 7.0, 15 % w/v PEG 20,000 for SUMO2, 2.1 M DL-Malic acid pH 7.0 for SUMO1 or 0.2 M Lithium sulfate, 0.1 M Sodium acetate 5.5, 8 % w/v PEG 20,000 and 8 % v/v PEG 500 MME for SUMO1E67D. After 1 week, the crystals were harvested and soaked for 10 s in 15% ethylene glycol before flash freezing in liquid nitrogen.
Diffraction data were collected to 2.36 Å, 1.73 Å, and 2.18 Å resolution for SUMO1, SUMO1E67D, and SUMO2 complexes, respectively, at the ALBA synchrotron beamline BL13-XALOC (Barcelona, Spain)76 (Table 1). Data was processed, scaled, reduced, and further analyzed with the automatic autoPROC software package77 including XDS, POINTLESS, AIMLESS, CCP4, and STARANISO. Crystallographic details are summarized in Table 1.
The structure of SENP5CD-∆14SUMO2, SENP5CD-∆18SUMO1, and SENP5CD-∆18SUMO1E67D was solved by molecular replacement using SENP5 AlphaFold-2 model and SUMO2 (1WM3) or SUMO1 (1TGZ) as search models with Phaser. Following rounds of model building and refinement were carried out with Coot and Phenix78,79 (Table 1).
Transient cell transfection
HEK293F cells were maintained at 37 °C in FreeStyle™ 293 Expression Medium (Gibco, Life Technologies). Cells were seeded at a density of 0.5 × 106 cells/mL in a final volume of 2 mL per well in a 6-well plate. The cells were incubated for 24 h in an orbital shaker incubator at 37 °C, and a humidified atmosphere with 8% CO2 until they reached a density of 1.0 × 106 cells/mL. Subsequently, the cells were co-transfected with 1 µg of DNA per 1 mL of final cell culture using a 1:3 (w/w) ratio of DNA to polyethylenimine (PEI linear, 25,000 Da, Polysciences, Warrington, PA, USA). The cells were then incubated in an orbital shaker for 96 h at 37 °C, 120 rpm, and 8% CO2. After 96 h, the cells were harvested by centrifugation at 3000 × g for 5 min. Lysis was performed using M-PER® Mammalian Protein Extraction Reagent (Thermo Scientific) according to the manufacturer’s instructions. N-Ethylmaleimide was added at a final concentration of 10 µM during lysis. Protein quantification was performed using the Bradford Protein Assay. To perform western blot analysis, 30 µg of protein were separated by SDS/PAGE in a NuPAGETM 4–12% Bis-Tris gel (NP0323BOX, Invitrogen) with NuPAGETM MOPS SDS running buffer (NP0001, Invitrogen).
COS-7 cells were maintained at 37 °C in Dulbecco’s Eagle’s medium (Sigma-Aldrich, BE12-604F) supplemented with 10% (v/v) of fetal bovine serum (631106, Clontech) and 1% (v/v) penicillin/streptomycin (Invitrogen, 15140-122). COS-7 cells were cultured at 70% confluency and co-transfected with 16 μg of DNA per 10 cm-Petri dish, in a ratio of 1:3. Lipotransfectin reagent (Niborlab) was used according to the manufacturer’s instructions. Green Fluorescent Protein expression construct was used as a positive control for transfection. 48 h after transfection, cells were lysed in 1 mL per 10 cm-petri dish of Pierce IP Lysis Buffer (Thermo Scientific, 87787) supplemented with Halt Protease Inhibitor Cocktail (Thermo Scientific, 78438). Lysates were incubated on ice for 20 min and clarified by centrifugation at 3000 × g for 15 min at 4 °C to precipitate proteins. Bradford Protein Assay was used to quantify protein in the spectrophotometer Sinergy HT and analyzed using MultiGauge Software (Fujifilm). To detect the mono-HA-SUMO-2 form, 50 μg of protein were separated by SDS/PAGE in a 12% acrylamide gel.
Western blot analysis
SDS/PAGE, proteins were transferred for 2 h on to PVDF membranes (Millipore), which were blocked with 5% (v/v) non-fat milk in TBS-T [TBS containing 0.1% (v/v) Tween20] for 1 h. Membranes were incubated overnight at 4 °C with the primary antibody (see below) in TBS-T containing 5% (v/v) non-fat milk, washed three times with TBS-T, and incubated for a further 1 h in TBS-T containing 5% (v/v) non-fat milk containing the appropriate secondary antibody. After three washes with TBS-T, membranes were processed with Immobilon® Forte Western Blotting HRP Substrate (Merck Millipore, WBLUF0100) in Li-Cor Odyssey XF imager. Primary antibodies used in Western blotting were anti-HA tag antibody—ChIP Grade (Abcam, ab9110, 1:2000) and anti-alpha Tubulin antibody [DM1A] (Abcam, ab7291, 1:1000), anti-SUMO1 (C-terminal) (S5446, Sigma-Aldrich 1:2000), anti-SUMO2 (S9571, Sigma-Aldrich 1:2000), HRP-conjugated Mouse Anti-Rabbit antibody (light chain-specific) (Cell Signaling Technology, 93702, 1:1000), polyclonal Rabbit anti-mouse immunoglobulin HRP (Dako, P0260, 1:5000).
SUMO-AMC hydrolysis assays
SENP5 wild type and mutants were incubated with SUMO1-AMC (BML-UW0040-0050, ENZO Life Sciences) and SUMO2-AMC (BML-UW0045-0025, ENZO Life Sciences) at 37 °C and fluorescence emission was measured using a Jasco FP-8200 spectrofluorometer at 345 nm excitation and 445 nm emission wavelengths. All measurements were carried out with 50 nM SENP5 and 0.1 μM Ub-AMC in 250 mM NaCl, 20 mM Tris-HCl pH 8, 4 mM DTT buffer.
SUMO-Rhodamine Morpholine hydrolysis assays
The wild-type SENP5 catalytic ___domain was incubated with labeled SUMO substrates, including WTSUMO1, E67DSUMO1, WTSUMO2, and D63ESUMO2, conjugated to SUMO-Rhodamine Morpholine (prepared in Monique Mulder’s lab). Reactions were carried out at 37 °C, and fluorescence emission was measured using a Jasco FP-8200 spectrofluorometer, with excitation at 485 nm and emission at 520 nm. For all measurements, 50 nM SENP5 and 0.5 μM SUMO-Rho were used in a buffer consisting of 250 mM NaCl, 20 mM Tris-HCl (pH 8.0), and 4 mM DTT.
For RANGAP1-SUMO competition assays, the methodology was similar, except that WTSUMO2 was labeled with Rhodamine 110 (purchased from South Bay Bio LLC, SBB-PS0029), with excitation at 485 nm and emission at 535 nm. Increasing concentrations of unlabeled NΔ419RANGAP1-SUMO1, NΔ419RANGAP1-SUMO1E67D, NΔ419RANGAP1-SUMO2, or NΔ419RANGAP1-SUMO2D63E were added to the reaction mixture. An increase in fluorescence signal indicates proteolytic hydrolysis of the amide bond between the C-terminal diGly motif and the rhodamine compound. Conversely, the fluorescence emission rate should decrease in the presence of competitive substrates. The initial SENP5CD activity (initial velocity of fluorescence emission, slope) was set to 100% in the absence of competitive substrates, and the remaining SENP5CD activity (%) was measured with increasing concentrations of the competitive substrates.
In vitro de-SUMOyation assays
SENP5CD protease activity was determined by incubating NΔ419RanGAP1-SUMO1 WT/E67D, NΔ419RanGAP1-SUMO2WT/D63E (1 μM), preSUMO1WT/E67D, preSUMO2WT/D63E (2 μM) or diSUMO2 (1 μM) with 5 nM or 150 nM of SENP5 wild type and mutants at 37 °C in a buffer containing 20 mM Tris-HCl (pH 8.0), 250 mM NaCl, and 2 mM DTT. Reactions were stopped with SDS-BME loading buffer and gel electrophoresis (SDS-PAGE) was performed. Gels were stained by SYPRO (Bio-Rad), and proteins were detected and quantified using a Gel-Doc machine with associated integration software (ImageLab; Bio-Rad). The fraction of analyzed bands was plotted as error bar graphs with standard deviations representing the mean ± SEM.
Labeling of endogenous SENP3 in cell lysates
HEK293T cells were harvested in lysis buffer (150 mM NaCl, 50 mM Tris pH 7.5, 0.5% Triton-X100, Protease inhibitor cocktail (Roche, 11873580001)), and collected after sonication (30 s on/off, 5 cycles, 4 °C) and centrifugation (14000 rpm, 4 °C, 10 min). Lysate (40 μL) was incubated with Rho/Bio-SUMO PA probe (40 μL, 1 μM final concentration) for 5 min at 37 °C. Reactions were stopped by the addition of LDS sample buffer containing 10% BME, followed by boiling for 10 min. Samples were resolved by SDS-PAGE, followed by in-gel fluorescence scanning and transfer to nitrocellulose membranes. Membranes were blocked in 5% milk in PBS-T followed by 1 h incubation at room temperature of primary antibody: rabbit anti-SENP3 (1:1000 dilution; Cell Signalling Technology; 5591) and mouse anti-β-actin (1:10000 dilution; Santa Cruz Biotechnology.; sc-47778). This was followed by 1 h incubation at room temperature of fluorescently labelled secondary antibody: anti-rabbit-800 (1:20000 dilution; LiCOR; 926-68021) and anti-mouse-800 (1:20000 dilution; LiCOR; 926-68020). Labelled SENP3 was visualized on a LICOR Odyssey system v3.0.
Data availability
Structures reported here have been deposited in the Protein Data Bank under accession codes 9GNN, 9GNV, and 9GNX. Other Protein Data Bank accession codes used in this study: 2IO0 (SENP2-SUMO2) and 7R2E (SENP7-SUMO2). All other data supporting the findings of this study are available within the article and its supplementary information files. Source data are provided with this paper.
References
Varejão, N. et al. Molecular mechanisms in SUMO conjugation. Biochem. Soc. Trans. 48, 123–135 (2020).
Bhachoo, J. S. & Garvin, A. J. SUMO and the DNA damage response. Biochem. Soc. Trans. 52, 773–792 (2024).
Vertegaal, A. C. O. Signalling mechanisms and cellular functions of SUMO. Nat. Rev. Mol. Cell Biol. 23, 715–731 (2022).
Wu, W. & Huang, C. SUMOylation and DeSUMOylation: prospective therapeutic targets in cancer. Life Sci. 332, 122085 (2023).
Lascorz, J. et al. SUMO-SIM interactions: from structure to biological functions. Semin Cell Dev. Biol. 132, 193–202 (2022).
Chang, H.-M. & Yeh, E. T. H. SUMO: from bench to bedside. Physiol. Rev. 100, 1599–1619 (2020).
Yau, T.-Y. et al. SUMO interacting motifs: structure and function. Cells 10, 2825 (2021).
Claessens, L. A. & Vertegaal, A. C. O. SUMO proteases: from cellular functions to disease. Trends Cell Biol. https://doi.org/10.1016/j.tcb.2024.01.002 (2024).
Kunz, K. et al. SUMO-specific proteases and isopeptidases of the SENP family at a glance. J. Cell Sci. 131, jcs211904 (2018).
Hickey, C. M. et al. Function and regulation of SUMO proteases. Nat. Rev. Mol. Cell Biol. 13, 755–766 (2012).
Yeh, E. T. H. SUMOylation and De-SUMOylation: Wrestling with Life’s Processes *. J. Biol. Chem. 284, 8223–8227 (2009).
Bouchard, D. et al. SUMO paralogue-specific functions revealed through systematic analysis of human knockout cell lines and gene expression data. Mol. Biol. Cell 32, 1849–1866 (2021).
Wang, W. & Matunis, M. J. Paralogue-specific roles of SUMO1 and SUMO2/3 in protein quality control and associated diseases. Cells 13, 8 (2024).
Owerbach, D. et al. A proline-90 residue unique to SUMO-4 prevents maturation and sumoylation. Biochem. Biophys. Res. Commun. 337, 517–520 (2005).
Drag, M. & Salvesen, G. S. DeSUMOylating enzymes-SENPs. IUBMB Life 60, 734–742 (2008).
Nayak, A. & Müller, S. SUMO-specific proteases/isopeptidases: SENPs and beyond. Genome Biol. 15, 422 (2014).
Mukhopadhyay, D. & Dasso, M. Modification in reverse: the SUMO proteases. Trends Biochem Sci. 32, 286–295 (2007).
Barrett, A. J. & Rawlings, N. D. Evolutionary lines of cysteine peptidases. Biol. Chem. 382, 727–733 (2001).
Mossessova, E. & Lima, C. D. Ulp1-SUMO crystal structure and genetic analysis reveal conserved interactions and a regulatory element essential for cell growth in yeast. Mol. Cell 5, 865–876 (2000).
Li, S. J. & Hochstrasser, M. A new protease required for cell-cycle progression in yeast. Nature 398, 246–251 (1999).
Suh, H.-Y. et al. Crystal structure of DeSI-1, a novel deSUMOylase belonging to a putative isopeptidase superfamily. Proteins 80, 2099–2104 (2012).
Li, Y. et al. Structural basis for the SUMO protease activity of the atypical ubiquitin-specific protease USPL1. Nat. Commun. 13, 1819 (2022).
Schulz, S. et al. Ubiquitin-specific protease-like 1 (USPL1) is a SUMO isopeptidase with essential, non-catalytic functions. EMBO Rep. 13, 930–938 (2012).
Wagner, K. et al. The SUMO isopeptidase SENP6 functions as a rheostat of chromatin residency in genome maintenance and chromosome dynamics. Cell Rep. 29, 480–494.e5 (2019).
Lima, C. D. & Reverter, D. Structure of the human SENP7 catalytic ___domain and poly-SUMO deconjugation activities for SENP6 and SENP7. J. Biol. Chem. 283, 32045–32055 (2008).
Hay, R. T. SUMO-specific proteases: a twist in the tail. Trends Cell Biol. 17, 370–376 (2007).
Hang, J. & Dasso, M. Association of the Human SUMO-1 Protease SENP2 with the Nuclear Pore *. J. Biol. Chem. 277, 19961–19966 (2002).
Reverter, D. & Lima, C. D. A basis for SUMO protease specificity provided by analysis of human Senp2 and a Senp2-SUMO complex. Structure 12, 1519–1531 (2004).
Reverter, D. & Lima, C. D. Structural basis for SENP2 protease interactions with SUMO precursors and conjugated substrates. Nat. Struct. Mol. Biol. 13, 1060–1068 (2006).
Shen, L. et al. SUMO protease SENP1 induces isomerization of the scissile peptide bond. Nat. Struct. Mol. Biol. 13, 1069–1077 (2006).
Liu, W. et al. Lactate regulates cell cycle by remodelling the anaphase promoting complex. Nature 616, 790–797 (2023).
Heo, K.-S. et al. Disturbed flow-activated p90RSK kinase accelerates atherosclerosis by inhibiting SENP2 function. J. Clin. Invest. 125, 1299–1310 (2015).
Raman, N. et al. mTOR signaling regulates nucleolar targeting of the SUMO-specific isopeptidase SENP3. Mol. Cell Biol. 34, 4474–4484 (2014).
Li, Y.-Y. et al. TCR-induced tyrosine phosphorylation at Tyr270 of SUMO protease SENP1 by Lck modulates SENP1 enzyme activity and specificity. Front Cell Dev. Biol. 9, 789348 (2022).
Jiang, M. et al. SUMO-specific protease 2 in Mdm2-mediated regulation of p53. Cell Death Differ. 18, 1005–1015 (2011).
Cubeñas-Potts, C. et al. SENP1 and SENP2 affect spatial and temporal control of sumoylation in mitosis. Mol. Biol. Cell 24, 3483–3495 (2013).
Goeres, J. et al. The SUMO-specific isopeptidase SENP2 associates dynamically with nuclear pore complexes through interactions with karyopherins and the Nup107-160 nucleoporin subcomplex. MBoC 22, 4868–4882 (2011).
Gong, L. & Yeh, E. T. H. Characterization of a family of nucleolar SUMO-specific proteases with preference for SUMO-2 or SUMO-3. J. Biol. Chem. 281, 15869–15877 (2006).
Haindl, M. et al. The nucleolar SUMO-specific protease SENP3 reverses SUMO modification of nucleophosmin and is required for rRNA processing. EMBO Rep. 9, 273–279 (2008).
Yun, C. et al. Nucleolar protein B23/nucleophosmin regulates the vertebrate SUMO pathway through SENP3 and SENP5 proteases. J. Cell Biol. 183, 589–595 (2008).
Zunino, R. et al. The SUMO protease SENP5 is required to maintain mitochondrial morphology and function. J. Cell Sci. 120, 1178–1188 (2007).
Zunino, R. et al. Translocation of SenP5 from the nucleoli to the mitochondria modulates DRP1-dependent fission during mitosis. J. Biol. Chem. 284, 17783–17795 (2009).
Maison, C. et al. The SUMO protease SENP7 is a critical component to ensure HP1 enrichment at pericentric heterochromatin. Nat. Struct. Mol. Biol. 19, 458–460 (2012).
Bawa-Khalfe, T. et al. Differential expression of SUMO-specific protease 7 variants regulates epithelial-mesenchymal transition. Proc. Natl. Acad. Sci. USA 109, 17466–17471 (2012).
Romeo, K. et al. The SENP7 SUMO-protease presents a module of Two HP1 interaction motifs that Locks HP1 protein at pericentric heterochromatin. Cell Rep. 10, 771–782 (2015).
Yamada, S. et al. Drp1 SUMO/deSUMOylation by Senp5 isoforms influences ER tubulation and mitochondrial dynamics to regulate brain development. iScience 24, 103484 (2021).
Di Bacco, A. & Gill, G. SUMO-specific proteases and the cell cycle: An essential role for SENP5 in cell proliferation. Cell Cycle 5, 2310–2313 (2006).
Kim, E. Y. et al. SENP5, a SUMO isopeptidase, induces apoptosis and cardiomyopathy. J. Mol. Cell Cardiol. 78, 154–164 (2015).
Panse, V. G. et al. Formation and nuclear export of preribosomes are functionally linked to the small-ubiquitin-related modifier pathway. Traffic 7, 1311–1321 (2006).
Dönig, J. et al. Characterization of nucleolar SUMO isopeptidases unveils a general p53-independent checkpoint of impaired ribosome biogenesis. Nat. Commun. 14, 8121 (2023).
Yu, B. et al. SUMO proteases SENP3 and SENP5 spatiotemporally regulate the kinase activity of Aurora A. J. Cell Sci. 134, jcs249771 (2021).
Li, Y. et al. Structural basis for the SUMO2 isoform specificity of SENP7. J. Mol. Biol. 434, 167875 (2022).
Alegre, K. O. & Reverter, D. Swapping small ubiquitin-like modifier (SUMO) isoform specificity of SUMO proteases SENP6 and SENP7. J. Biol. Chem. 286, 36142–36151 (2011).
Alegre, K. O. & Reverter, D. Structural insights into the SENP6 Loop1 structure in complex with SUMO2. Protein Sci. 23, 433–441 (2014).
Di Bacco, A. et al. The SUMO-specific protease SENP5 is required for cell division. Mol. Cell Biol. 26, 4489–4498 (2006).
Mulder, M. P. C. et al. Total chemical synthesis of SUMO and SUMO-based probes for profiling the activity of SUMO-specific proteases. Angew. Chem. Int. Ed. 57, 8958–8962 (2018).
Sommer, S. et al. Covalent inhibition of SUMO and ubiquitin-specific cysteine proteases by an in situ thiol–alkyne addition. Bioorg. Med. Chem. 21, 2511–2517 (2013).
Hemelaar, J. et al. Chemistry-based functional proteomics: mechanism-based activity-profiling tools for ubiquitin and ubiquitin-like specific proteases. J. Proteome Res. 3, 268–276 (2004).
Krissinel, E. & Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 372, 774–797 (2007).
Reverter, D. et al. Structure of a complex between Nedd8 and the Ulp/Senp protease family member Den1. J. Mol. Biol. 345, 141–151 (2005).
Gan-Erdene, T. et al. Identification and characterization of DEN1, a deneddylase of the ULP family *. J. Biol. Chem. 278, 28892–28900 (2003).
Garvin, A. J. et al. The deSUMOylase SENP7 promotes chromatin relaxation for homologous recombination DNA repair. EMBO Rep. 14, 975–983 (2013).
Claessens, L. A. et al. SENP6 regulates localization and nuclear condensation of DNA damage response proteins by group deSUMOylation. Nat. Commun. 14, 5893 (2023).
Liebelt, F. et al. The poly-SUMO2/3 protease SENP6 enables assembly of the constitutive centromere-associated network by group deSUMOylation. Nat. Commun. 10, 3987 (2019).
Albuquerque et al. Binding to small ubiquitin-like modifier and the nucleolar protein Csm1 regulates substrate specificity of the Ulp2 protease. J. Biol. Chem. 293, 12105–12119 (2018).
Roberts, C. G. et al. Ubiquitin-targeted bacterial effectors: rule breakers of the ubiquitin system. EMBO J. 42, e114318 (2023).
Pruneda, J. N. et al. The molecular basis for ubiquitin and ubiquitin-like specificities in bacterial effector proteases. Mol. Cell 63, 261–276 (2016).
Sánchez-Alba, L. et al. Structural diversity of the CE-clan proteases in bacteria to disarm host ubiquitin defenses. Trends Biochem. Sci. 49, 1111–1123 (2024).
Lara-Ureña, N. et al. Cancer-associated dysregulation of sumo regulators: proteases and ligases. Int. J. Mol. Sci. 23, 8012 (2022).
van den Ent, F. & Löwe, J. RF cloning: a restriction-free method for inserting target genes into plasmids. J. Biochem Biophys. Methods 67, 67–74 (2006).
Bachman, J. Chapter Ninteen - Site-directed mutagenesis. In Methods in Enzymology Vol. 529 (ed. Lorsch, J.), pp. 241–248, (Academic Press, 2013).
Vernier, M. et al. Regulation of E2Fs and senescence by PML nuclear bodies. Genes Dev. 25, 41–50 (2011).
Terui, Y. et al. Dual role of sumoylation in the nuclear localization and transcriptional activation of NFAT1. J. Biol. Chem. 279, 28257–28265 (2004).
Békés, M. et al. The dynamics and mechanism of SUMO chain deconjugation by SUMO-specific proteases. J. Biol. Chem. 286, 10238–10247 (2011).
Kooij, R. et al. Small-molecule activity-based probe for monitoring ubiquitin C-terminal hydrolase L1 (UCHL1) activity in live cells and zebrafish embryos. J. Am. Chem. Soc. 142, 16825–16841 (2020).
Juanhuix, J. et al. Developments in optics and performance at BL13-XALOC, the macromolecular crystallography beamline at the Alba Synchrotron. J. Synchrotron Rad. 21, 679–689 (2014).
Vonrhein, C. et al. Data processing and analysis with the autoPROC toolbox. Acta Cryst. D. 67, 293–302 (2011).
Emsley, P. et al. Features and development of Coot. Acta Cryst. D. 66, 486–501 (2010).
Adams, P. D. et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Cryst. D. 66, 213–221 (2010).
Acknowledgements
This work was supported by the Spanish Ministry of Science and Innovation (MICINN) PID2021−124602OB-I00 to D.R. and by ICREA to D.R., ICREA-Academia-2022, from Generalitat de Catalunya. Y.L. acknowledges her support by the National Natural Science Foundation of China (grant No.32300040). L.S.A. acknowledges her FPI fellowship from the Spanish Government (grant No PRE2019-088509). MPCM acknowledges her support by the Netherlands Organization for Scientific Research (NWO) (VIDI Grant VI. 213.110). D.R. acknowledges support from the Serra Hunter program from Generalitat de Catalunya. X-ray experiments were performed at BL-13 XALOC beamline at ALBA Synchrotron with the collaboration of ALBA staff.
Author information
Authors and Affiliations
Contributions
Y.L. and L.S.A. conducted crystallization experiments. Y.L., L.S.A., B.L., H.B.G., and N.V. conducted all in vitro activity assays. L.S.A., A.D.B., and V.A. conducted cultured cell assays. M.D.M. and M.P.C.M. synthesized SUMO chemical probes and conducted cell lysate labeling. D.R., N.V., and L.S.A. contributed to the correction and writing of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Sánchez-Alba, L., Ying, L., Maletic, M.D. et al. Structural basis for the human SENP5’s SUMO isoform discrimination. Nat Commun 16, 4764 (2025). https://doi.org/10.1038/s41467-025-60029-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-025-60029-4
