
Abstract
Recent studies have highlighted the potential of ferroptosis in treating breast cancer. However, the efficacy of ferroptosis induction in the most common subtype, estrogen receptor-positive (ER + ) breast cancer, remains inadequately explored. This study unveils that both short-term and long-term treatment with ER-targeted endocrine agents sensitizes ER+ breast cancer cells to ferroptosis inducers, particularly the GPX4 inhibitor, revealing a non-mutational sensitization mechanism. Based on this finding, we introduce a 55-gene signature score (FERscore) tailored to assess ferroptosis susceptibility in breast cancer. Data from cell lines and primary tumors demonstrate significant lower FERscores in ER+ breast cancer compared to other subtypes; however, FERscores dramatically increase in endocrine-resistant ER+ tumor cells and residual tumors post-endocrine therapy. Furthermore, FERscore correlates positively with mesenchymal traits, stemness, immune cell infiltration, and cancer-associated fibroblasts enrichment, while inversely correlating with estrogen responsiveness and DNA repair capacity. Additionally, the FERscore proves effective in predicting therapeutic responses to anti-ER, anti-HER2, poly (ADP-ribose) polymerase inhibitor, and anti-angiogenesis therapies in breast cancer. In summary, ferroptosis induction emerges as a promising avenue in breast cancer therapy. The FERscore offers an innovative tool for identifying patients who may benefit from ferroptosis-inducing therapies, especially those responsive to GPX4 inhibitors.
Similar content being viewed by others


Introduction
Ferroptosis is a form of programmed cell death mediated by reactive oxygen species-induced lipid peroxidation and requires intracellular iron1. It has been implicated in various pathological conditions, including degenerative diseases, ischemic disorders, and carcinogenesis2. Therefore, molecules involved in ferroptosis signaling pathways are potential therapeutic targets for these diseases. Recently, ferroptosis has provided new insights into the therapeutic strategies for breast cancer. In the HER2-enriched subtype, lapatinib-tolerant persister cells derived from the breast cancer BT474 cell line exhibited a dependency on the lipid hydroperoxidase, glutathione peroxidase 4 (GPX4)3. Shao and colleagues identified the heterogeneity of ferroptosis-related pathways and metabolites across four transcriptomic subtypes of triple-negative breast cancer (TNBC). Targeting glutathione metabolism, particularly GPX4, can induce ferroptosis in the luminal androgen receptor (LAR) subtype4. Estrogen receptor-positive (ER + ) breast cancer, constitutes nearly 80% of all breast cancers5. However, it remains unclear whether ferroptosis induction holds great promise for the treatment of ER+ breast cancer.
De novo and acquired resistance to endocrine therapy is a major challenge in treating ER+ breast cancers. Recent seminal studies identified vulnerability to ferroptosis as a common feature of therapy-resistant cancer cells in the mesenchymal state across different cancer lineages3,6. Remarkably, the acquisition of a mesenchymal-like morphology was observed during the induction of endocrine therapy-resistant ER+ breast cancer cell lines, consistent with previous report7. This study aims to investigate the sensitivity of ER+ breast cancer cells to ferroptosis inducers under various conditions and to establish a specific ferroptosis-related gene signature (FERscore) for evaluating susceptibility to ferroptosis in breast cancer.
Results
Enhancing sensitivity to ferroptosis in ER+ breast cancer cells via extended and short-term endocrine drug exposure
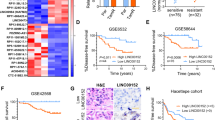
To explore whether therapy-resistant breast cancer cells in the mesenchymal state are sensitive to ferroptosis inducers, we first performed GSEA using public data from GSE67916 dataset. The results revealed significant enrichment in the EMT (NES = 1.63, p = 5*10−4) and ferroptosis-related pathways (NES = 1.60, p = 0.0102) in tamoxifen-resistant MCF7 cells compared to the parental cell line (Fig. 1A). In vitro data showed that both 4-hydroxytamoxifen- and fulvestrant-induced endocrine-resistant T47D cells (T47D-TamR and T47D-FulvR) were considerably more vulnerable to the classic ferroptosis inducer, GPX4 inhibitor RSL3, compared to the parental cell line (T47D-P) (Fig. 1B, C). Specifically, the median survival rates of T47D-TamR and T47D-FulvR were approximately half of that of T47D-P cell line after exposing to RSL3 at a dose of 1 µM for three days.

A The enrichment of hallmark_epithelial-mesenchymal transition and wp_ferroptosis (WP4313 from the Wikipathways database) gene sets between breast cancer tamoxifen-resistant and parental MCF7 cell lines by GSEA (data from GSE67916 dataset). Fractional viabilities (B) and morphological changes (C) of tamoxifen-resistant, fulvestrant-resistant and parental T47D cell lines treated with GPX4 inhibitor RSL3. Data represent mean ± standard deviation of n = 6 biological replicates. D The enrichment of ferroptosis-related gene sets between breast cancer patients before and after short-term neoadjuvant endocrine therapy by GSEA (data from GSE147271 dataset). E Fractional viabilities of T47D cell lines pretreated with 4-hydroxytamoxifen or fulvestrant for 2-, 4- and 6-days before RSL3. Data represent mean ± standard deviation of n = 6 biological replicates. *P < 0.05; **P < 0.01; ****P < 0.0001; ns, not significant (P > 0.05); two-tailed t-tests. F Morphological changes in T47D cells treated with 4-hydroxytamoxifen or fulvestrant for 6 days. All data are representative of two separate experiments.
To further validate our results in clinical practice, we analyzed the transcriptome differences in patients with breast cancer before and after neoadjuvant endocrine therapy. Based on GSEA of RNA-seq data (the GSE147271 dataset) from a pre-operative window trial with a regimen of tamoxifen neoadjuvant therapy, ferroptosis (NES = 1.78, p = 0.0011) and reactive oxygen pathways (NES = 1.71, p = 0.0053) were significantly enriched in tumors after a median duration of tamoxifen therapy for 20.7 ± 9.6 days (Fig. 1D).
Due to the relatively brief duration of endocrine therapy in the aforementioned trial, we inferred that ER+ breast cancer cells became sensitized to ferroptosis after short-term induction of endocrine drugs, which was not unique for long-term induced endocrine-resistant cells. Notably, we found that T47D cells became susceptible to RSL3 after only six days of administration of 4-hydroxytamoxifen or fulvestrant (T47D-Tam/6d and T47D-Fulv/6d, respectively, Fig. 1E, F). Indeed, short-term induced T47D cells showed a more pronounced sensitivity to RSL3 and also a moderately enhanced sensitivity to erastin (Fig. 2A, B), an inhibitor of the solute carrier SLC7A11 which acts upstream of GPX4 by depleting glutathione8. Conversely, both T47D-Tam/6d and T47D-Fulv/6d cells exhibited reduced vulnerability to chemotherapeutic drug 5-fluorouracil (5-FU) compared to parental cells, most likely because of cell cycle arrest caused by endocrine drugs (Fig. 2C). Similar results were observed in MCF7 cells (Supplementary Fig. 1A–C). To further confirm that the above breast cancer cells underwent canonical ferroptosis under GPX4 inhibition, the lipid peroxidation levels of both T47D and MCF7 cells were quantified after different pretreatments using the fluorescent probe C11-BODIPY (581/591). Cells that pretreated with endocrine drugs after a short-term exposure followed by RSL3 induction exhibited a significant increase in cellular lipid peroxidation levels (Fig. 2D, E and Supplementary Fig. 1D, E). Besides, RSL3-induced cell death was largely restrained when co-cultured with the ferroptosis-rescuing lipophilic antioxidant ferrostatin-1, the iron chelator DFO, or the lipoxygenase inhibitors NDGA (Fig. 2F and Supplementary Fig. 1F).

Fractional viabilities of T47D parental and 4-hydroxytamoxifen or fulvestrant pretreated cells treated with (A) RSL3, (B) system Xc- inhibitor Erastin, and (C) chemotherapeutic agent 5-Fluorouracil. Data represent mean ± standard deviation of n = 6 biological replicates. Lipid peroxidation stained by BODIPY-C11 (D) and quantified via flow cytometry (E) for 4-hydroxytamoxifen-pretreated, fulvestrant-pretreated, and parental T47D cell lines before and after exposed to RSL3 for 3 days. F Fractional viabilities of 4-hydroxytamoxifen- or fulvestrant-pretreated T47D cell lines treated with RSL3 and ferroptosis rescue compounds for 3 days. Data represent mean ± standard deviation of n = 6 biological replicates. *P < 0.05; ***P < 0.001; ns, not significant (P > 0.05); two-tailed t-tests. All data are representative of two separate experiments.
Collectively, the ER+ breast cancer cells became adequately sensitive to ferroptosis inducers, especially the GPX4 inhibitor RSL3, immediately after short-term pretreatment with endocrine drugs.
Establishment and validation of a robust signature to predict the sensitivity of breast cancers to ferroptosis
A total of 906 and 1969 differentially expressed genes (DEGs) between T47D-Tam/6d and parental cells, and T47D-Fulv/6d and parental cells (log FC > 1, FDR < 0.05) were obtained, which were called gene sets 1 and 2, respectively. Spearman’s correlation analysis of 40 breast cancer cell-line gene-expression data from the CCLE portal and their corresponding AUC data of the three most classical GPX4 targeting small molecules, including RSL3, ML162, and ML210, were calculated. Erastin was excluded due to the minimal overlap in gene correlation patterns compared to other three GPX4 inhibitors. Genes with a correlation coefficient < ˗0.25 were described as gene set 3, containing a total of 1427 genes. Fifty-five genes shared across these three gene pools were proposed to represent the gene set most relevant to ferroptosis sensitivity in breast cancer (Supplementary Table 1).
The 55 identified genes correlated with each other (Supplementary Fig. 2A). GO and KEGG analyses revealed the functions enriched, including ROS-related pathways (response to ROS and hypoxia-inducible factor-1 signaling pathway), EMT-related pathways (epithelial cell migration and fibroblast proliferation), and immune infiltration-related pathways (regulation of inflammatory response and IL-17 signaling pathway) (Supplementary Fig. 2B, C). As detailed in Supplementary Table 2 and 3, significant correlations were uncovered between our FERscore and the expression levels of established ferroptosis regulator genes. Subsequently, we investigated the predictive ability of our FERscore in determining the sensitivity to ferroptosis in both ER+ and non-ER+ breast cancer subtypes. First, we calculated Spearman’s correlation coefficients between the AUCs of mechanism-diverse compounds that induce ferroptosis and FERscores of the corresponding cell lines using breast cancer cell-line data from the CTRP and CCLE databases. Remarkably, the results showed consistent negative correlations (Table 1), suggesting that higher FERscores corresponded to increased drug sensitivity to ferroptosis. Next, we delved deeper into validating the applicability of our FERscore across breast cancer subtypes. The lapatinib-tolerant BT474 persister cells, confirmed to be GPX4-dependent, exhibit significantly higher sensitivity to RSL3 and ML2103. Their FERscores are notably elevated compared to the parental cells (Fig. 3A). The LAR subtype in TNBC is distinguished by the activation of numerous ferroptosis-related pathways and the accumulation of glutathione-related metabolites4,9. In vitro data suggests that RSL3 has a notably greater impact on LAR tumors4, as confirmed by higher FERscores within this subtype (Fig. 3B). Our in vitro data demonstrated vulnerability to RSL3 in T47D and MCF7 cell lines, after both long- and short-term induction with tamoxifen or fulvestrant. We utilized our own cell line sequencing data, along with data from ER+ breast cancer cell lines and patients before and after endocrine treatment sourced from public datasets, to test the robustness of our FERscore. As depicted in Fig. 3C–G, the results illustrated that the FERscores for both long- and short-term induced breast cancers are significantly higher than those in parental tumor cells from both cell lines and breast cancer patients. Collectively, these results demonstrated the efficacy of our FERscore for assessing ferroptosis sensitivity in breast cancer. However, the existing ferroptosis pathway or ferroptosis-related risk scores10,11 are not applicable (Supplementary Fig. 2D).

A FERscores in parental and lapatinib-induced persister BT474 cells. B FERscores in the luminal androgen receptor (LAR) and non-LAR subtypes of triple-negative breast cancer. FERscores in parental, tamoxifen-resistant and fulvestrant-resistant (C) T47D and (D) MCF7 cells. E FERscores in T47D and MCF7 cells treated with endocrine drugs after a short time. F, G FERscores in ER+ breast cancer patients before and after neoadjuvant endocrine therapy (baseline: pretreatment, mid: after 10–14 days Letrozol, post: after 90 days Letrozol in GSE20181; NAET-Tam: after a median duration of tamoxifen for 20.7 ± 9.6 days in GSE147271). The distribution of FERscores for (H) breast cancer cell lines (data from CCLE) and (I, J) breast cancer patients (data from METABRIC dataset) across different subtypes. For (J), Chischisq-test were used. K Kaplan-Meier (KM) plot of the differential prognosis between breast cancer patients in groups of high or low FERscores. For figures (A–I), box-and-whisker plots indicate median; 25th and 75th percentiles; whiskers, minima and maxima of the distributions; and the Wilcoxon signed rank test was used. For (K), a log-rank test is used to compare the difference in overall survival.
Next, we assessed the distribution of FERscores across different breast cancer subtypes. As illustrated in Fig. 3H, the ER+ cell lines exhibited the lowest scores. Similar results were observed for primary breast cancers in the METABRIC case series (Fig. 3I). The proportion of ER-negative patients was more than twice in the high-FERscore subgroup, whereas 75% of patients in the low-FERscore subgroup were ER+HER2- (Fig. 3J). Furthermore, prognostic analysis revealed a prominent survival disadvantage in patients with high FERscores (Fig. 3K). As anticipated, using the TCGA-BRCA cohort as a validation set, we obtained concordant results (Supplementary Fig. 2E, F).
Collectively, these results strongly emphasize the robustness and effectiveness of our FERscore for predicting sensitivity to ferroptosis in breast cancer.
FERscore reflects certain core characteristics of tumor and tumor microenvironment in breast cancer
To determine the potential signaling pathways and biological processes enriched in the high- or low-FERscore subgroup as categorized above, we performed GSEA using the HALLMARK gene sets. A total of 30 pathways were significantly enriched (FDR-adjusted p < 0.05) in METABRIC (Fig. 4A) and TCGA-BRCA cohorts (Supplementary Fig. 3A). The functional classes of the significant gene sets in the high-FERscore subgroup included tumor growth and invasion, signal transduction, angiogenesis, and immune activation, whereas pathways related to cell cycle, DNA repair, and estrogen response were identified in the low-FERscore subgroup. Utilizing a differentiation-related gene set consisting of 49 genes12, we confirmed that the FERscore was notably associated with the score levels of immature basal/stem and luminal progenitor cells in breast cancer (Fig. 4B). Similarly, the scores of four distinct EMT signatures13,14, and the expression levels of EMT-related transcription factors were significantly higher in the high-FERscore subgroup (Fig. 4C, D). In contrast, the scores of pathways involved in DNA damage response, DNA replication, and cell cycle were significantly higher in the low-FERscore subgroup (Fig. 4E). These results suggest a strong correlation between our FERscore and specific core tumor characteristics of breast cancer, potentially influencing the sensitivity of breast cancer cells to ferroptosis induction.

A Hallmark gene sets enriched in breast cancers with high- and low-FERscores. Correlation between FERscore and (B) stemness, (C) epithelial-mesenchymal transition (EMT), (D) EMT-related transcription factors, and (E) DNA damage and repair. F The Spearman correlation between the tumor microenvironment (TME) and FERscore. G Correlation between the FERscore and the abundance of specific infiltrated immune and stromal cells computed according to the xCell algorithm. H Differences in expression of immune checkpoints and co-stimulatory receptors between high- and low-FERscore groups. For (B, C, E), box plots indicate median; 25th and 75th percentiles; ***P < 0.001; ns, not significant (P > 0.05); two-tailed t-tests. For (D, F, H), box-and-whisker plots indicate median; 25th and 75th percentiles; whiskers, minima and maxima of the distributions; *P < 0.05; ***P < 0.001; ns, not significant (P > 0.05); the Wilcoxon signed rank test.
To establish a correlation between our FERscore and the characteristics of the tumor microenvironment (TME) in breast cancer, we visualized the Spearman’s correlation coefficients between the ESTIMATE score, immune score, stromal score, tumor purity and the FERScore. Except for tumor purity, all these characteristics were linearly positively correlated with the FERscore (Fig. 4F). Subsequently, we analyzed the correlation between the FERscore and the abundance of specific infiltrated immune and stromal cells computed according to the xCell algorithm15. As presented in Fig. 4G, the relative infiltration level of stromal cells, as well as various adaptive and innate immune cells were significantly positively correlated with the FERscore of breast cancer. Moreover, the expressions of the representative immune checkpoints and co-stimulatory receptors, including TNFRSF4 (OX40), TNFRSF 9 (4-1BB, CD137), ICOS, SLAMF1, CTLA4, IDO1, and BTLA, were much higher in the high-FERscore subgroup (Fig. 4H).
Among the stromal cells analyzed, fibroblasts showed the strongest correlation with our FERscore. Fibroblasts, referred to as cancer-associated fibroblasts (CAFs), are important components of the tumor stroma and have been implicated in tumor initiation, progression, and metastasis16. The signature scores of TGFβ-related pathways13, which are critical mechanisms driving the functions of CAFs, were evidently more enriched in the breast cancer subgroup with high FERscores (Fig. 5A). To better illustrate the relationship between the FERscore and CAFs, we applied three different algorithms to infer the accumulation of CAFs in breast cancer cases. Correlation analysis indicated consistent results among algorithms. The levels of CAFs were highly positively correlated with the expression of 55 ferroptosis sensitivity-related genes and FERscores (Fig. 5B). Subsequently, we screened out a total of 75 candidate cytokines that were notably associated with CAFs calculated by all three algorithms (R > 0.3, Fig. 5C). We compared the differences in gene expression between high- and low-FERscore subgroups using three independent datasets, including the METABRIC, TCGA-BRCA, and GSE20685 patient cohorts, and found that the median expression values of these 75 cytokines were markedly higher in the high-FERscore cases (Fig. 5D).

A Differences in signatures of TGFβ-related pathways between high- and low-FERscore groups. Pan-F-TBRS: pan-fibroblast TGFβ response signature. Box plots indicate median; 25th and 75th percentiles; ***P < 0.001; two-tailed t-tests. B Correlation between expression of the 55 ferroptosis-related genes as well as FERscores and the infiltrated CAF levels. C Candidate CAF-associated cytokines. D Differences in the expression of 75 cytokines between high- and low-FERscore groups from 3 clinical cohorts, including METABRIC, TCGA-BRCA and GSE20685 datasets.
Collectively, certain core characteristics of the tumor and tumor microenvironment in breast cancer represent the biological significance of the FERscore. These characteristics accurately reflect the sensitivity of specific breast cancer subtypes or cell lines to ferroptosis induction.
The FERscore predicts the therapeutic efficacy of different drugs for breast cancer
We attempted to determine the role of our FERscore in predicting the therapeutic efficacy of different drugs and identifying situations where ferroptosis induction could be a potentially effective approach for breast cancer treatment. The Spearman’s correlation results between the FERscores and AUCs of multiple therapeutic agents from breast cancer cell lines varied considerably depending on the drugs (Table 2, Supplementary Fig. 3B). A positive correlation between the FERscore and the sensitivity of anti-HER2 agents and taxanes was observed, while anti-ER agents and cyclophosphamide exhibited the opposite pattern. However, the FERscore could not adequately indicate the response to most chemotherapeutic drugs.
We then studied the FERscore levels in different breast cancer subgroups classified by therapeutic response. Based on data from the GSE20181 dataset, the FERscore levels for ER+ breast cancers responding to letrozole were much lower than those of non-responders (Fig. 6A). For HER2+ patients, those with high FERscores showed an increased susceptibility to anti-HER2 therapies (Fig. 6B–E). Using data from the I-SPY2 neoadjuvant platform trials17,18, we confirmed the significant therapeutic advantages and clinical response to the combination of PD-L1 inhibitor durvalumab and PARP inhibitor olaparib and anti-angiogenesis therapy in patients with high FERscores compared to those with low FERscores (Fig. 6F, G). Moreover, our FERscore well predicted response to PARP inhibitor veliparib, but not PD-1 inhibitor pembrolizumab according to data from the GSE194040 dataset (Fig. 6H, I). Chemotherapy remains the cornerstone of breast cancer treatment. However, similar results from three independent data cohorts indicated that the FERscore does not have predictive specificity for chemotherapy in breast cancer (Fig. 6J–L).

A Distribution of FERscores in distinct clinical response groups to neoadjuvant letrozole for ER+ breast cancer patients. B–E Distribution of FERscores in distinct anti-HER2 clinical response groups for HER2+ patients (neoadjuvant therapy regimens: T-DM1 + pertuzumab or paclitaxel + trastuzumab ± pertuzumab). F–I Distribution of FERscores in distinct clinical response groups to PD-L1 blockade, PARP inhibition and collaborative anti-angiogenesis therapy (In GSE173839: neoadjuvant durvalumab with olaparib and paclitaxel therapy; AMG 386: trebananib, an inhibitor of ANG1/2-Tie2 interaction; ABT 888: veliparib, an inhibitor of both PARP-1 and PARP-2). J–L Distribution of FERscores in distinct clinical response groups to chemotherapy in breast cancer (neoadjuvant therapy regimens: doxorubicin + cyclophosphamide followed by ixabepilone or paclitaxel in GSE41998; taxane-based chemotherapy in GSE163882; taxane-anthracycline chemotherapy in GSE25066). Box-and-whisker plots indicate median; 25th and 75th percentiles; whiskers, minima and maxima of the distributions; and the Wilcoxon signed rank test was used.
Collectively, our work strongly indicated that the FERscore is an effective index for predicting the therapeutic efficacy of both classical and innovative drugs for breast cancer.
Discussion
The present study revealed for the first time that ferroptosis is a potential therapeutic target for ER+ breast cancer. Short-term induction with endocrine agents sensitizes ER+ breast cancer cells to ferroptosis inducers, indicating a non-mutational sensitization mechanism. Based on this, we originally constructed a 55-gene signature score - FERscore to exclusively predict susceptibility to ferroptosis in breast cancer. Breast cancer patients with low FERscores displayed significantly prolonged survival. Both cell line and primary cancer data indicated that the FERscore in the ER+ subtype was much lower than that in other subtypes; however, it dramatically increased in endocrine-resistant ER+ tumor cells and residual tumors after endocrine therapy. In breast cancer, the FERscore level was positively correlated with characteristics such as mesenchymal state, stemness, immune cell infiltration, and CAF enrichment; however, it was negatively correlated with features such as estrogen response and DNA repair capacity. We also demonstrated that our FERscore is an effective index for predicting the therapeutic response of anti-ER, anti-HER2, PARP inhibitor, and anti-angiogenesis therapies in breast cancer, but not for chemotherapy or monospecific checkpoint blockade.
Multiple studies have indicated that ferroptosis plays an important role in the occurrence, development and therapeutic resistance of different tumor types. Therapies targeting tumor ferroptosis have emerged as innovative cancer-curing strategies; however, they remain in the preclinical stage19,20,21. According to the literature, drug-resistant cancer cells enriched in EMT traits exhibited a survival dependency on GPX46. Short-term chemotherapy or targeted-therapy led to a residual population of cancer cells, which also acquired vulnerability to GPX4 inhibitors with increased expression of mesenchymal-like and stemness marker genes3. However, studies on ferroptosis in breast cancer have mostly focused on the HER2-positive and triple-negative subtypes3,4,22. Therefore, the potential of ferroptosis induction as a therapeutic strategy for ER+ breast cancer remains unclear. Our study found that primary ER+ breast cancers were mostly categorized into the low-FERscore subgroup and exhibited irresponsiveness to ferroptosis inducers, and their vulnerability to ferroptosis was closely correlated with resistance to endocrine therapy. Besides, we found that short-term exposure to ER-targeted drugs remarkably increased the ferroptosis sensitivity in ER+ breast cancer cells. This result indicated a non-mutational sensitization mechanism, which remains uncertain. Recently, Jiang and colleagues revealed that ER could transcriptionally upregulate the expression of membrane bound O-acyltransferase ___domain containing 1 (MBOAT1) in ER+ breast cancer, which suppresses ferroptosis through phospholipid remodeling independently of GPX423. This signaling axis can serve as one of the key molecular mechanisms explaining the intriguing phenomena we have discovered. Similar to the established practice of combining chemotherapy with immunotherapy in TNBC, the early combined or sequential use of endocrine therapy and ferroptosis-inducing drugs in ER+ breast cancer holds promise as a therapeutic approach. However, extensive preclinical studies and clinical trials are imperative to validate its efficacy.
In recent years, various gene- or lncRNA-based ferroptosis-related signatures have been studied and explored. Most of these signatures were derived from the FerrDb database24 and were developed through disease-related prognostic models. These signatures are primarily used to investigate survival outcomes, immune infiltration in the tumor microenvironment, and immunotherapy response in patients with different tumor types10,25,26; however, they are insufficiently specific or sensitive to evaluate ferroptosis susceptibility. In the present study, we developed a breast cancer cell line-derived and patient-confirmed signature of a high FERscore proposed to yield a ferroptosis-sensitive state. Therefore, our FERscore could be used to identify patients who may benefit from ferroptosis-inducing therapy.
Clinically, treatment resistance remains a bottleneck for improving the prognosis of breast cancer patients. Multiple novel prognostic and predictive biomarkers, including individual genes, proteins, and gene signatures, have been incorporated into routine clinical practice or are undergoing phase II or III trials27,28,29,30. Using the MINDACT study as an example, the employment of MammaPrint resulted in chemotherapy avoidance for 46.2% of clinically high-risk ER+ breast cancer patients29. The FERscore was confirmed as a prognostic and predictive biomarker for breast cancer in this study. Our results demonstrated that a high FERscore is closely related to primary and acquired resistance to endocrine therapy in ER+ breast cancer. In the HER2-enriched subtype, breast cancers with concomitantly higher FERscores were found to be the preferred subgroup for anti-HER2 therapeutic strategies. However, as reported by Viswanathan et al.6, there may exist a subclone of anti-HER2 persisting cells with exceptionally high FERscores, making them susceptible to ferroptosis inducers. This suggests a potential need to combine anti-HER2 therapy with ferroptosis induction in HER2-enriched subtype. The development of agents and clinical trials related to ferroptosis induction in tumors is currently underway or being planned31. Further comprehensive research is warranted in our future endeavors to explore the potential of the FERscore as a screening tool to identify patients most suitable for ferroptosis-inducing treatment, particularly GPX4 inhibitors, akin to genetic testing tools such as MammaPrint29 and Oncotype DX30, thereby contributing to the clinical treatment landscape for breast cancer patients.
This study has certain limitations. First, our analyses relied primarily on experimental data and public datasets. Therefore, further validation using clinical samples from larger prospective cohorts is necessary. Second, ferroptosis-inducing agents are not extensively utilized in current clinical practice, resulting in insufficient clinical data to validate and optimize our model. Finally, in this study, we did not thoroughly investigate the mechanism underlying the heightened sensitivity of breast cancer cells to ferroptosis after extended and short-term exposure to endocrine agents. Further studies are warranted to comprehensively investigate this aspect.
In summary, this study offers a novel perspective on the potential utilization of ferroptosis induction as a therapeutic strategy for ER+ breast cancer. Additionally, we identified a specific ferroptosis-related gene signature that demonstrated efficacy in assessing ferroptosis sensitivity and predicting therapeutic response to various drugs in breast cancer. Our FERscore model still requires validation and optimization through prospective clinical data for practical implementation.
Methods
Cell culture
Regular MCF7 and T47D cell lines were obtained from ATCC and cultured according to ATCC’s recommendations in Dulbecco’s modified Eagle’s medium (DMEM; Gibco) with 10% FBS (Gibco) and 1% penicillin/streptomycin (P/S). Endocrine-resistant cells, including 4-hydroxytamoxifen- and fulvestrant-resistant (TamR and FulvR) cell models, were induced following the previous research methods32,33. The endocrine-resistant cells were kept in a phenol-red free medium supplemented with 10% FBS and 1% P/S with the addition of 5 µM 4-hydroxytamoxifen (Tam, HY-16950, MCE) or 0.5 µM fulvestrant (Fulv, S1191, Selleck). All the cells were maintained at 37 °C in a 5% CO2 incubator.
In vitro cell viability assays
For the cell viability assays, cells were seeded at optimal densities in 96-well plates with containing growth medium and allowed to adhere overnight. The optimal seeding densities for MCF7 and T47D cells determined in this study were 2000 and 8000 cell per well, respectively. The following day, the growth medium from the overnight incubation was replaced with 100 μL fresh medium containing the corresponding concentrations of various inhibitors. After 72 h, cell viability was evaluated using a Cell Counting Kit-8 (#40203ES92: Yeasen), and the absorbance was measured at 450 nm using a Cytation 5 imaging reader. Chemicals, including (1 S,3 R)-RSL3 (RSL3, S8155), erastin (S7242), 5-fluorouracil (5-FU, S1209), ferrostatin-1 (S7243), deferoxamine mesylate (DFO, S5742), nordihydroguaiaretic acid (NDGA, S3984), and Z-VAD-FMK (ZVAD, S7023), were purchased from Selleck.
Lipid-peroxidation assay
Cellular lipid peroxidation was measured, as previously described in refs. 6,34. The cells were seeded in a confocal culture dish (2 × 105 cells per dish) with 0.1 μM RSL3 and incubated for 24 h. The cells were then stained with 1 mL of BODIPY- C11 (1.5 μM) at 37 °C for 30 min and observed under a confocal fluorescence microscope or be resuspended in 500 μL of phosphate-buffered saline with 1% 4-(2-hydroxyethyl) piperazine-1-ethanesulfonic acid and analyzed using a flow cytometry system (Beckman).
RNA isolation and RNA sequencing (RNA-seq)
Total RNA was isolated using a GeneJET™ RNA Purification Kit (Thermol) according to the manufacturer’s protocol and 1 μg of RNA was converted to cDNA using the Hifair III 1st Strand cDNA Synthesis SuperMix (Yeasen) in the presence of both oligo (dT) and random primers. RNA integrity was assessed using the RNA Nano 6000 Assay Kit on the Bioanalyzer 2100 system (Agilent Technologies, CA, USA). For sequencing, samples with specific adaptors were sequenced on an Illumina NovaSeq platform, and 150 bp paired-end reads were generated according to the manufacturer’s instructions.
Data source and data preprocessing
The gene-expression profile and clinical data of various breast cancer cell lines and patient cohorts from 16 eligible datasets, including tamoxifen-resistant and parental tamoxifen-sensitive MCF7 (GSE67916) and T47D (GSE125738) cell lines, fulvestrant-resistant and parental fulvestrant-sensitive MCF7 (GSE74391 and GSE85536) and T47D (GSE85536) cell lines, parental and lapatinib-induced persister BT474 cell lines (GSE84896), the Cancer Genome Atlas Breast Invasive Carcinoma (TCGA-BRCA) and Molecular Taxonomy of Breast Cancer International Consortium (METABRIC) cohorts, a Chinese breast cancer cohort (GSE20685), primary breast cancers before and after neoadjuvant tamoxifen therapy (GSE147271), breast cancers from postmenopausal patients before and after neoadjuvant letrozole therapy (GSE20181), breast cancers in different subtypes before receiving different neoadjuvant therapy regimens targeting diverse biology from the I-SPY2 platform trial (GSE181574, GSE194040, and GSE173839), and breast cancers before neoadjuvant chemotherapy (GSE41998, GSE163882, and GSE25066), were collected from the publicly available Gene Expression Omnibus (GEO), TCGA, and METABRIC databases. The FPKM values of TCGA-BRCA dataset were converted into transcripts per kilobase million (TPM). We directly downloaded the normalized microarray data from other platforms. Transcriptome sequencing data related to different breast cancer cell lines were extracted from the Cancer Cell Line Encyclopedia (CCLE) database (https://sites.broadinstitute.org/ccle/). The area-under-the-curve (AUC) metrics as sensitivity measurements of breast cancer cell lines conditioned by various small-molecules were extracted from the Cancer Therapeutics Response Portal (CTRP v2, https://portals.broadinstitute.org/ctrp.v2.1/). A comprehensive dataset of 649 ferroptosis regulator genes, derived from human data, was compiled from the FerrDb V2 database24 and the latest research studies23,35,36.
Gene sets and signature score (FERscore) related to the sensitivity of breast cancers to ferroptosis
The DEGs between T47D cells after six days of induction with 4-hydroxytamoxifen or fulvestrant and parental cells were identified using the DESeq2 R package. The AUC metrics of 40 breast cancer cell lines treated with small molecules, including RSL3, ML162, and ML210, were obtained from the CTRP database. Spearman’s correlation analysis of these cell-line gene-expression data from the CCLE portal and their corresponding AUC data of the three drugs were calculated. We then established a signature score (FERscore) to quantify the sensitivity of breast cancers to ferroptosis using the single-sample gene-set enrichment analysis (ssGSEA) algorithm based on the 55 proposed genes.
Gene set enrichment analysis (GSEA)
The gene sets of “h.all.v7.5.1.symbols.gmt”, “c2.cp.kegg.v7.5.1.symbols”, “c5.go.v7.5.1.symbols”, and “c2.cp.wikipathways.v7.5.1.symbols.gmt” were downloaded from the Molecular Signatures Database (MSigDB) website. The “clusterProfiler” package was used for enrichment analyses of functional annotation estimates to each sample. Statistical significance was set at P < 0.05.
Analysis of the immune features in the tumor microenvironment (TME) of breast cancer correlated with the FERscore
To characterize the immune infiltration in the TME of breast cancer, ssGSEA, xCell15, and CIBERSORT37 algorithms, and the “ESTIMATE” package38 were used. The relative abundance of fibroblasts in cancer samples (CAF.BRCA) was estimated based on several classical cancer-associated fibroblast (CAF) markers, including PDGFRA, PDGFRB, ACTA2, THY1, PDPN, COL1A1, and FAP39. The CAF-related cytokines were derived from a study by Sun et al.40. The correlation between our FERscore and stromal cells in breast cancer was calculated using “Spearman’s” correlation.
Analysis of the tumor biological processes of breast cancer correlated with the FERscore
The gene sets considered as indicators of crucial biological processes were primarily derived from previous reports by Mariathasan et al.13 and Lawson et al.12. These included signatures related to epithelial-mesenchymal transition (EMT), immune response, pan-fibroblast TGFβ response, DNA repair, cell cycle, and breast cancer stemness. Here, we calculated the enrichment scores of the above signatures using ssGSEA and compared the enrichment of these pathways between breast cancers belonging to high- and low-FERscore subgroups.
Statistical Analyses
All statistical analyses were performed using the R (version 4.1.0) and R Bioconductor packages. We applied the “limma” R package to identify the DEGs41. The enrichment analysis of gene ontology (GO), functional annotation, and Kyoto encyclopedia of genes and genomes (KEGG) were performed using the “clusterProfiler” package42. The R packages “survival” and “survminer” were used for survival analysis. The optimal cut-off value between the two groups with high and low FERscore was determined using the surv_cutpoint function43. Unless otherwise noted, unpaired, two-tailed t-test was used to evaluate the differences between the two independent groups. All results shown as stacked graphs, heatmaps, and box and whisker plot are using the ggplot2 R package. Box-and-whisker plots indicate the median, 25th and 75th percentiles, with whiskers representing the minima and maxima of the distributions. These box plots are generated using GraphPad Prism (version 8), with the data representing the mean ± standard deviation of n = 6 biological replicates. Statistical analysis is performed using the Wilcoxon signed-rank test, unpaired two-tailed t-test, and log-rank test. If not specified above, statistical significance was set at P < 0.05.
Data availability
The raw data of RNA sequencing have been deposited in the National Center for Biotechnology Information (NCBI) (https://www.ncbi.nlm.nih.gov/) with accession number PRJNA1129994. The remaining data generated in this study are provided in the Supplementary Information.
Code availability
The underlying code for this study is not publicly available but may be made available to qualified researchers on reasonable request from the corresponding author.
References
Wu, Y. et al. Ferroptosis in Cancer Treatment: Another Way to Rome. Front. Oncol. 10, 571127 (2020).
Stockwell, B. R. et al. Ferroptosis: A Regulated Cell Death Nexus Linking Metabolism, Redox Biology, and Disease. Cell 171, 273–285 (2017).
Hangauer, M. J. et al. Drug-tolerant persister cancer cells are vulnerable to GPX4 inhibition. Nature 551, 247–250 (2017).
Yang, F. et al. Ferroptosis heterogeneity in triple-negative breast cancer reveals an innovative immunotherapy combination strategy. Cell Metab. 35, 84–100.e108 (2023).
Giaquinto, A. N. et al. Breast Cancer Statistics, 2022. CA: A Cancer J. Clin. 72, 524–541 (2022).
Viswanathan, V. S. et al. Dependency of a therapy-resistant state of cancer cells on a lipid peroxidase pathway. Nature 547, 453–457 (2017).
Bi, M. et al. Enhancer reprogramming driven by high-order assemblies of transcription factors promotes phenotypic plasticity and breast cancer endocrine resistance. Nat. cell Biol. 22, 701–715 (2020).
Yagoda, N. et al. RAS-RAF-MEK-dependent oxidative cell death involving voltage-dependent anion channels. Nature 447, 864–868 (2007).
Jiang, Y. Z. et al. Genomic and Transcriptomic Landscape of Triple-Negative Breast Cancers: Subtypes and Treatment Strategies. Cancer cell 35, 428–440.e425 (2019).
Wu, Z. H., Tang, Y., Yu, H. & Li, H. D. The role of ferroptosis in breast cancer patients: a comprehensive analysis. Cell Death Discov. 7, 93 (2021).
Li, P. et al. Development and Validation of a Robust Ferroptosis-Related Gene Panel for Breast Cancer Disease-Specific Survival. Front. cell Developmental Biol. 9, 709180 (2021).
Lawson, D. A. et al. Single-cell analysis reveals a stem-cell program in human metastatic breast cancer cells. Nature 526, 131–135 (2015).
Mariathasan, S. et al. TGFβ attenuates tumour response to PD-L1 blockade by contributing to exclusion of T cells. Nature 554, 544–548 (2018).
Barbie, D. A. et al. Systematic RNA interference reveals that oncogenic KRAS-driven cancers require TBK1. Nature 462, 108–112 (2009).
Aran, D., Hu, Z. & Butte, A. J. xCell: digitally portraying the tissue cellular heterogeneity landscape. Genome Biol. 18, 220 (2017).
Chen, Y., McAndrews, K. M. & Kalluri, R. Clinical and therapeutic relevance of cancer-associated fibroblasts. Nat. Rev. Clin. Oncol. 18, 792–804 (2021).
Pusztai, L. et al. Durvalumab with olaparib and paclitaxel for high-risk HER2-negative stage II/III breast cancer: Results from the adaptively randomized I-SPY2 trial. Cancer cell 39, 989–998.e985 (2021).
Wolf, D. M. et al. Redefining breast cancer subtypes to guide treatment prioritization and maximize response: Predictive biomarkers across 10 cancer therapies. Cancer cell 40, 609–623.e606 (2022).
Chen, X., Kang, R., Kroemer, G. & Tang, D. Broadening horizons: the role of ferroptosis in cancer. Nat. Rev. Clin. Oncol. 18, 280–296 (2021).
Lei, G., Zhuang, L. & Gan, B. Targeting ferroptosis as a vulnerability in cancer. Nat. Rev. Cancer. https://doi.org/10.1038/s41568-022-00459-0 (2022).
Liang, C., Zhang, X., Yang, M. & Dong, X. Recent Progress in Ferroptosis Inducers for Cancer Therapy. Adv. Mater. (Deerfield Beach, Fla.) 31, e1904197 (2019).
Lei, G. et al. BRCA1-mediated dual regulation of ferroptosis exposes a vulnerability to GPX4 and PARP co-inhibition in BRCA1-deficient cancers. Cancer Discov. https://doi.org/10.1158/2159-8290.cd-23-1220 (2024).
Liang, D. et al. Ferroptosis surveillance independent of GPX4 and differentially regulated by sex hormones. Cell 186, 2748–2764.e2722 (2023).
Zhou, N. et al. FerrDb V2: update of the manually curated database of ferroptosis regulators and ferroptosis-disease associations. Nucleic acids Res. 51, D571–d582 (2023).
Shi, Z. et al. Identification and Validation of a Novel Ferroptotic Prognostic Genes-Based Signature of Clear Cell Renal Cell Carcinoma. Cancers 14, https://doi.org/10.3390/cancers14194690 (2022).
Wang, X. et al. A novel oxidative stress- and ferroptosis-related gene prognostic signature for distinguishing cold and hot tumors in colorectal cancer. Front. Immunol. 13, 1043738 (2022).
Tarighati, E., Keivan, H. & Mahani, H. A review of prognostic and predictive biomarkers in breast cancer. Clin. Exp. Med. 23, 1–16 (2023).
Tutt, A. N. J. et al. Adjuvant Olaparib for Patients with BRCA1- or BRCA2-Mutated Breast Cancer. N. Engl. J. Med. 384, 2394–2405 (2021).
Cardoso, F. et al. 70-Gene Signature as an Aid to Treatment Decisions in Early-Stage Breast Cancer. N. Engl. J. Med. 375, 717–729 (2016).
Sparano, J. A. et al. Clinical and Genomic Risk to Guide the Use of Adjuvant Therapy for Breast Cancer. N. Engl. J. Med. 380, 2395–2405 (2019).
Gu, Y. et al. Targeting ferroptosis: Paving new roads for drug design and discovery. Eur. J. Med. Chem. 247, 115015 (2023).
Nagelkerke, A. et al. LAMP3 is involved in tamoxifen resistance in breast cancer cells through the modulation of autophagy. Endocr.-Relat. cancer 21, 101–112 (2014).
Li, Y. et al. Novel Pyrrolopyridone Bromodomain and Extra-Terminal Motif (BET) Inhibitors Effective in Endocrine-Resistant ER+ Breast Cancer with Acquired Resistance to Fulvestrant and Palbociclib. J. Med. Chem. 63, 7186–7210 (2020).
Pan, X. et al. Zinc oxide nanosphere for hydrogen sulfide scavenging and ferroptosis of colorectal cancer. J. Nanobiotechnol. 19, 392 (2021).
Freitas, F. P. et al. 7-Dehydrocholesterol is an endogenous suppressor of ferroptosis. Nature 626, 401–410 (2024).
Li, Y. et al. 7-Dehydrocholesterol dictates ferroptosis sensitivity. Nature 626, 411–418 (2024).
Newman, A. M. et al. Robust enumeration of cell subsets from tissue expression profiles. Nat. methods 12, 453–457 (2015).
Yoshihara, K. et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat. Commun. 4, 2612 (2013).
Zheng, S. et al. Landscape of cancer-associated fibroblasts identifies the secreted biglycan as a protumor and immunosuppressive factor in triple-negative breast cancer. Oncoimmunology 11, 2020984 (2022).
Sun, C. et al. Risk Signature of Cancer-Associated Fibroblast-Secreted Cytokines Associates With Clinical Outcomes of Breast Cancer. Front. Oncol. 11, 628677 (2021).
Ritchie, M. E. et al. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic acids Res. 43, e47 (2015).
Wu, T. et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innov. (Camb. (Mass.)) 2, 100141 (2021).
Kassambara, A., Kosinski, M. & Biecek, P. survminer: Drawing Survival Curves using ‘ggplot2’. (2016).
Acknowledgements
This research was supported in part by National Natural Science Foundation of China (82203077), National Natural Science Foundation of China (81802628) and Bureau and province jointly build traditional Chinese medicine modernization research program of Zhejiang traditional Chinese medicine administration bureau 2023 + ZYY-09-01. The funder played no role in study design, data collection, analysis and interpretation of data, or the writing of this manuscript.
Author information
Authors and Affiliations
Contributions
Y.N.W. and K.M.H. contributed to the concept design of the study and supervised all works. K.M.H. and J.L.Q. drafted the work and revised it critically for important intellectual content; Y.Y.W. and C.C.Y. provided some experimental materials and guidance; Y.H., J.L.Q., and Y.N.W. edited the manuscript with the input from all other authors to ensure that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Hu, K., Qiu, J., Hu, Y. et al. Efficacy of FERscore in predicting sensitivity to ferroptosis inducers in breast cancer. npj Breast Cancer 10, 74 (2024). https://doi.org/10.1038/s41523-024-00685-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41523-024-00685-9
