
Abstract
Tumour-derived microparticles (TMPs), extracellular vesicles traditionally obtained upon ultraviolet (UV) radiation of tumour cells, hold promise in tumour immunotherapies and vaccines and have demonstrated potential as drug delivery systems for tumour treatment. However, concerns remain regarding the limited efficacy and safety of UV-derived TMPs. Here we introduce a microwave (MW)-assisted method for preparing TMPs, termed MW-TMPs. Brief exposure of tumour cells to short-wavelength MW radiation promotes the release of TMPs showing superior in vivo antitumour activity and safety compared with UV-TMPs. MW-TMPs induce immunogenic cell death and reprogramme suppressive tumour immune microenvironments in different lung tumour models, enabling dual targeting of tumour cells by natural killer and T cells. We show that they can efficiently deliver methotrexate to tumours, synergistically boosting the efficacy of PD-L1 blockade. This MW-TMP development strategy is simpler, more efficient and safer than traditional UV-TMP methods.
Similar content being viewed by others


Main
Extracellular vesicles (EVs) have emerged as a promising cell-free platform for tumour therapy, effectively delivering therapeutic agents in oncology1. On the basis of size and biogenesis, EVs are categorized into three types: small EVs (50–150 nm) derived from endosomes, medium-sized EVs (200–800 nm) from plasma membrane-derived ectosomes, including microparticles (MPs) and secreted midbody remnants, and large EVs (~1,000 nm) comprising apoptotic bodies, exospheres and migrasomes2. Tumour-derived microparticles (TMPs), a subtype of EVs originating from tumour cells, have shown potential in tumour immunotherapies and vaccine development owing to their natural targeting capabilities and immunostimulatory properties3. TMPs contain multiple tumour-associated antigens and DNA fragments to activate dendritic cells (DCs), including via the cGAS–STING pathway, eliciting innate immune responses4,5. Our previous study has demonstrated that TMPs engineered with lipopolysaccharide notably promote DC maturation and migration, highlighting their promise as antitumour vaccine candidates6.
Despite advancements, TMP application in cancer treatment remains constrained, primarily attributed to limited production efficiency7. Efforts to enhance TMP production have focused on optimizing upstream processes, including adjusting the medium composition and exposing parent cells to specific stress stimuli8,9. Our previous research has demonstrated that ultraviolet (UV) radiation importantly increases the production of TMPs, which referred to UV-induced TMPs (UV-TMPs)10. Although UV-TMPs loaded with drugs such as methotrexate (MTX) and fluvastatin have effectively inhibited tumour growth in preclinical and clinical studies, stand-alone UV-TMPs lack robust tumoricidal activity11,12. Despite demonstrating acceptable safety profiles and no pro-tumour potential in our earlier study, concerns regarding long-term risks associated with nucleic acid material within TMPs persist13. Therefore, exploring an innovative approach for TMP preparation that improves both biosafety and manufacturing efficiency, while simultaneously enhancing antitumour therapeutic efficacy, is imperative for clinical translation.
Microwave (MW) technology, specifically electromagnetic waves between 900 MHz and 2,450 MHz, is widely used in tumour ablation owing to its advantages of rapid healing, safety and precise targeting14. Here we introduce an MW-assisted method for the preparation of MW-TMPs, utilizing a domestic MW oven to briefly stimulate tumour cells. We first examined the effects of MW irradiation on parental tumour cells and the subsequent release of TMPs. The resultant MW-TMPs were shown to induce immunogenic cell death (ICD) through the shuttling of HMGB1, performing superior antitumour activity in vivo compared with UV-TMPs. Single-cell RNA (scRNA) sequencing and flow cytometry (FCM) analysis revealed that MW-TMPs dual-targeted tumours via natural killer (NK) and T cells, reprogramming the immunosuppressive microenvironment. Validation in clinical malignant pleural effusion (MPE) samples, as well as in zebrafish patient-derived tumour xenograft (PDX) models, confirmed antitumour efficacy, while combinatorial anti-PD-L1 studies in Lewis lung carcinoma (LLC) mice demonstrated synergistic effects. Coupled with their capacity for tumour-targeted MTX delivery, MW-TMPs demonstrated their dual roles as both therapeutic agents and drug delivery nanoplatforms (Fig. 1a–d).

a, MW-TMPs, derived from MW-stimulated dying LLC cells, are enriched with HMGB1 and can effectively deliver small-molecule drugs, such as MTX. b, MW-TMPs induce ICD and prime the subsequent chain reaction of antitumour immunity in the TME. c, MW-TMPs enable dual targeting of tumour cells by NK and T cells and reprogramme TAMs into antitumour subtypes. d, Multiple preclinical tumour models. TCR, T cell receptor.
MW-induced cell death and EV secretion
Morphological analysis revealed that cells exposed to MW exhibited shrinkage, membrane blebbing and pore formation, indicating MW-triggered cytotoxicity (Fig. 2a and Supplementary Fig. 1a). Annexin V staining confirmed elevated apoptosis in MW-treated cells (Supplementary Fig. 1b). The observed membrane disruptions suggest potential involvement of pyroptosis or necroptosis as alternative cell death mechanisms15. To elucidate MW-specific cell death regulation, we conducted transcriptome sequencing. Gene Ontology analysis of differentially expressed genes (DEGs) revealed significant enrichment of inflammatory and apoptotic processes, along with changes in extracellular region and protein binding (Extended Data Fig. 1a). Cell death-related DEGs showed upregulation of Zbp1 (necroptosis inducer), Casp4 (pyroptosis activator) and Gsdmd (pyroptosis executor), validated by qPCR/western blotting (WB), indicating an MW-triggered multifaceted cell death involving not only apoptosis but also immunogenic pyroptosis and inflammatory necroptosis16 (Fig. 2b and Extended Data Fig. 1b,c).

a, SEM images of LLC cells without stimulation (control) or exposed to UV or MW stimulation (n = 3). b, Western blots show the protein levels of EEA1, CLCN3, SH3GL3, ZBP1, CASP4 and GSDMD in the LLCs stimulated with UV or MW (n = 3). c, Characteristics of cell surface detected by atomic force microscope (n = 3). d, Schematic representation of MW-TMP preparation and isolation, created in BioRender. Y, Z. (2025) https://BioRender.com/s46r100. e, Representative transmission electron microscopy images of MW-TMPs and UV-TMPs (n = 3). Scale bars, 100 nm. f, Sodium dodecyl sulfate–polyacrylamide gel electrophoresis gel staining with Coomassie brilliant blue of LLC, UV-TMPs and MW-TMPs (n = 4). g,h, Concentration (g) and size distribution (h) of MW-TMPs induced by multiple MW conditions detected by NTA (n = 3). L10, L20 and L30 represent MW-stimulating cells under 175 W 10 s, 20 s and 30 s, respectively. H10, H20 and H30 refer to MW conditions of 700 W 10 s, 20 s and 30 s, respectively. i, The size distribution of MW-TMPs and UV-TMPs (n = 3). j, Fluorescence intensity of Annexin V within MW-TMPs by FCM (n = 3). The data in g and h are shown in analytic plots as mean ± s.d., and ordinary one-way ANOVA was used. The exact P value was noted.
Next, single-cell Raman spectroscopy revealed consistent spectral signatures across MW- and UV-stimulated cells, with characteristic spectral peaks demonstrating extensive homology to untreated LLC cells (Supplementary Fig. 1c). Notably, MW-stimulated cells exhibited decreased Raman intensity at the main bands (678 cm−1, 784 cm−1, 811 cm−1, 830 cm−1, 916 cm−1, 1,320 cm−1 and 1,610 cm−1), suggesting reduced cellular nucleic acids17,18,19 (Supplementary Fig. 1c). RNA and DNA electrophoresis, along with concentration analyses, confirmed this reduction (Supplementary Fig. 1d,e). Collectively, MW exposure exerts cytotoxic effects on cells, initiating various cell death mechanisms, including apoptosis, necroptosis and pyroptosis, and induces cellular nucleic acid reduction.
EVs are increasingly released under cellular stress, such as starvation, hypoxia and UV irradiation7,20. Atomic force microscopy analysis revealed a higher density of convex protrusions, suggesting enhanced membrane vesicle formation (Fig. 2c). These changes aligned with upregulated genes of endocytosis regulators Sh3gl3 and Eea1 (Extended Data Fig. 1d). Synchronization experiments confirmed elevated messenger RNA and protein levels of these genes (Fig. 2b and Extended Data Fig. 1c).
On the basis of our previous work with UV-TMPs, we successfully isolated MW-TMPs from MW-stimulated LLC suspension by consecutive centrifugation (Fig. 2d). Transmission electron microscopy revealed that both MW-TMPs and UV-TMPs exhibited an elliptical shape (Fig. 2e). Coomassie brilliant blue-stained sodium dodecyl sulfate–polyacrylamide gel electrophoresis showed distinct protein compositions of LLC cells, UV-TMPs and MW-TMPs, suggesting functional changes (Fig. 2f). Nanoparticle tracking analysis (NTA) indicated that the most efficient MW-TMP production occurred at 700 W of MW power for 20 s, whereas lower power or prolonged exposure diminished yields (Fig. 2g). This suggests that optimal MW intensity is crucial for maximal MW-TMP yield, while excessive intensity is counterproductive. The nano-size of MW-TMPs remained consistent under multiple MW conditions (Fig. 2h). Similarly, MW-TMPs and UV-TMPs shared comparable size distribution profiles (Fig. 2i). Notably, MW-TMPs demonstrated higher yield efficiency compared with UV-TMPs, indicating that MW could be a more efficient method for TMP preparation (Supplementary Fig. 2a).
Furthermore, the typical biomarkers of MW-TMPs were determined. Annexin V positivity, a microvesicle marker, was confirmed via FCM21 (Fig. 2j). The presence of critical EV markers such as CD63, CD9 and TSG101 confirmed the EV properties of MW-TMPs (Supplementary Fig. 2b,c). Given the origin of parental cells from lung adenocarcinoma (LUAD), WB of EGFR, EpCAM and HIF-1α confirmed tumorigenic features (Supplementary Fig. 2c). Nucleic acid staining revealed the low levels of nucleic acids in MW-TMPs, benefiting TMP-based therapies by reducing the risk of unintended gene transfer or incorporation (Supplementary Fig. 2d). Stability assessments revealed that MW-TMPs retained size integrity over multiple conditions, indicating their potential for in vivo transport in a monodisperse single-particle state (Supplementary Fig. 2e). These findings confirmed that MW exposure triggers EV production with reduced nucleic acid content and enhanced yield efficiency.
MW-TMPs are enriched with HMGB1 and facilitate ICD
MW-TMPs exhibited time-dependent cellular uptake (Supplementary Fig. 3a,b) and induced apoptosis as effectively as UV-TMPs (Fig. 3a and Extended Data Fig. 2a). Immunologically, MW-TMPs significantly enhanced DC maturation and CD86+CD206− bone marrow-derived macrophages (BMDMs) (M1) expansion, with no significant change in the percentages of CD86−CD206+ BMDMs (M2) compared with untreated or UV-TMP-stimulated cells (Fig. 3b,c and Supplementary Fig. 4a,b). We speculated that the observed immunological effects may stem from various mechanisms, including tumour antigens, the death receptors or immunomodulatory molecules within MW-TMPs, or potential synergistic interactions. To elucidate the underlying mechanisms, we compared MW-TMPs from ovalbumin (OVA)-overexpressing LLC cells (LLC-OVA) versus wild-type cells. Our results indicated that higher OVA levels within MW-TMPs were associated with enhanced immune activation (Supplementary Fig. 4c). Further analysis using MW-TMPs from another cancer cell line with a distinct mutational antigen profile demonstrated comparable T cell distribution and interferon-γ (IFNγ) levels between B16F10- and LLC-derived groups (Supplementary Fig. 4d–f). This suggests that although tumour antigens in MW-TMPs can initiate immune responses, the inherently low immunogenicity of natural LLC-derived MW-TMPs limits their capacity to amplify immune activation through tumour-associated antigens alone.

a, Apoptotic analysis of LLC cells treated with MW-TMPs or UV-TMPs by FCM analysis (n = 3). b, Quantitative data of CD86 expression in DC cells under the indicated treatment (n = 4). c, The percentages of CD86 (standing for M1 type macrophages) in BMDMs treated with MW-TMPs (n = 4). d, The circular diagram illustrating the enrichment analysis of the pathways of interest (n = 3). Statistical significance was defined as log2FC > 2, adjusted P value < 0.05. e, Heat map of cluster analysis for proteomic profiling in MW-TMPs and UV-TMPs samples (n = 3). f, FCM analysis of CRT expression in LLC cells under the indicated treatment (n = 4). g, WB analysis of HMGB1 expression in LLC cells under the indicated treatment (n = 3). h, ATP concentrations in the supernatant of LLC cells under the indicated treatment for 24 h (n = 3). i–k, After the treatment of MW-TMPs (H−M+), HMGB1 inhibitor glycyrrhizic acid (H+M−) or both of them (H+M+), the apoptosis of LLC cells (i), HMGB1 levels (j) and ATP concentrations in LLC supernatant (k) were analysed (n = 3). l, Assessment of naked tumour volume and weight in subcutaneous tumour models under indicated treatment (n = 6). m, The quantitative analysis of orthotopic lung tumour models at 4 and 12 days (n = 5). n, Body weight measurements of orthotopic tumour models during the administration (n = 6). o, Umap showing re-clustered subgroups in malignant cells. p, Umap showing Ifit1 in MW, PBS and UV group (n = 3). q, The proportion of cells in each population in their respective groups (n = 3). r, WB analysis of the expression of CRT in subcutaneous tumours from mice after the intervention of PBS, UV-TMPs or MW-TMPs (n = 5). s, The statistical analysis of WB results in r. t, The HMGB1 release profile detected with ELISA (n = 5). The data in a–c, f, h–m, s and t are shown in analytic plots as mean ± s.d., and ordinary one-way ANOVA was used for data in a–c, f, h–l, s and t. Two-way ANOVA was used for m and n. The exact P value was noted.
Further exploration into the presence of death receptors or immunomodulatory molecules in MW-TMPs was conducted via proteomic analysis. We identified enriched pathways involving complement and coagulation cascades, epithelial cell apoptotic processes, positive regulation of defence responses and responses to external stimuli, based on differentially expressed protein sets between MW-TMPs and UV-TMPs (Fig. 3d and Supplementary Fig. 4g). Kyoto Encyclopedia of Genes and Genomes (KEGG) showed that the number of proteins annotated to the subclass of immune systems ranked ahead in the organismal systems category (Supplementary Fig. 4h). Notably, HMGB1—a canonical damage-associated molecular pattern (DAMP)—was markedly upregulated in MW-TMPs, ranking 16th in differential expression (Supplementary Fig. 4i). This higher levels of HMGB1 in MW-TMPs were confirmed by cluster analysis (Fig. 3e and Supplementary Table 5), WB and FCM (Extended Data Fig. 2b and Supplementary Fig. 4j). Given HMGB1’s established role in DC and macrophage activation22, we propose that DAMP expression (including HMGB1) in MW-TMPs critically activates these immune cells.
As HMGB1 is a hallmark of ICD23, we proceeded with further evaluations to determine whether HMGB1-enriched MW-TMPs could induce ICD in tumour cells. The results showed that MW-TMP-treated LLC cells exhibited a significant upregulation of ICD markers, including HMGB1, ATP and calretinin (CRT) (Fig. 3f–h and Supplementary Fig. 4k–m). This upregulation was abrogated by HMGB1 inhibitors (Fig. 3i–k and Supplementary Fig. 4n,o). These findings suggest that MW-TMPs induce ICD in tumour cells through the shuttling of HMGB1. Cells undergoing ICD release DAMPs, which trigger a pro-inflammatory cascade, ultimately leading to a robust adaptive immune response and enhanced T cell activation23. Consistently, MW-TMP-pretreated tumour cells enhanced CD86+ bone marrow-derived DC (BMDC) maturation (Extended Data Fig. 2c), elevated CD8+/CD4+ ratios (Extended Data Fig. 2d and Supplementary Fig. 4q) and increased IFNγ secretion, compared with phosphate-buffered saline (PBS)/UV-TMP controls (Extended Data Fig. 2e), indicating the greater immunogenic potential of MW-TMP-treated tumour cells. Notably, the production of IFNγ was significantly inhibited by an HMGB1 inhibitor (Extended Data Fig. 2f), suggesting that the enhanced immunostimulatory capability of MW-TMP-pretreated tumour cells is partly mediated by HMGB1 within the MW-TMPs. Together, these data demonstrated that MW-TMPs directly promoted DC maturation and macrophage reprogramming. In addition, MW-TMPs potentiate the delivery of HMGB1 to tumour cells, thereby inducing ICD and amplifying antitumour immune responses.
Before in vivo ICD efficacy evaluation, we assessed in vivo targeting and biodistribution of MW-TMPs. MW-TMPs exhibited superior targeting efficiency over DSPE-mPEG liposomes (Extended Data Fig. 3a–c and Supplementary Fig. 5a–e). Moreover, the preferential accumulation of LLC-derived TMPs in tumours, compared with MW-MPs derived from noncancerous cells or MW-TMPs from other tumour cells, supports our hypothesis that TMPs inherently target tumour cells of their origin, likely owing to the retained cell-specific surface markers. To further distinguish between active targeting and passive accumulation via the enhanced permeability and retention effect, we utilized a xenograft model in nude mice by subcutaneously implanting A549 tumour cells. Consistent with previous observations, A549-derived MW-TMPs exhibited the highest accumulation in tumour tissues, reinforcing the hypothesis that MW-TMPs actively target tumour cells beyond passive enhanced permeability and retention mechanisms (Supplementary Fig. 5f–i). Our previous studies demonstrated that unmodified UV-TMPs exhibited insufficient tumoricidal activity in vivo9,10. Here MW-TMPs significantly suppressed tumour growth, compared with UV-TMPs, both in subcutaneous (Extended Data Fig. 3d–h and Supplementary Fig. 5j) and orthotopic models (Fig. 3m,n and Extended Data Fig. 3i–l) without systemic toxicity (Supplementary Fig. 5k,l).
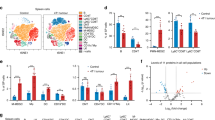
scRNA sequencing on LUAD tissues was carried out (Extended Data Fig. 4a). The canonical transcriptome profiles along with the inferCNV algorithm were used24 (Supplementary Fig. 6a). Subsequent analysis identified several clusters (Extended Data Fig. 4b). We re-organized tumour subsets into eight clusters (Fig. 3o and Supplementary Table 6). Notably, cluster 1_Col5a2, cluster 2_Slc2a1 and cluster 4_Ifit1 exhibited significant variations in their responsiveness to MW-TMP or UV-TMP treatment (Fig. 3q). KEGG analysis showed that MW-TMPs suppressed matrix-associated pathways (cluster 1_Col5a2), amplified metabolic rewiring (cluster 2_Slc2a1) and elevated interferon-stimulated genes (cluster 4_Ifit1) (Fig. 3p, Extended Data Fig. 4c and Supplementary Fig. 6b), suggesting that both interventions have the potential to drive interactions between tumour cells and the immune system. Further ligand–receptor analysis highlighted dominant MIF_CD74_CD44 and Ccl2–Ccr2 interactions (Supplementary Table 7). MW-TMPs upregulated tumour-T cell major histocompatibility complex (MHC)-I signalling while downregulating tumour-endothelial matrix crosstalk (Extended Data Fig. 4d). Hallmark gene set enrichment analyses revealed that pathways associated with the inflammatory response and other antitumour signalling were upregulated in the MW-TMP group (Supplementary Fig. 6c). Moreover, enrichment of ICD markers (Anxa1, Hsp90aa1, Hmgb1 and Cxcl10) and apoptosis genes (Casp3) corroborated ICD induction (Fig. 3r–t and Extended Data Fig. 4e–h). Collectively, our findings revealed that MW-TMPs enriched with HMGB1 can treat tumour by inducing ICD effect.
MW-TMPs activate DCs for enhanced antigen presentation and reprogramme TAM to antitumour phenotype in the TME
Analysis of immune cell infiltration revealed elevated T cell proportions in UV-TMP- and MW-TMP-treated groups compared with PBS controls (Fig. 4a). Given the pivotal role of DCs in initiating and regulating innate and adaptive immune responses, we initially focused on the intratumoural DC subpopulations. With reference to the literature24,25, DCs were classified into three subtypes, including conventional DC type 1 (cDC1), conventional DC type 2 (cDC2) and mregDC (Fig. 4b and Extended Data Fig. 5a). cDC1 and cDC2 have been previously reported to activate CD8+ T cells and CD4+ T cells, respectively25. The mregDC was described as ‘mature DCs enriched in immunoregulatory molecules’ owing to the expression of immunoregulatory genes and maturation genes25,26. These cells have been reported to restrain DC immunostimulatory function and control the threshold of T cell activation25. Trajectory analysis revealed that mregDCs likely originated from both cDC1s and cDC2s, aligning with recent reports that mregDCs are derived from cDC1s and cDC2s upon uptake of tumour antigens25,26 (Fig. 4c and Extended Data Fig. 5b).

a, The proportion analysis of CD45+ cells in the three groups (n = 3). b, Umap plot of DCs coloured by clusters. c, The developmental trajectory analysis of DCs. d, The proportion of the DC subtypes in PBS, UV-TMP and MW-TMP groups. e, Cell-to-cell communication analysis of cDC1s, cDC2s and mregDCs. f, Representative immunofluorescence graphs of tumours, spleens and lymph nodes (n = 5). Green, CD11c; red, CD86. Scale bars, 20 μm. g,h, DC infiltration and the ratios of CD86+ DCs (g) and CD80+ DCs (h) in tumours via FCM (n = 5). i, Distribution of CD8a+ cDC1s in tumour tissues by FCM (n = 5). j, The proportion of MHCII in CCR7+ mregDCs by FCM in tumour sites (n = 5). MFI, mean fluorescence intensity. k, Umap plot of monocytes/macrophages coloured by clusters among PBS, UV-TMPs and MW-TMPs. l, The proportion of six subgroups in specific groups. m, M0, M1 and M2 signature scores based on the gene signatures from CIBERSORT. n, The developmental trajectory analysis of monocytes/macrophages. o, The specific marker gene expression in developmental trajectory. p,q, The proportions of M1-like (p) and M2-like (q) macrophages by FCM in tumour sites (n = 5). The data in a, g–j, p and q are shown in analytic plots as mean ± s.d., and ordinary one-way ANOVA was used for data in g–j, p and q. Two-way ANOVA was used for a. The exact P value was noted.
Following MW-TMP therapy, the fractions of cDC1s significantly increased, indicating the activation of DCs (Fig. 4d). Of note, MW-TMP treatment resulted in a significant elevation in the proportion of mregDCs. This finding reminisces us of a previous study that identified the HMGB1 peptide as a potential immunoadjuvant, since it boosted the number of intratumoural mregDCs and enhanced their MHC class I antigen-presenting programme27. Thus, we analysed the antigen-presenting gene expression on mregDCs, and the results showed that the MW-TMP treatment enhanced the expression of MHC class I antigen presentation-related genes (Extended Data Fig. 5c). We also found higher levels of mregDCs’ capability in communication with intratumoural T cells upon MW-TMP treatments (Fig. 4e and Supplementary Fig. 6d). Collectively, MW-TMP elevates the abundance of intratumoural cDC1s and mregDCs, concurrently potentiating the expression of MHC class I antigen presentation programme in mregDCs.
Further immunofluorescence revealed enhanced DC activation in tumours, spleens and lymph nodes in MW-TMP groups (Fig. 4f). FCM analysis of single-cell suspensions from tumour tissues revealed that the general percentages and maturation ratios of DCs in tumour tissues increased upon MW-TMP treatment (Fig. 4g,h, Extended Data Fig. 5d and Supplementary Fig. 7a,b). CD8a and CD103 were used to identify resident and migratory cDC1s, respectively27,28. Subset analysis demonstrated that MW-TMPs preferentially expanded resident cDC1s over migratory cDC1s (Fig. 4i, Extended Data Fig. 5e and Supplementary Fig. 7c,d). As reported, CCR7+CD11b+CD11c+Ly6C−MHC class II+ cell populations were identified as CCR7+ mregDCs28. Increased intratumoural CCR7+ mregDCs were observed in tumours treated with MW-TMPs (Extended Data Fig. 5f and Supplementary Fig. 7e). Furthermore, CCR7+ mregDCs in the MW-TMP treatment group exhibited higher MHCII and CD80, indicating greater immunostimulatory and antigen presentation capability (Fig. 4j and Extended Data Fig. 5g). Overall, these findings suggest that DCs are efficiently activated and recruited following MW-TMP therapy, potentially contributing to the function of cytotoxic T cells.
Parallel macrophage subclustering identified six functionally distinct subsets (Fig. 4k and Extended Data Fig. 6a). Unsupervised clustering revealed distinct macrophage subset distributions across treatment groups (Fig. 4l). Ccl8_macro and Saa3_macro were predominantly found in PBS-treated mice, whereas Cxcl9+ macrophages and Fn1+ macrophages were abundant in both UV-TMP- and MV-TMP-treated mice. Both Cxcl9_macro and Fn1+_macro exhibited augmented inflammatory responses, coupled with enhanced interferon responses (Ifnb1, Isg15, Ifit1, Rsad2 and Ifit2) and robust antigen presentation capabilities (H2-ab1, H2-eb1, H2-aa and Cd74), respectively. By contrast, Ccl8_macro exhibited characteristics of alternative activation (Folr2, Cd163, Cd209 and Mrc1) and tolerance to T cell genes (Tgfbr2). Saa3_macro, despite harbouring seemingly conflicting genes related to inflammation (Il1b, Tnf, Nlrp3 and Ptgs2) and angiogenesis (Thbs1, Ecm1, Lrg1, Il1b, Thbs1 and Hif1a), have been implicated in driving pathogenic inflammation that contributes to pancreatic ductal adenocarcinoma progression29.
Macrophages are traditionally classified into three subtypes: M0 (non-polarized or neutral), M1 (pro-inflammatory or antitumour) or M2 (anti-inflammatory or pro-tumour)30. Module scoring confirmed M1 enrichment (Cxcl9_macro/Fn1_macro) and M2 reduction (Ccl8_macro) post-MW-TMPs (Fig. 4l,m and Supplementary Table 8). Trajectory analysis determines M1-biased positioning of Cxcl9/Fn1_macro and M2-biased clustering of Ccl8_macro, with Spp1/Saa3_macro distributed across M0–M1–M2 branches, indicating the great function flexibility (Fig. 4n). Furthermore, the trajectory heat map revealed sequential gene expression changes (Extended Data Fig. 6b,c). Clearly, gradually upregulated or downregulated expression of genes was shown during the switch from M2 to M1 (Fig. 4o and Extended Data Fig. 6d). The MW-TMP group exhibited a higher score in M1 rather than M2 compared with the PBS group (Extended Data Fig. 6e). FCM corroborated increased M1-like tumour-associated macrophages (TAMs) and reduced M2-TAMs/pMDSCs in MW-TMP-treated tumours, without altering mMDSCs or neutrophil subsets (Fig. 4p,q and Supplementary Fig. 7f,g). These findings indicate that MW-TMP promotes pro-inflammatory macrophage expansion and TAM reprogramming towards an antitumour phenotype while reducing immunosuppressive pMDSCs, thereby alleviating the immunosuppressive tumour microenvironment (TME).
MW-TMPs promote cytotoxic T/NK cell expansion
Next, we re-clustered CD8+ T and NK cells into three subtypes of CD8+ T cells and two subtypes of NK cells (Fig. 5a,b). Among three CD8+ T cell subclusters, CD8_c1 demonstrated higher expression of effector molecules (Ifng, granzyme B (Gzmb) and Nkg7), exhaustion markers (Havcr2, Pdcd1, Lag3, Ctla4 and Entpd1) and other canonical genes (Cx3cr1, Ki67 and Tbx21), indicative of a phenotype characteristic of proliferating but transitory exhausted T cells. CD8_c2 showed stem-like properties marked by TCF7, Sell, Cxcr3 and Lef1. CD8_c3 exhibited characteristics associated with T cell differentiation, including the expression of Anxa1, Vim, bx3 and Srm genes, coupled with elevated levels of the effector phenotypes (Ccl2, Ccl3, Ccl4 and Csf1), representing CD8+ effector/memory T cells. Gene signature scoring revealed the prominently high activation/inhibitory score of CD8_c1, indicating its distinct activation and inhibitory characteristics. CD8_c3 shows a relatively elevated memory/differentiation score along with robust effector function signatures, suggesting its memory-like properties and effector functions. Conversely, CD8_c2 demonstrates the lowest scores across all three subclusters, emphasizing its unique yet relatively subdued characteristics (Fig. 5c and Supplementary Table 8). Monocle2 trajectory analysis of CD8+ T cells identified bifurcating differentiation trajectories originating from CD8_c3 effector/memory/stem-like cells and terminating in exhausted/effector states, confirming sequential activation–exhaustion dynamics (Fig. 5d).

a, The umap plot of NK and T cells. b, Heat map of normalized expression of NK and T cell classical genes among six clusters. TFs, Transcription factors. c, The gene signature scores of memory/differentiation, activation/inhibitory, effector and transcription factor expression in CD8-c1, CD8-c2 and CD8-c3. d, The developmental trajectory analysis of CD8+ T cells. e, The distribution of CD8-subtypes among groups treated with respective intervention. f, The cytotoxic and exhausted gene signature scores among PBS, MW-TMP and UV-TMP groups (n = 3). g, Ratios of CD8+ T, CD4+ T cells in tumours via FCM (n = 5). h, Representative immunofluorescence graphs of tumours, spleens and lymph nodes (n = 5). Green, CD8; red, CD4. Scale bars, 20 μm. i, The proportions of Treg cells in tumours (n = 5). j,k, The proportions of Th1 (j) and Th2 (k) cells in tumours (n = 5). l,m, Cytotoxic CD8+ T cells in tumours secrete higher percentages of Gzmb (l) and IFNγ (m) with the treatment of MW-TMPs (n = 5). n, The umap plot of cytotoxic effector genes coloured by cluster in the NK group. o, The cytotoxic and exhausted gene signatures for the two NK subclusters (n = 3). p, The proportion of NK subgroups in the untreated and treated groups (n = 5). q, The proportions of total NK cells and activated subgroups (IFNγ+ NK) in tumours (n = 5). The data in f, g, i–m, o and q are shown in analytic plots as mean ± s.d., and ordinary one-way ANOVA was used for data in f, g, i–m, o and q. The exact P value was noted.
We observed a higher fraction of CD8_c1 in both UV-TMP and MW-TMP groups relative to the PBS group (Fig. 5e). Recent studies have indicated that these experienced T cells are specifically derived from tumour-specific cytotoxic T cells31, correlated with immune checkpoint blockade therapy response32. Notably, MW-TMP preferentially expanded CD8_c3 effector/memory populations, while UV-TMP significantly induced CD8_c2 stem-like cycling T cells, indicating distinct responses to the different treatment modalities (Fig. 5e). Moreover, the cytotoxic gene signature scores of CD8_c3 were higher after MW-TMP treatment, suggesting the enhanced and plastic tumour killing effect of this subcluster cells (Fig. 5f). FCM and immunofluorescence demonstrated MW-TMP-induced systemic CD8+ T cell activation in lymphoid organs, suggesting the effective elicitation of a systemic immune response, which was required for tumour rejection33 (Fig. 5g,h). Functional profiling revealed that MW-TMP significantly increased Gzmb/IFNγ production in CD8+ T cells and reduced Treg frequencies, in contrast to PBS/UV-TMP groups via FCM (Fig. 5i–m). Taken together, these data demonstrate that MW-TMP intervention promotes APC sensing of tumour antigen and cross-priming of tumour-specific CD8+ T cells and expands the intratumoural effector/memory T cell population.
NK cells, a crucial component in the TME, regulate tumour control through direct killing and pro-inflammatory cytokine secretion34. Transcriptomic profiling characterized two functionally distinct NK subsets: NK_FCGR3 cells showed elevated expression of cytotoxic effectors, including perforin (Prf1) and granzyme family members most granzyme (Gzmb, Gzmg, Gzmf, Gzmd, Gzme and Gzmc), whereas NK_SELL cells retained moderate cytotoxicity but were enriched in naive-associated markers (Tcf7/Sell/Lef1) (Fig. 5b,n and Supplementary Table 9).
Since NK_FCGR3 cells were reported to be the most representative of cytotoxic cells35, we further calculated the cytotoxic and exhausted gene signatures for the two NK subclusters (Fig. 5o and Supplementary Table 8). As expected, NK_FCGR3 cells showed higher cytotoxic signatures than NK_SELL cells. MW-TMP treatment selectively expanded NK_FCGR3 populations, while UV-TMP preferentially induced NK_SELL expansion, demonstrating MW-TMP-driven enhancement of NK cytolytic function (Fig. 5p). This functional reprogramming was corroborated by elevated cytotoxic signatures in MW-TMP-treated NK cells (Fig. 5o) and validated through FCM, which revealed both increased NK cell infiltration and enhanced IFNγ production following MW-TMP administration (Fig. 5q). Collectively, these findings underscore the ability of MW-TMP to promote the expansion and activation of cytotoxic T and NK cells, thereby enhancing antitumour immune responses.
MW-TMPs can treat MPE, serve as carriers and enhance PD-L1 immunotherapy efficacy
To explore the clinical potential of MW-TMPs, we collected clinical MPEs, during thoracentesis from patients with confirmed MPE cytology (Fig. 6a). FCM analysis revealed that approximately 30% of MPE cells were tumour cells, and 60% were diverse immune cells, dominated by immunosuppressive monocytes/macrophages and neutrophils with scarce T lymphocytes, corroborating previous observations that MPEs generally have an ‘immune-cold’ profile36 (Fig. 6b).

a, Schematic illustration of the treatment process of MPE from patients with lung cancer, created in BioRender. Y, Z. (2025) https://BioRender.com/lgc5c9p. b, FCM analysis of the cell composition within human MPE (n = 5). c, Representative immunofluorescence images showing uptake of autologous MW-MPs (n = 5). Scale bars, 100 μm for the wider perspective. Scale bars, 50 μm for the smaller perspective. d, The proportion of cells taking up autologous MW-MPs in MPE (n = 5). e, Cellular composition of cells taking MPs (n = 6). f, WB analysis of the expression of CRT (n = 5). g, The concentrations of ATP in the supernatant of MPE cells (n = 3). h–o, The ratios of CD80+ DCs (h), the proportions of M1-like (i), the ratios of M1/M2 (j), the cytokine levels of Foxp3 (k) and IFNγ (l) in CD4+ T cells, the proportions of CD69 (m) and CD25 (n) in CD8+ T cells in MPE and the levels of NKG2D in NKT cells (o) (n = 5). p, The representative illustration of 3D spherical models constructed by MPE cells (n = 3). Scale bar, 200 μm. q, The representative images of the spherical size after 6-day stimulation and live/dead fluorescence staining of 3D spherical models (n = 3). Scale bars, 200 μm. r, The volume curve of 3D spherical models (n = 3). s, The statistical analysis of live/dead fluorescence staining of 3D spherical models (n = 3). t, Schematic illustrating the establishment and treatment of zebrafish PDX models, created in BioRender. Y, Z. (2025) https://BioRender.com/0g7gooc. u, The representative images and fluorescence intensity analysis of tumour cells in PDX models (n = 10). Scale bar, 50 μm. The data in d–q, t, u and w are shown in analytic plots as mean ± s.d., and ordinary one-way ANOVA was used for data in d, f–q and u. Two-way ANOVA was used for e and t. The exact P value was noted.
Tumour cells from MPE pallets were isolated for the preparation of the MW-TMPs and UV-TMPs. Autologous MPE co-culture assays demonstrated preferential MW-TMP uptake by tumour cells, followed by neutrophils and macrophages (Fig. 6c–e). Building on in vitro evidence of MW-TMP-induced ICD, human MPE models demonstrated superior induction of ICD markers (CRT and ATP) by MW-TMPs compared with UV-TMPs, suggesting their immunogenic potential (Fig. 6f,g). Ex vivo MW-TMP treatment of MPE cells enhanced DC maturation (Fig. 6h and Supplementary Fig. 7a) and promoted macrophage repolarization (Fig. 6i,j and Supplementary Fig. 7b). Although T cell infiltration showed non-significant increases (Supplementary Fig. 7c,d), MW-TMP treatment significantly reduced Treg populations while elevating activated T cell subsets (IFNγ+CD4+, CD25+CD8+ and CD69+CD8+) and NKG2D+ NKT cells (Fig. 6k–o).
To directly assess tumour suppression potential of MW-TMPs, we developed a three-dimensional (3D) multicellular spheroid model recapitulating the solid TME using fresh MPE cells (Fig. 6p). MW-TMPs significantly inhibited spheroid growth versus controls (Fig. 6q) and demonstrated enhanced cytotoxicity over UV-TMPs (Fig. 6q–s). The therapeutic potential of MW-TMPs was further validated in zebrafish PDX models derived from resected human lung tumours (zebrafish PDX models; refs. 37,38). In this model, MW-TMPs demonstrated effective antitumour effects (Fig. 6t,u). Together with findings from our in vitro pleural fluid 3D model, subcutaneous model and in situ model, these findings confirm the robust antitumour capabilities of MW-TMPs, thereby reinforcing our conclusions and paving the way for potential clinical applications.
We explored the drug-loading capacity of MPs. Our study demonstrates that MTX-loaded MW-TMPs exhibit effective drug encapsulation and sustained release kinetics, with cumulative release rates of 8% and 32% at 4 h and 24 h, respectively (Extended Data Fig. 7a and Supplementary Fig. 9a,b). In vitro assays revealed that MW-TMPs-MTX significantly suppressed LLC proliferation through enhanced apoptosis (Extended Data Fig. 7b,c), while in vivo administration resulted in marked tumour growth inhibition in LUAD-bearing mice without significant body weight alterations compared with PBS or blank MW-TMP groups, reinforcing the potential of MW-TMPs as a drug delivery system, similar to other EVs (Extended Data Fig. 7d–g). Notably, MW-TMPs synergized with anti-PD-L1 antibody therapy to enhance antitumour responses beyond monotherapy or PBS controls, while maintaining systemic safety throughout treatment (Extended Data Fig. 7d,h–j). These findings collectively establish MW-TMPs as a dual-functional nanoplatform integrating drug delivery with immunotherapeutic enhancement, demonstrating significant translational value in oncology.
Conclusion
MW irradiation offers a convenient, cost-effective and high-efficiency technique for TMP preparation. MW-TMPs exhibit remarkable antitumour efficacy as a monotherapy and serve as carriers for the anticancer agent MTX, enabling targeted tumour cell elimination. Moreover, combining MW-TMPs with anti-PD-L1 immunotherapy synergistically enhances therapeutic outcomes. Mechanistically, MW-TMPs induce a dual immunologic attack mediated by T cells and NK cells, attributed to their direct immunostimulatory effects and the ability to induce ICD in tumour cells through the horizontal transfer of HMGB1. Future studies on the MW-TMP concept are anticipated to explore its therapeutic potential and potentially translate it into clinical applications.
Methods
Cell lines
Murine LLC cell line (#CRL-1642) was purchased from the American Type Culture Collection. B16-F10 cell line was a gift from J. Tao of the Department of Dermatology, Wuhan Union Hospital, Tongji Medical College, Huazhong University of Science and Technology. Subsequently, LLC cells were transfected with the pcDNA3-OVA plasmid (Genecreale) to obtain LLC-OVA cells. Mouse T cell hybridoma cells B3Z (#BFN608006444) were purchased from Bluefbio Biology Technology Development. Murine lung epithelial cells (MLE, #iCell-m036), murine colon adenocarcinoma cell line (MC38, #iCell-m032) and human bronchial epithelioid cells (BEAS-2B, #iCell-h023) were purchased from the Cellverse. Murine embryonic fibroblast cell line (NIH/3T3, #CL-0171) was purchased from the Pricella. These cell lines were cultured in DMEM (Gibco) supplemented with 10% FBS (Newzerum) at 37 °C in 5% CO2–95% air.
Mice
C57BL/6 mice were purchased from Hubei Beiente Biotechnology. They were kept in specific pathogen-free facility in the Wuhan Laboratory Animal Center of Tongji Medical College. All animal-related procedures have been performed according to the Declaration of Helsinki and gained permission from the Animal Care and Use Committee of Tongji Medical College ((2023) IACUC number 3958).
Preparation and isolation of MW-TMPs
We established an MW-assisted method to collect TMPs from tumour cells. After 24 h of incubation under standard conditions, the cell culture medium was replaced with a serum-free medium. The Petri dishes were subsequently placed into a domestic MW oven (setting condition 700 W, Midea) and subjected to MW heating for 20 s, a condition we designated as H20. To compare the products from different MW conditions, several settings were applied to LLC cells: L10 (175 W, 10 s), L20 (175 W, 20 s), L30 (175 W, 30 s), H10 (700 W, 10 s), H20 (700 W, 20 s) and H30 (700 W, 30 s). After 48 h of culture, the cell-conditioned medium from each MW treatment was collected for isolation of MW-TMPs. TMPs were extracted according to our previous study via a serial centrifugation strategy9. In brief, the medium was centrifuged at 200 × g for 10 min and 2,000 × g for 30 min at 4 °C to remove cells and debris, respectively. Second, pellet was obtained after 18,000 × g for 60 min at 4 °C and then resuspended in PBS. Finally, MW-TMPs were washed once and then stored using PBS. For drug-loaded MW-TMPs, the incubation method was used to encapsulate MW-TMPs with small molecules, such as MTX. After MW treatment, 200 μM MTX was added into the culture medium and the extraction of MW-TMPs-MTX followed the aforementioned procedures.
TMT quantification proteomics
UV-TMPs were prepared and isolated based on our former protocols10. Abundant proteins of UV-TMPs (named as U1, U2 and U3) and MW-TMPs (named as M1, M2, M3 and M4) as Mus_musculus samples were identified via single mass spectrometry run using the TMT quantitative method. Totally, 747,975 spectrums were generated, 19,595 peptides and 3,967 proteins were detected with 1% false discovery rate, and the further protein annotation with the help of multiple bioinformatic databases, including KEGG and Gene Ontology, was carried out. For differential protein analysis, an automated software, named IQuant, was applied and proteins with 1.5-fold change and P < 0.05 were regarded as differential expression12.
High-performance liquid chromatography of MW-TMPs-MTX
MW-stimulated LLC cells were incubated with 50 μM, 100 μM, 200 μM and 300 μM MTX for 24 h. MW-TMPs-MTX was collected and resuspended in ddH2O. Samples were sonicated for 10 min and passed through a 0.2 μm water film. As for the cumulative MTX release analysis by MW-TMPs, prepared MW-TMPs-MTX were incubated in the environment of pH = 7.4 and 37 °C and the stirring of 200 rpm at intervals of 4 h, 6 h, 12 h and 24 h. Then high-performance liquid chromatography was utilized to detect the drug content in MW-TMPs. Mobile phase (phase A, 0.025 mol l−1 potassium dihydrogen phosphate buffer (pH 5.5); phase B, acetonitrile) with the liquid samples (MTX or MW-TMPs-MTX) went through the chromatographic column (Athena-C18, temperature, 30 °C; flow rate, 1.0 ml min−1) under 303 nm wavelength in the model of isocratic elution. According to the chromatograms of standards and samples, the concentration of MTX loaded in MW-TMPs-MTX was analysed. Drug loading efficiency was evaluated with the ratio of MTX package quality and TMP quality39.
Detection of the ICD biomarkers
LLC cells were treated with either 20 μg ml−1 MW-TMPs, UV-TMPs or PBS, respectively. In addition, some LLC cells were pretreated with the HMGB1 inhibitor (glycyrrhizic acid, #HY-N0184) before MW-TMPs stimulation. The expression of canonical ICD markers, including HMGB1, ATP and calreticulin (CRT), was detected 24 h post-stimulation. HMGB1 expression levels were analysed by the WB assay (Supplementary Table 2). HMGB1 concentrations in the cell supernatant were measured using an ELISA kit (Bioswamp, catalogue number MU30043) following the manufacturer’s guidelines. CRT expression was evaluated by FCM and visualized through immunofluorescence (Supplementary Table 4). ATP content in the LLC supernatant was quantified using an ATP assay kit (Beyotime, catalogue number S0026).
MW-TMPs fluorescence labelling and uptake in vitro and in vivo
MW-TMPs were labelled with lipophilic membrane tracers DiI (Beyotime, #C1036), DiO (Beyotime, #C1038) or DiR (MedChemExpress, #HY-D1048) referring to the manufacturer’s instruments. After 10 min incubation in darkness at room temperature, MW-TMPs were washed using PBS twice to remove free dye solutions.
Fluorescence-labelled MW-TMPs were added to tumour cells. At various incubation intervals, cell medium was removed and PBS was used to wash cells twice. Cell nuclei were stained with DAPI and then the internalization of MW-TMPs was assessed by a fluorescence digital scanner (3DHISTECH, Pannoramic SCAN) or FCM (BD LSRFortessa X-20).
Female subcutaneous tumour-bearing C57BL/6 mice were intraperitoneally injected with DiR-labelled MW-TMPs. After post-administration of 1 h, 12 h and 24 h, the presence of MW-TMPs in tumour sites and freshly removed vital organs were visualized under bioluminescence imaging (Bruker MS FX Pro Imaging System). Excitation and emission wavelengths were set at 600 nm and 570 nm, respectively.
Immune cell extraction and induction
Murine femurs from C57BL/6 mice (6–8 weeks) were dissociated under sterile conditions, followed by immediate centrifugation at 12,000 rpm for 1 s to isolate bone marrow cells. The cells were treated in 1 ml red blood cell lysis buffer for 3 min and then neutralized with RPMI-1640 medium supplemented with 10% FBS. The cell suspension was then centrifuged at 1,800 rpm, 5 min at 4 °C. For induction of BMDCs, primary bone marrow cells were cultured in RPMI-1640 medium supplemented with 10% FBS, 20 ng ml−1 interleukin-4 (IL-4) (Peprotech, #214-12) and GM-CSF (Peprotech, #315-03) for 1 week. In addition, BMDMs were induced by culturing bone marrow cells in RPMI-1640 medium supplemented with 10% FBS and 20 ng ml−1 M-CSF (Peprotech, #315-02) for 1 week.
Lymphocytes were extracted from the spleens of 6–8-week-old C57BL/6 mice using the mouse CD3+ T cell isolation kit (Vazyme, catalogue number CS101) according to the manufacturer’s instructions. The isolated spleen lymphocytes were incubated in RPMI-1640 medium supplemented with 10% FBS (Gibco), 20 ng ml−1 IL-2 (Peprotech), HEPES solution (100×, Procell), β-mercaptoethanol (1,000×, Procell) and sodium pyruvate solution (100×, Procell).
To assess the immune-stimulating capacity of tumour cells treated with MW-TMPs, LLC tumour cells were treated with LLC-derived MW-TMPs for 24 h. These resulting LLC cell pellets were then co-cultured with BMDCs for another 24 h. Subsequently, these pretreated BMDCs were co-cultured with either B3Z cell line or isolated CD3+ T cell for 48 h, allowing for the evaluation of the proportion of mature DCs, CD4+ T, CD8+ T cells, as well as the secretion of IFNγ (#1210002, Dakewe) and IL-2 (#1210203, Dakewe).
Subcutaneous and orthotopic tumour-bearing model and in vivo treatment
LLC cells (5 × 105 cells suspended in 100 μl DMEM solution) were subcutaneously inoculated into the unilateral posterior flanks of C57BL/6 mice (6 weeks). After approximately 1 week of observation, they were randomized into several groups (n = 6). The tumour volume and body weight were recorded every other day until the end of the intervention. For lung orthotopic tumour models, 6 × 105 LLC-LUC cells were injected into the exposed lung tissue of mice. After 3 days, the orthotopic tumour models were randomly classified into three groups and give specific intervention. The growth of orthotopic tumour was monitored by using the IVIS Spectrum imaging system to detect the spontaneous luminescence formed after d-luciferase binds to the substrate. The treatment (100 μl PBS, 3 μg g−1 MPs, 5 μg g−1 anti-PD-L1 antibody) was administrated intraperitoneally once every 2 days. Mice were sacrificed after the fifth intervention. Subcutaneous or naked tumour volume was estimated as follows: volumeSubcutaneous tumour = length × width2 × 0.5; volumeNaked tumour = length × width × height × 0.5. To estimate the antitumour efficiency, the formula about tumour growth inhibition rate (TGI %) was used: TGI (%) = (1-mean volumetreatment/mean volumecontrol) × 100%. According to the ethics committee’s regulations, the maximal tumour burden permitted is 2,000 mm3. We confirm that the maximal tumour burden was not exceeded during the study.
scRNA sequencing
When the intervention was completed, fresh subcutaneous tumours were stored in magnetic-activated cell sorting tissue storage solution (#130100008) and cell suspensions were prepared by tissue homogenate. The single-cell suspension was added to the single-cell sorting honeycomb plate and excessive magnetic beads with barcodes were put into the plate. During the phase of cell lysis, the RNA was labelled as bounding to the barcode. Subsequently, reverse transcription of RNA and cDNA synthesis were carried out to obtain cDNA library for subsequent sequencing. Paired-end sequencing was conducted in the NovaSeq 6000 sequencing platform to get the transcriptome results. BD Rhapsody Analysis pipeline was applied to identify the UMI sequence and the cell tag sequence alignment to the reference genome (GRCm39). After removing low-quality cells, the normalization methods ‘LogNormalize’ and ‘ScaleData’ were used to standardize and scale each gene and then linear dimensional reduction was performed according to the PCA score. Uniform manifold approximation and projection (UMAP) techniques were chosen to visualize the cell clusters (the visualized PCA dimension = 38). The raw scRNA sequencing datasets have been deposited in the NCBI database (GSE289959).
Pseudotime trajectory analysis and calculation of signature score
According to differential expression genes across cells, monocle package (version 2.26.0) was used for analysing the trajectory processing of DCs, CD8+ T cells and TAMs to infer their dynamic differentiation and functions. To quantify the function of immune cells, including DCs, CD8+ T cells and NK cells, the ‘AllModuleScore’ function (Seurat version 4.4.0) was used to calculate the enrichment scores according to signature gene sets (Supplementary Table 8).
Cell-cell interaction and pathway enrichment analysis
We used the classic ‘CellChat’ package (version 1.6.1) to predict and analyse the strength of interactions between different cell types, mainly including tumour cells and various immune cells, as well as DC subpopulations and T cells, based on cell gene expression. After 1,000 pairing tests, ligand–receptor pairs with significant enrichment and P-value significance were extracted (Supplementary Table 7). On the basis of the differential expression genes among treated and untreated groups by performing ‘FindAllMarkers’, KEGG and hallmark pathway analysis were investigated to estimate the differential activities of cells comparing groups.
Distribution of tumour-infiltrating immunocytes
The detection of immune cells in tumour tissues using FCM was carried out as previously described10. In brief, single-cell suspension was collected through grind, digestion, filtration and lysis of red blood cells. Cells may need stimulation by PMA (0.1 μg ml−1, Solarbio, P6741-1 mg) and ionomycin (1 μg ml−1, Aladdin, 1139530-1 mg) before evaluation of concentrations of Gzmb, IFNγ and IL-4. Suspension was blocked by anti-mouse CD16/32 antibody. Next, multiple fluorescent antibodies (Supplementary Table 3) were stained in darkness for 30 min at 4 °C. Finally, FCM instrument (Beckman Coulter, DxFLEX) was used to analyse the infiltration of immune cells in the TME.
Tissue in situ immunofluorescence
Tissues of tumours, spleens and lymph nodes were acquired at the end of intervention and washed with PBS before they were fixed and embedded in paraffin. Then the sections underwent dewaxing, hydration, antigen retrieval and quenching of endogenous peroxidases. The tissues were blocked with 2% bovine serum albumin (BSA) and then incubated with fluorescence-coupled antibodies (Supplementary Table 4) overnight at 4 °C. Next, nuclei in tissues were stained with DAPI. Finally, the sections were scanned using a fluorescence microscope scanner.
Acquisition and treatment of human MPE samples
Human MPE samples were obtained from lung cancer patients undergoing thoracentesis to alleviate fluid accumulation at Wuhan Union Hospital, Tongji Medical College, Huazhong University of Science and Technology. The study protocol was approved by the ethical committee of Tongji Medical College, Huazhong University of Science and Technology and all MPE samples were collected with patients’ informed consent ((2019) IEC (S1041)).
Approximately 500 ml of fresh MPE samples per patient were subjected to centrifugation at 1,000 × g for 15 min, resulting in separation into supernatants and cell pellets. A portion of the cell pellets was immediately labelled with FCM antibodies to characterize the cellular composition within the pleural effusion. A list of the anti-human antibodies used is summarized in Supplementary Table 3. All antibodies for human FCM were used at a 1:100 dilution. Another portion of the cell pellets was seeded into agarose gel-coated 96-microwell plates for the subsequent formation of 3D multicellular spheroids. After 5 days, we used 3D multicellular spheroids that formed in the wells to represent a human solid TME for further immunomodulation experiments with various TMP types. The formed 3D multicellular spheroids were treated with various TMPs for 6 days, and spheroid sizes were recorded. At the end point of observation, we immobilized 3D multicellular spheroids with 4% paraformaldehyde and stained them with DAPI for the subsequent imaging. In addition, cytotoxicity assays were conducted using Live/Dead detection kits on the multicellular spheroids to evaluate their viability at the end of the experimental observation.
The remaining cells underwent treatment with red blood cell lysis buffer to remove erythrocytes, followed by washing twice with PBS. Tumour cells were then isolated by magnetic-activated cell sorting using magnetic beads conjugated with human Epcam microbeads (Miltenyi Biotec) following the manufacturer’s instructions. The sorted tumour cells were subjected to either UV or MW stimulation for the preparation of UV-TMPs and MW-TMPs. Simultaneously, a smaller fraction of the whole-cell pallets was seeded into different 12-well plates for analyses of cell uptake of MW-TMPs and immune activation detection.
To assess cellular uptake capacity of TMPs by various cell types within the MPE, Dil-labelled MPs were incubated with MPE pallets in the 12-well plates. After a 24 h incubation period, FCM was used to quantify the phagocytosis levels of Dil-MW-TMPs by individual cell types. Alternatively, some cell pellets were fixed for further immunofluorescence.
To further explore the impact of UV-TMPs and MW-TMPs on immune cells, they were introduced to additional 12-well plates. Upon 24 h of incubation, FCM was utilized to evaluate cell proportion and functional changes in the MPEs from each patient. Concurrently, the cell supernatant was collected to measure ATP concentrations using an ELISA kit. Another portion of the cells was lysed and denatured to facilitate the detection of CRT expression via WB.
Zebrafish PDX models
To establish the PDX models in zebrafish, lung cancer tumour tissues obtained from the patient during the surgical operation were processed into single-cell suspensions using mechanical and enzymatic digestion. These cells were subsequently labelled with Vybrant CM-DiI (Thermo Fisher Scientific). In parallel, peripheral blood samples from the patients were collected to isolate peripheral blood mononuclear cells (PBMCs), which were activated in vitro and stained with DiO fluorescent dye (Thermo Fisher Scientific). The labelled tumour cells and PBMCs were mixed in a 1:1 ratio and microinjected into the yolk sacs of 2 days post-fertilization (dpf) wild-type AB zebrafish from Hunter Biotech, at a density of 300 cells per fish. Then the zebrafish were all reared in fish-rearing water at 35 °C. Regarding the water quality, 200 mg of instant sea salt was added to each 1 l of reverse osmosis water, with the electrical conductivity ranging from 450 μS cm−1 to 550 μS cm−1; the pH value was between 6.5 and 8.5; and the hardness was 50–100 mg l−1 CaCO3. The license number for the utilization of laboratory animals is SYXK (Zhejiang) 2022-0004.
At 3 dpf, zebrafish showing consistent tumour cell expression were selected under the microscope and randomly divided into 3 groups for culture in 6-well plates with a 3 ml well volume, with 10 zebrafish per group. A549-MW-TMPs or A549-UV-TMPs were administered into the yolk sacs of zebrafish models, alongside a model control group. Two days after the intervention, fluorescence intensity was assessed by photographing 10 zebrafish per group under a fluorescence microscope. The images were processed using NIS-Elements D 3.20 advanced image-processing software to evaluate the antitumour growth efficacy of TMPs.
Statistical analysis
The experiments in the study were repeated independently at least three times. Statistical analysis was performed using the software GraphPad 8.0. All data were shown as mean values ± s.d. Significant comparison between two groups was determined through unpaired two-tailed Student’s t-test. For more than two groups, one-way analysis of variance (ANOVA) with further multiple comparisons using post-Turkey’s multiple comparison test was carried out. P < 0.05 indicated statistical significance.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data supporting the conclusions of this study are presented herein and in the Supplementary Information. The raw single-cell RNA sequencing datasets have been deposited in the NCBI database (GSE289959). The transcriptomic data have been identified by the accession code GSE292405. Owing to the substantial size and volume of the remaining raw data, these materials are available from the corresponding authors upon reasonable request. Source data are provided with this paper.
Code availability
All custom code used in this study is freely accessible from the corresponding authors upon reasonable request.
References
Zhang, M. et al. Engineered exosomes from different sources for cancer-targeted therapy. Signal Transduct. Target. Ther. 8, 124 (2023).
Welsh, J. A. et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J. Extracell. Vesicles 13, e12404 (2024).
Ortiz, A. Extracellular vesicles in cancer progression. Semin. Cancer Biol. 76, 139–142 (2021).
Prakash, A., Gates, T., Zhao, X., Wangmo, D. & Subramanian, S. Tumor-derived extracellular vesicles in the colorectal cancer immune environment and immunotherapy. Pharmacol. Ther. 241, 108332 (2023).
Torralba, D. et al. Priming of dendritic cells by DNA-containing extracellular vesicles from activated T cells through antigen-driven contacts. Nat. Commun. 9, 2658 (2018).
Tan, Q. et al. Tumor-derived microparticles-based nanomaterial as platform for delivery of tumor antigens to enhance immunogenicity. Chem. Eng. J. 464, 142497 (2023).
de Jong, O. G. et al. Cellular stress conditions are reflected in the protein and RNA content of endothelial cell-derived exosomes. J. Extracell. Vesicles 1, 18396 (2012).
Yong, T., Wei, Z., Gan, L. & Yang, X. Extracellular-vesicle-based drug delivery systems for enhanced antitumor therapies through modulating the cancer-immunity cycle. Adv. Mater. 34, e2201054 (2022).
Guo, M. et al. Autologous tumor cell–derived microparticle-based targeted chemotherapy in lung cancer patients with malignant pleural effusion. Sci. Transl. Med. 11, eaat5690 (2019).
Chen, W. et al. Tumor cell-derived microparticles packaging monocarboxylate transporter4 inhibitor fluvastatin suppress lung adenocarcinoma via tumor microenvironment remodeling and improve chemotherapy. Chem. Eng. J. 451, 138972 (2023).
Zhou, H. et al. Copper-cysteamine nanoparticle-mediated microwave dynamic therapy improves cancer treatment with induction of ferroptosis. Bioact. Mater. 24, 322–330 (2023).
Wen, B. et al. IQuant: an automated pipeline for quantitative proteomics based upon isobaric tags. Proteomics 14, 2280–2285 (2014).
Hänggi, K. & Ruffell, B. Cell death, therapeutics, and the immune response in cancer. Trends Cancer 9, 381–396 (2023).
Bernard, J. J. et al. Ultraviolet radiation damages self noncoding RNA and is detected by TLR3. Nat. Med. 18, 1286–1290 (2012).
Chen, X. et al. Pyroptosis is driven by non-selective gasdermin-D pore and its morphology is different from MLKL channel-mediated necroptosis. Cell Res. 26, 1007–1020 (2016).
Wandel, M. P. et al. Guanylate-binding proteins convert cytosolic bacteria into caspase-4 signaling platforms. Nat. Immunol. 21, 880–891 (2020).
Iturrioz-Rodríguez, N., De Pasquale, D., Fiaschi, P. & Ciofani, G. Discrimination of glioma patient-derived cells from healthy astrocytes by exploiting Raman spectroscopy. Spectrochim. Acta A 269, 120773 (2022).
Xu, Y. et al. Phenotype identification of HeLa cells knockout CDK6 gene based on label-free Raman imaging. Anal. Chem. 94, 8890–8898 (2022).
Movasaghi, Z., Rehman, S. & Rehman, I. U. Raman spectroscopy of biological tissues. Appl. Spectrosc. Rev. 42, 493–541 (2007).
Koren, E. & Torchilin, V. P. Cell-penetrating peptides: breaking through to the other side. Trends Mol. Med. 18, 385–393 (2012).
Connolly, K. D. et al. Characterisation of adipocyte-derived extracellular vesicles released pre- and post-adipogenesis. J. Extracell. Vesicles 4, 29159 (2015).
Jang, G.-Y. et al. Interactions between tumor-derived proteins and Toll-like receptors. Exp. Mol. Med. 52, 1926–1935 (2020).
Krysko, D. V. et al. Immunogenic cell death and DAMPs in cancer therapy. Nat. Rev. Cancer 12, 860–875 (2012).
Wculek, S. K. et al. Dendritic cells in cancer immunology and immunotherapy. Nat. Rev. Immunol. 20, 7–24 (2020).
Maier, B. et al. A conserved dendritic-cell regulatory program limits antitumour immunity. Nature 580, 257–262 (2020).
Cheng, S. et al. A pan-cancer single-cell transcriptional atlas of tumor infiltrating myeloid cells. Cell 184, 792–809.e23 (2021).
Chen, C.-Y. et al. Combining an alarmin HMGN1 peptide with PD-L1 blockade results in robust antitumor effects with a concomitant increase of stem-like/progenitor exhausted CD8+ T cells. Cancer Immunol. Res. 9, 1214–1228 (2021).
Mildner, A. & Jung, S. Development and function of dendritic cell subsets. Immunity 40, 642–656 (2014).
Caronni, N. et al. IL-1β+ macrophages fuel pathogenic inflammation in pancreatic cancer. Nature 623, 415–422 (2023).
Noy, R. & Pollard, J. W. Tumor-associated macrophages: from mechanisms to therapy. Immunity 41, 49–61 (2014).
Oliveira, G. et al. Phenotype, specificity and avidity of antitumour CD8+ T cells in melanoma. Nature 596, 119–125 (2021).
Bassez, A. et al. A single-cell map of intratumoral changes during anti-PD1 treatment of patients with breast cancer. Nat. Med. 27, 820–832 (2021).
Spitzer, M. H. et al. Systemic immunity is required for effective cancer immunotherapy. Cell 168, 487–502.e15 (2017).
Tong, L. et al. NK cells and solid tumors: therapeutic potential and persisting obstacles. Mol. Cancer 21, 206 (2022).
Cózar, B. et al. Tumor-infiltrating natural killer cells. Cancer Discov. 11, 34–44 (2021).
Liu, Y. et al. Intrapleural nano-immunotherapy promotes innate and adaptive immune responses to enhance anti-PD-L1 therapy for malignant pleural effusion. Nat. Nanotechnol. 17, 206–216 (2022).
Fazio, M., Ablain, J., Chuan, Y., Langenau, D. M. & Zon, L. I. Zebrafish patient avatars in cancer biology and precision cancer therapy. Nat. Rev. Cancer 20, 263–273 (2020).
Fior, R. et al. Single-cell functional and chemosensitive profiling of combinatorial colorectal therapy in zebrafish xenografts. Proc. Natl Acad. Sci. USA 114, E8234–E8243 (2017).
Tong, L. et al. Milk-derived extracellular vesicles protect intestinal barrier integrity in the gut-liver axis. Sci. Adv. 9, eade5041 (2023).
Acknowledgements
Our research has been supported by grants from the National Science and Technology Major Project of the Ministry of Science and Technology of China (number 2022YFF1203300), the National Natural Science Foundation of China (numbers 82330003, 82100112, 82270110 and 82200112). This work was also supported by grants from the National Natural Science Foundation of Hubei Province, China (number 2023AFB1073). We appreciate the available and helpful online drawing tool BioRender (https://biorender.com/).
Author information
Authors and Affiliations
Contributions
Conceptualization, Y.J., Y.W. and W.C.; original draft preparation, W.C., M.G. and Y.W.; review and editing, J.D., X.C., Z.Y. and Y.J.; visualization, J.C. and Q.T.; experiment, W.C., Y.W., J.D., X.C., M.L., J.L. and E.Z.; funding acquisition, M.G. and Y.J. All authors have read and agreed to the published version of the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Nanotechnology thanks Honglin Jin and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Microwave-induced cell death and extracellular vesicle secretion, related to Fig. 2.
(a) The top up-regulated GO pathways in MW-induced LLC cells compared to control, including three categories of biology: biological process (BP), cellular component (CC) and molecular function (MF) (n = 3). (b) Heat maps of DEGs related to cell death. (c) mRNA expressions related to EV secretion (Eea1, Clcn3 and Sh3gl3) and cell death (Casp4, Zbp1 and Gsdmd) in the LLCs stimulated with UV or MW (n = 3). (d) Heat maps of DEGs associated with EV exocytosis (n = 3). The data in (d) are shown in analytic plots as mean ± s.d., and ordinary one-way ANOVA was used. The exact p value was noted.
Extended Data Fig. 2 MW-TMPs facilitate ICD in vitro, related to Fig. 3.
(a) Representative FCM plots of apoptotic analysis for LLC cells treated with MW-TMPs or UV-TMPs by FCM analysis (n = 3). b) FCM analysis of HMGB1 expression in UV-TMPs and MW-TMPs (n = 3). (c, d) Representative FCM plots and quantitative analysis of CD86+ BMDCs (c) and T cell distribution (d) following co-culture of BMDCs and T cells with LLC cells pretreated with either UV-TMP or MW-TMP (n = 3). (e, f) Production of IFN-γ in the co-incubation system (e) or in the presence of the HMGB1 inhibitor (f) (n = 3). The data in (b and c-f) are shown in analytic plots as mean ± s.d. The t test (with two sides) was performed for the two independent groups in (b), and one-way ANOVA was used to test statistical significance among three or more groups in (c-f). The exact p value was noted.
Extended Data Fig. 3 Biodistribution, antitumor efficacy, and biosafety of MW-TMPs in vivo, related to Fig. 3.
(a) Biodistribution of MW-TMPs in tumors and vital organs at 1 hour, 24 hours and 48 hours post-administration (n = 4). (b) Representative images of MW-TMPs accumulation after 1 hour, 24 hours and 48 hours of post-intervention (n = 4). (c) Analysis of DiR-labeled MW-TMPs accumulation in subcutaneous tumors after 1 hour, 24 hours and 48 hours of post-intervention (n = 4). (d) Schematic diagram of treatment protocol for subcutaneous tumor-bearing mice, created in BioRender. Y, Z. (2025) https://BioRender.com/yi407v8. (e) Monitoring of subcutaneous tumor volume during the intervention period (n = 6). (f) Evaluation of the antitumor efficacy of UV-TMPs and MW-TMPs (n = 3). (g) Body weight measurements of subcutaneous tumor models during the administration (n = 6). (h) Representative immunohistochemical images and statistical analysis of TUNEL in tumor sections (n = 4). Scale bar, 100 μm. (i) Schematic illustration of the establishment and treatment of orthotopic lung tumor models, Created in BioRender. Y, Z. (2025) https://BioRender.com/cp6j4hh. (j) The in-vivo representative images of orthotopic lung tumor models at 4 and 12 days (n = 5). (k) Determination of lung weight after mice were sacrificed (n = 6). (l) Representative H&E staining histology of lung tissues following intervention of PBS, UV-TMPs and MW-TMPs (n = 6). Scale bar, 2000 μm. The data in (a, c, f, h and k-l) are shown in analytic plots as mean ± s.d. The t test (with two sides) was performed for the two independent groups in (f), and one-way ANOVA was used among three or more groups in (a, c, h and k-l). Two-way ANOVA was performed in (e and g). The exact p value was noted.
Extended Data Fig. 4 MW-TMPs enhance the immune response of tumor cells, related to Fig. 3.
(a) Umap plot of all the cell types colored by clusters. (b) Dotpot of normalized expression of NK/T, DC Neutrophils, Monocytes/ Macrophages, Endothelial cells, Fibroblasts, Tumor cells marker gene expression among 19 clusters. (c) KEGG enrichment analysis of pathways among tumor clusters (Top 5). (d) The landscape of selected ligand-receptor interactions among tumor cells, DCs, endothelial, fibroblasts, mono/macrophages, neutrophils, NK cells and T cells. (e) Dotpot of ICD-associated genes and apoptosis marker in respective groups. (f) The standardized concentrations of ATP in tumor tissues assessed with an ATP kit (n = 5). (g, h) Representative immunohistochemical analysis of HMGB1 expression in tumor tissue and fluorescent images and intensity analysis of tumor tissues stained with CRT from subcutaneous tumor models (g) and orthotopic lung tumor mice (h) (n = 5). Scale bar, 20 μm. The data in (f-h) are shown in analytic plots as mean ± s.d. Ordinary one-way ANOVA was used among three groups in (f-h). The exact p value was noted.
Extended Data Fig. 5 MW-TMPs activate DCs and enhance the antigen presentation ability of DCs, related to Fig. 4.
(a) Heatmap description of marker genes expression of cDC1s, mregDCs and cDC2s. (b) Heatmap of the differential genes in DCs among clusters. (c) The antigen-presenting gene expression on mregDCs. Box plots display median (center line), IQR (box), 1.5×IQR whiskers, and outliers (open circles). Min/max represent the range of non-outlier values (n = 3). (d) DCs infiltration in tumors (n = 5). (e) Distribution of CD103 + cDC1s in tumor tissues (n = 5). (f) Distribution of CCR7+mregDCs in tumor tissues (n = 5). (g) The proportion of CD86+ in CCR7+mregDCs (n = 5). The data in (d-g) are shown in analytic plots as mean ± s.d. Ordinary one-way ANOVA was used among three groups in (d-g). The exact p value was noted.
Extended Data Fig. 6 MW-TMPs reprogram TAM into an anti-tumor phenotype in TME, related to Fig. 4.
(a) Heatmap of marker genes in six clusters of monocytes/macrophages. (b) The Heatmap of the differential genes in monocytes/macrophages among clusters. (c) Density distribution of different cell types along the pseudotime axis in macrophage. (d) The specific marker genes expression in developmental trajectory. (e) M0, M1, and M2 signature scores among three groups (n = 3). The data are presented in analytic plots as mean ± s.d., and two-way ANOVA was performed for statistical analysis. The exact p value was noted.
Extended Data Fig. 7 MW-TMPs act as a drug-loading nanomaterial and enhance combinational efficacy of anti-PD-L1 immunotherapy, related to Fig. 6.
(a) Standard curve of MTX under HPLC assay (top) and the MTX contents in MW-TMPs-MTX after co-culturing MW-induced LLC cells with various MTX concentrations (50, 100, 200, and 300 μM) (bottom) (n = 3). Avg. Area: Average area. (b) Representative images and corresponding statistical analysis of live (Calcein-AM) /dead (PI) fluorescence staining of LLC stimulated with MW-TMPs or MW-TMPs-MTX for 24 hours (n = 5). Scale bar, 100 μm. (c) Apoptotic analysis of LLC cells treated with MW-TMPs-MTX (n = 3). (d) Schematic illustration of the subcutaneous LLC model establishment and treatment protocols involving the combination of MW-TMPs and anti-PD-L1, created in BioRender. Y, Z. (2025) https://BioRender.com/s12lrdf. (e, f, g) Changes in subcutaneous tumor volume (e, f) and body weight (g) during the intervention of PBS, MW-TMPs and MW-TMPs-MTX (n = 6). (h, i, j) Subcutaneous tumor growth (h, i) and body weight (j) monitored during the intervention of PBS, anti-PD-L1 and the combination group (n = 6). The data in (a-c, f-g and i-j) are shown in analytic plots as mean ± s.d. And one-way ANOVA was used among three or more groups in (a-c). Two-way ANOVA was performed in (f-g and i-j). The exact p value was noted.
Supplementary information
Supplementary Information
Supplementary Figs. 1–9, Tables 1–9, Methods and Discussion.
Supplementary Data 1
Source data for Supplementary Information.
Source data
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 2
Statistical source data.
Source Data Extended Data Fig. 3
Statistical source data.
Source Data Extended Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 7
Statistical source data.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Wu, Y., Chen, W., Deng, J. et al. Tumour-derived microparticles obtained through microwave irradiation induce immunogenic cell death in lung adenocarcinoma. Nat. Nanotechnol. (2025). https://doi.org/10.1038/s41565-025-01922-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41565-025-01922-3
